

Role of Glutamatergic Projections from Lateral Habenula to Ventral Tegmental Area in Inflammatory Pain-Related Spatial Working Memory Deficits

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rodent Model
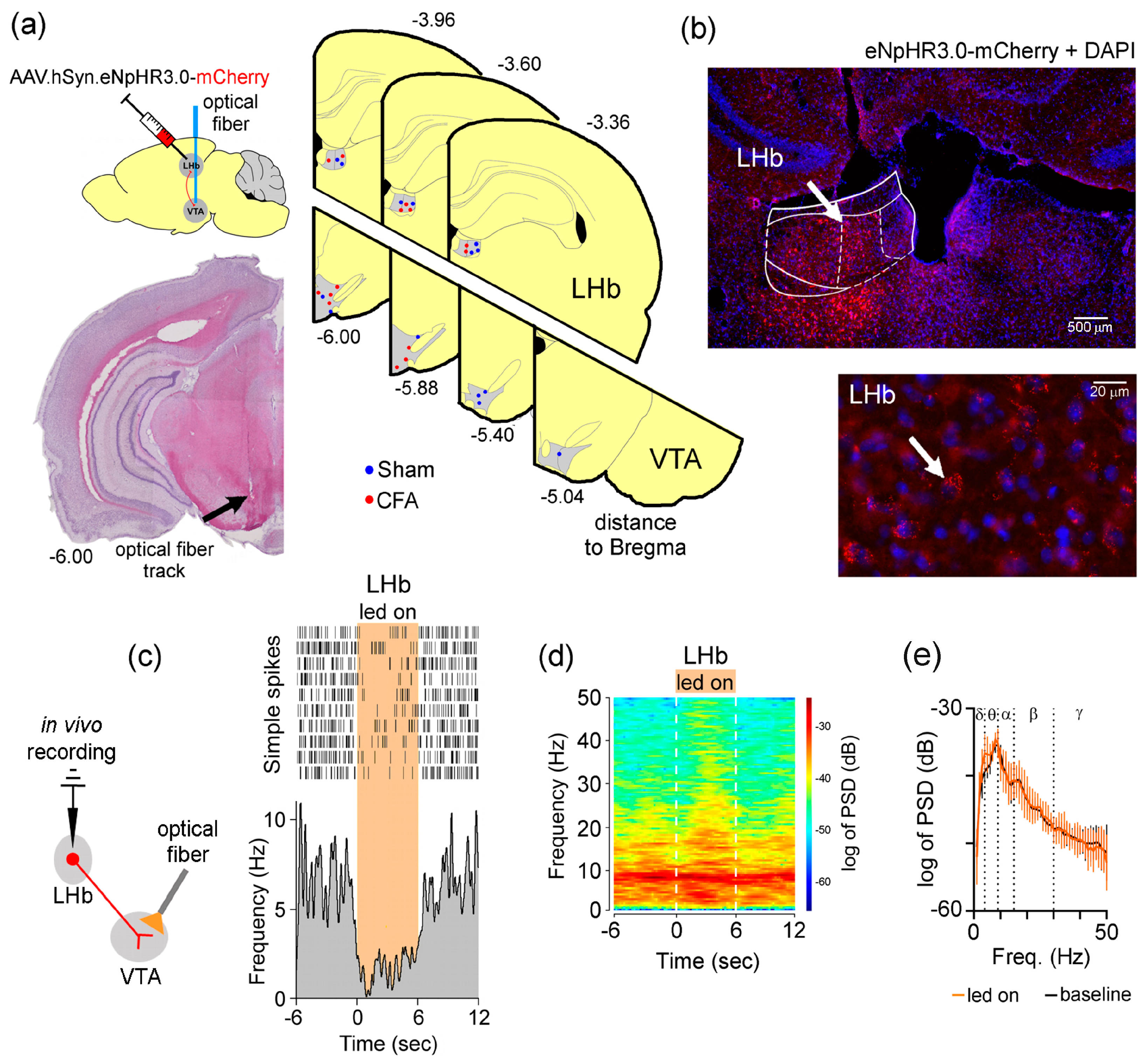
2.2. Stereotaxic Intracranial Virus Transfection, Optical Fiber and Optrode Implantion
2.3. Experimental Design and Behavioral Procedures
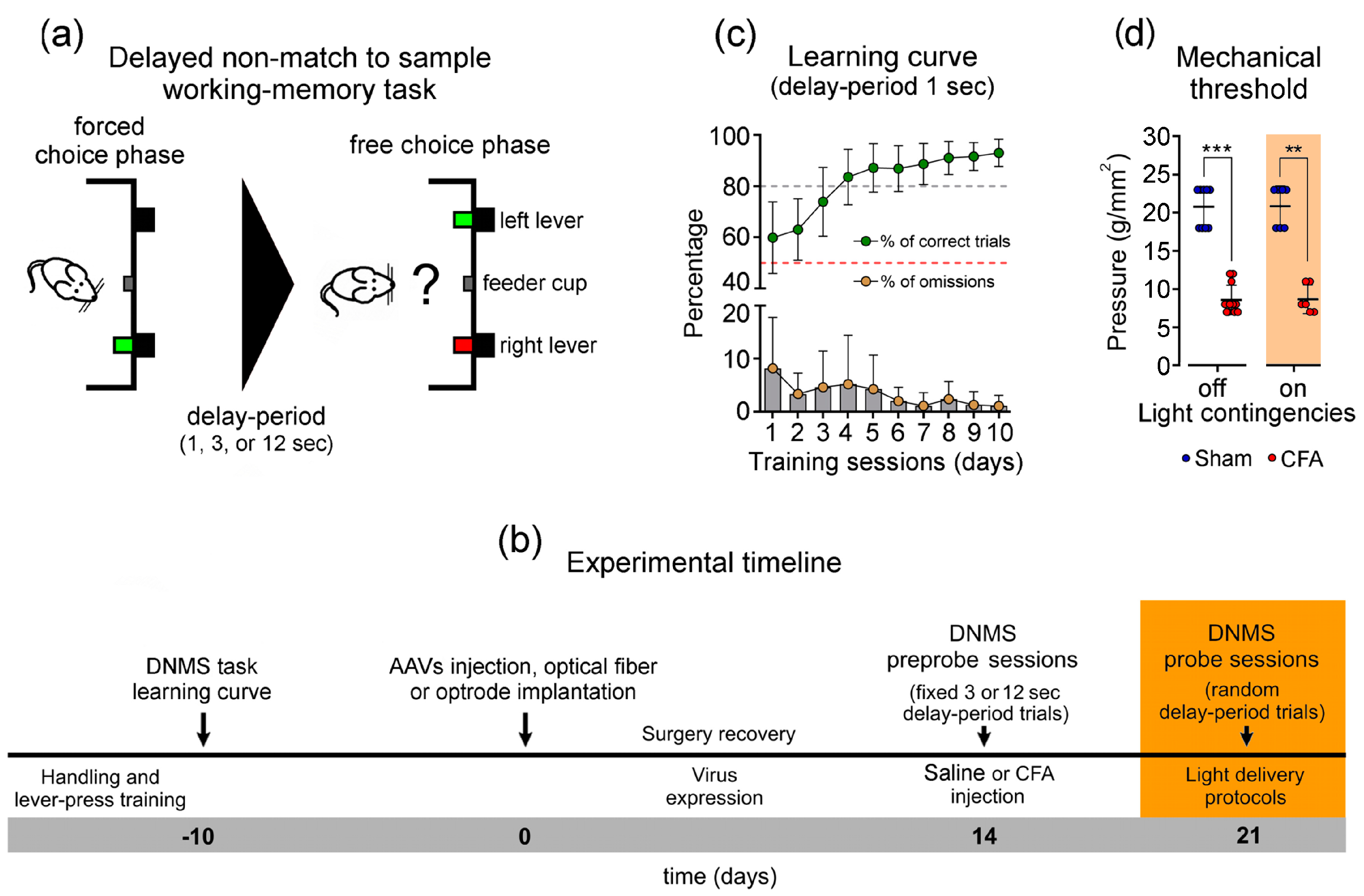
2.3.1. Apparatus and Working Memory Task
2.3.2. Inflammatory Pain Model
2.3.3. Optogenetics and Light Delivery
2.3.4. Neuroelectrophysiological Recordings
2.3.5. Histological Validation
2.3.6. Immunohistochemical Characterization of the VTA Structural Phenotype
2.4. Data Analysis and Representation
3. Results
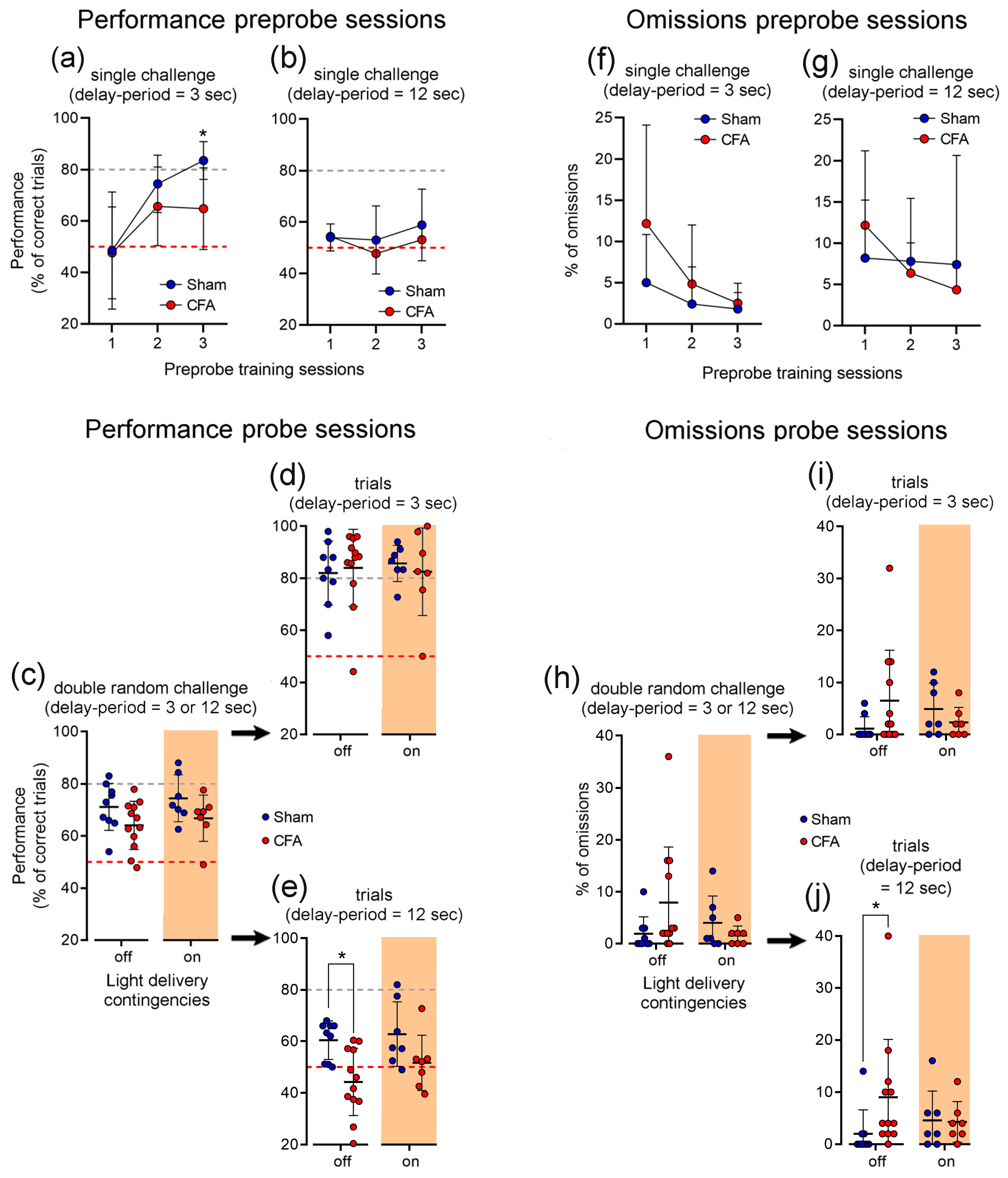
3.1. Working Memory Task and Learning Curve
3.2. Impact of Inhibition of LHb Glutamatergic Terminals into the VTA on Inflammatory Pain
3.3. Specific Optogenetic Inhibition Strategy of LHb Glutamatergic Neurons Terminals into the VTA
3.4. The Impact of Selective Photoinhibition of LHb Glutamatergic Terminals into the VTA in Inflammatory Pain-Related Working Memory Performance
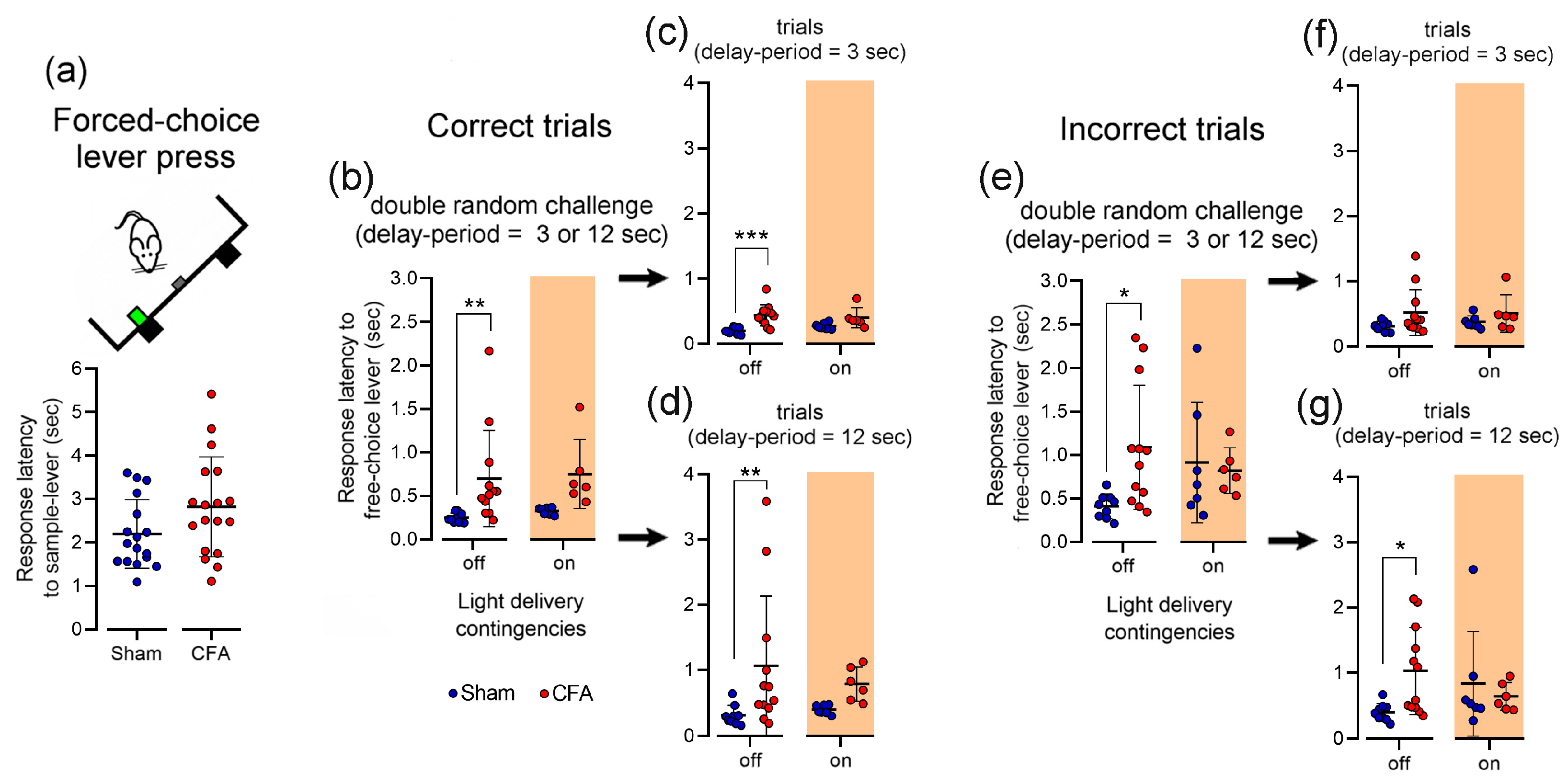
3.5. Effect of Inflammatory Pain on Lever Press Response Latency
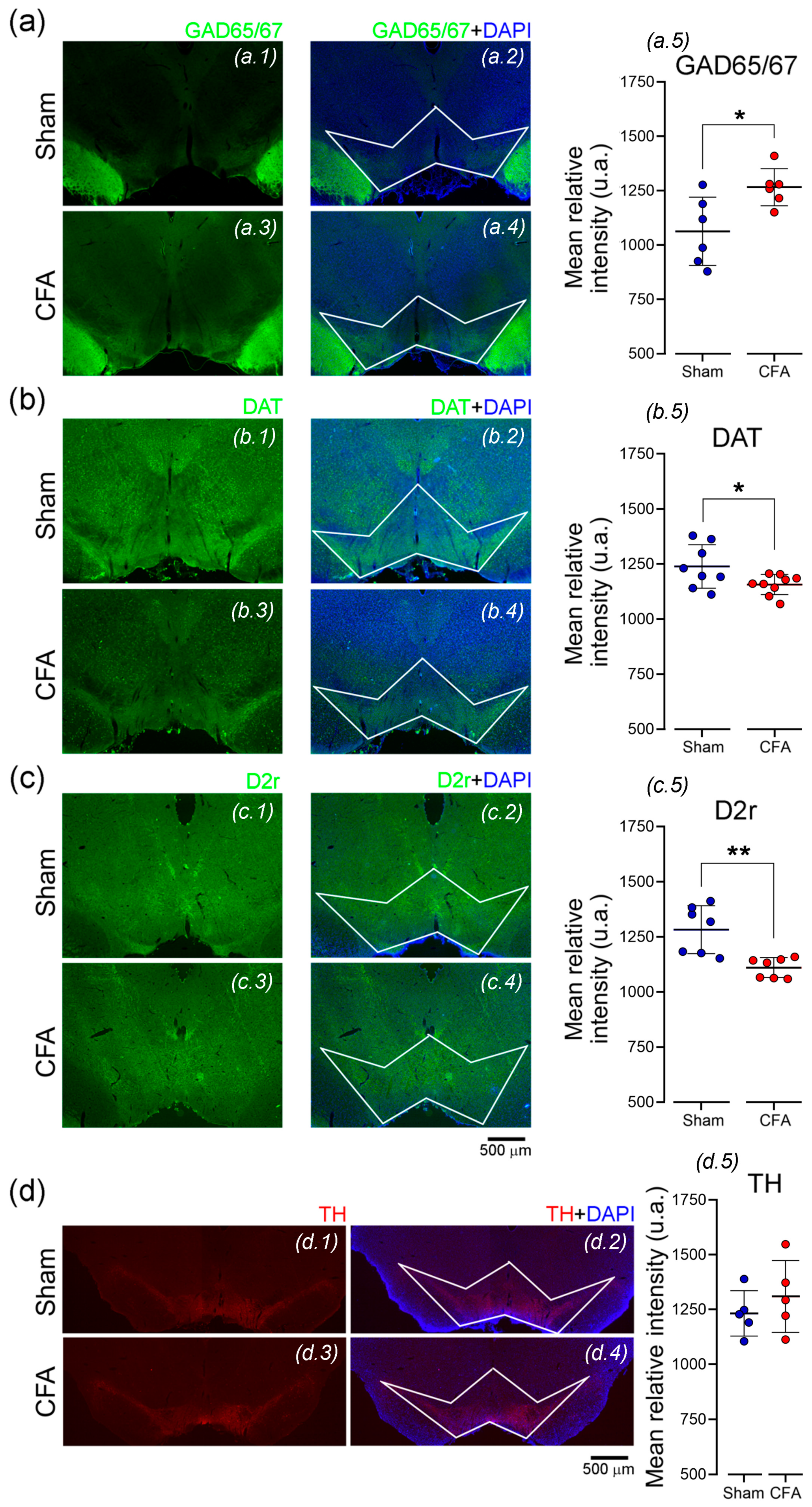
3.6. Pain-Related VTA Structural Phenotype Alterations
4. Discussion
4.1. Role of LHb Glutamatergic Neurons Projecting into the VTA during Cognitive Demand
4.2. Role of LHb Glutamatergic Neurons Projecting into the VTA in Pain-Related Responses
4.3. The Impact of Inflammatory Pain on Local VTA Cell-Type Phenotype
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Furman, D.J.; Gotlib, I.H. Habenula responses to potential and actual loss in major depression: Preliminary evidence for lateralized dysfunction. Soc. Cogn. Affect. Neurosci. 2016, 11, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yang, S.; Liu, X.; Han, Y.; Li, Y.; Feng, J.; Zhao, H. Hypoactivity of the lateral habenula contributes to negative symptoms and cognitive dysfunction of schizophrenia in rats. Exp. Neurol. 2019, 318, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Shelton, L.; Becerra, L.; Borsook, D. Unmasking the mysteries of the habenula in pain and analgesia. Prog. Neurobiol. 2012, 96, 208–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, U.; Lee, T. Topography of descending projections from anterior insular and medial prefrontal regions to the lateral habenula of the epithalamus in the rat. Eur. J. Neurosci. 2012, 35, 1253–1269. [Google Scholar] [CrossRef]
- Vertes, R.P. Interactions among the medial prefrontal cortex, hippocampus and midline thalamus in emotional and cognitive processing in the rat. Neuroscience 2006, 142, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Metz, A.E.; Yau, H.-J.; Centeno, M.V.; Apkarian, A.V.; Martina, M. Morphological and functional reorganization of rat medial prefrontal cortex in neuropathic pain. Proc. Natl. Acad. Sci. USA 2009, 106, 2423–2428. [Google Scholar] [CrossRef] [Green Version]
- Cardoso-Cruz, H.; Lima, D.; Galhardo, V. Impaired spatial memory performance in a rat model of neuropathic pain is associated with reduced hippocampus-prefrontal cortex connectivity. J. Neurosci. 2013, 33, 2465–2480. [Google Scholar] [CrossRef] [Green Version]
- Cardoso-Cruz, H.; Laranjeira, I.; Monteiro, C.; Galhardo, V. Altered prefrontal-striatal theta-band oscillatory dynamics underlie working memory deficits in neuropathic pain rats. Eur. J. Pain 2022, 26, 1546–1568. [Google Scholar] [CrossRef]
- Du, C.X.; Liu, J.; Guo, Y.; Zhang, L.; Zhang, Q.J. Lesions of the lateral habenula improve working memory performance in hemiparkinsonian rats. Neurosci. Lett. 2018, 662, 162–166. [Google Scholar] [CrossRef]
- Lecourtier, L.; DeFrancesco, A.; Moghaddam, B. Differential tonic influence of lateral habenula on prefrontal cortex and nucleus accumbens dopamine release. Eur. J. Neurosci. 2008, 27, 1755–1762. [Google Scholar] [CrossRef] [Green Version]
- Omelchenko, N.; Bell, R.; Sesack, S.R. Lateral habenula projections to dopamine and GABA neurons in the rat ventral tegmental area. Eur. J. Neurosci. 2009, 30, 1239–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stopper, C.M.; Floresco, S.B. What’s better for me? Fundamental role for lateral habenula in promoting subjective decision biases. Nat. Neurosci. 2014, 17, 33–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarcho, J.M.; Mayer, E.A.; Jiang, Z.K.; Feier, N.A.; London, E.D. Pain, affective symptoms, and cognitive deficits in patients with cerebral dopamine dysfunction. Pain 2012, 153, 744–754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lew, S.E.; Tseng, K.Y. Dopamine modulation of GABAergic function enables network stability and input selectivity for sustaining working memory in a computational model of the prefrontal cortex. Neuropsychopharmacology 2014, 39, 3067–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhou, T.C.; Geisler, S.; Marinelli, M.; Degarmo, B.A.; Zahm, D. The mesopontine rostromedial tegmental nucleus: A structure targeted by the lateral habenula that projects to the ventral tegmental area of Tsai and substantia nigra compacta. J. Comp. Neurol. 2009, 513, 566–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shumake, J.; Ilango, A.; Scheich, H.; Wetzel, W.; Ohl, F.W. Differential neuromodulation of acquisition and retrieval of avoidance learning by the lateral habenula and ventral tegmental area. J. Neurosci. 2010, 30, 5876–5883. [Google Scholar] [CrossRef] [Green Version]
- Stuber, G.D.; Stamatakis, A.M.; Kantak, P.A. Considerations when using cre-driver rodent lines for studying ventral tegmental area circuitry. Neuron 2015, 85, 439–445. [Google Scholar] [CrossRef] [Green Version]
- Li, A.-L.; Sibi, J.E.; Yang, X.; Chiao, J.-C.; Peng, Y.B. Stimulation of the ventral tegmental area increased nociceptive thresholds and decreased spinal dorsal horn neuronal activity in rat. Exp. Brain Res. 2016, 234, 1505–1514. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Zhang, B.; Shen, X.; Zhao, H. Why depression and pain often coexist and mutually reinforce: Role of the lateral habenula. Exp. Neurol. 2016, 284 Pt A, 106–113. [Google Scholar] [CrossRef]
- Shelton, L.; Pendse, G.; Maleki, N.; Moulton, E.A.; Lebel, A.; Becerra, L.; Borsook, D. Mapping pain activation and connectivity of the human habenula. J. Neurophysiol. 2012, 107, 2633–2648. [Google Scholar] [CrossRef] [Green Version]
- Brown, P.L.; Palacorolla, H.; Brady, D.; Riegger, K.; Elmer, G.I.; Shepard, P.D. Habenula-Induced Inhibition of Midbrain Dopamine Neurons Is Diminished by Lesions of the Rostromedial Tegmental Nucleus. J. Neurosci. 2017, 37, 217–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breton, J.; Charbit, A.R.; Snyder, B.J.; Fong, P.T.K.; Dias, E.V.; Himmels, P.; Lock, H.; Margolis, E.B. Relative contributions and mapping of ventral tegmental area dopamine and GABA neurons by projection target in the rat. J. Comp. Neurol. 2019, 527, 916–941. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.B.; Sesack, S.R. Projections from the rat prefrontal cortex to the ventral tegmental area: Target specificity in the synaptic associations with mesoaccumbens and mesocortical neurons. J. Neurosci. 2000, 20, 3864–3873. [Google Scholar] [CrossRef] [Green Version]
- Gordon-Fennell, A.; Stuber, G.D. Illuminating subcortical GABAergic and glutamatergic circuits for reward and aversion. Neuropharmacology 2021, 198, 108725. [Google Scholar] [CrossRef] [PubMed]
- Hauser, T.U.; Eldar, E.; Dolan, R.J. Separate mesocortical and mesolimbic pathways encode effort and reward learning signals. Proc. Natl. Acad. Sci. USA 2017, 114, E7395–E7404. [Google Scholar] [CrossRef] [Green Version]
- Martig, A.K.; Mizumori, S.J. Ventral tegmental area disruption selectively affects CA1/CA2 but not CA3 place fields during a differential reward working memory task. Hippocampus 2011, 21, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Ruan, X.; Zhao, H. Stimulation of midbrain dopaminergic structures modifies firing rates of rat lateral habenula neurons. PLoS ONE 2012, 7, e34323. [Google Scholar]
- Cardoso-Cruz, H.; Paiva, P.; Monteiro, C.; Galhardo, V. Selective optogenetic inhibition of medial prefrontal glutamatergic neurons reverses working memory deficits induced by neuropathic pain. Pain 2019, 160, 805–823. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates; Academic: San Diego, CA, USA, 1998. [Google Scholar]
- Aguiar, P.; Mendonça, L.; Galhardo, V. OpenControl: A free opensource software for video tracking and automated control of behavioral mazes. J. Neurosci. Methods 2007, 166, 66–72. [Google Scholar] [CrossRef]
- Butler, S.H.; Godefroy, F.; Besson, J.-M.; Weil-Fugazza, J. A limited arthritic model for chronic pain studies in the rat. Pain 1992, 48, 73–81. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Cruz, H.; Lima, D.; Galhardo, V. Instability of spatial encoding by CA1 hippocampal place cells after peripheral nerve injury. Eur. J. Neurosci. 2011, 33, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantitating confocal fluorescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar] [CrossRef]
- Mathis, V.; Barbelivien, A.; Majchrzak, M.; Mathis, C.; Cassel, J.-C.; Lecourtier, L. The Lateral Habenula as a Relay of Cortical Information to Process Working Memory. Cereb. Cortex. 2017, 27, 5485–5495. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.M.; Jhou, T.; Li, B.; Matsumoto, M.; Mizumori, S.J.; Stephenson-Jones, M.; Vicentic, A. The Lateral Habenula Circuitry: Reward Processing and Cognitive Control. J. Neurosci. 2016, 36, 11482–11488. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.; Kim, S.-H.; Kim, Y.-T.; Song, H.-J.; Lee, J.-J.; Kim, S.-H.; Han, S.W.; Nam, E.J.; Lee, H.J.; Lee, S.-J.; et al. Working memory impairment in fibromyalgia patients associated with altered frontoparietal memory network. PLoS ONE 2012, 7, e37808. [Google Scholar] [CrossRef] [Green Version]
- Cardoso-Cruz, H.; Sousa, M.; Vieira, J.B.; Lima, D.; Galhardo, V. Prefrontal cortex and mediodorsal thalamus reduced connectivity is associated with spatial working memory impairment in rats with inflammatory pain. Pain 2013, 154, 2397–2406. [Google Scholar] [CrossRef]
- Moriarty, O.; McGuire, B.E.; Finn, D.P. The effect of pain on cognitive function: A review of clinical and preclinical research. Prog. Neurobiol. 2011, 93, 385–404. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, M.; Hikosaka, O. Representation of negative motivational value in the primate lateral habenula. Nat. Neurosci. 2009, 12, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Y.; Xuan, C.; Li, Y.; Piao, L.; Li, J.; Zhao, H. Role of the Lateral Habenula in Pain-Associated Depression. Front. Behav. Neurosci. 2017, 11, 31. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Li, J.; Zuo, W.; Chen, P.; Gregor, D.; Fu, R.; Han, X.; Bekker, A.; Ye, J.-H. Downregulation of M-channels in lateral habenula mediates hyperalgesia during alcohol withdrawal in rats. Sci. Rep. 2019, 9, 2714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepard, R.D.; Langlois, L.D.; Browne, C.A.; Berenji, A.; Lucki, I.; Nugent, F.S. Ketamine Reverses Lateral Habenula Neuronal Dysfunction and Behavioral Immobility in the Forced Swim Test Following Maternal Deprivation in Late Adolescent Rats. Front. Synaptic. Neurosci. 2018, 10, 39. [Google Scholar] [CrossRef]
- Wang, D.; Li, Y.; Feng, Q.; Guo, Q.; Zhou, J.; Luo, M. Learning shapes the aversion and reward responses of lateral habenula neurons. Elife 2017, 6, e23045. [Google Scholar] [CrossRef] [PubMed]
- Mathis, V.; Cosquer, B.; Avallone, M.; Cassel, J.-C.; Lecourtier, L. Excitatory Transmission to the Lateral Habenula Is Critical for Encoding and Retrieval of Spatial Memory. Neuropsychopharmacology 2015, 40, 2843–2851. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.; Simkiss, D.; Braddy, D.; Baccus, S.; Morton, T.; Cannady, R.; Weaver, N.; Rose, J.; Levin, E. Nicotinic receptors in the habenula: Importance for memory. Neuroscience 2010, 166, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Barroso, J.; Branco, P.; Apkarian, A.V. Brain mechanisms of chronic pain: Critical role of translational approach. Transl. Res. 2021, 238, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Becker, S.; Schweinhardt, P.; Cahill, C. Mesolimbic dopamine signaling in acute and chronic pain: Implications for motivation, analgesia, and addiction. Pain 2016, 157, 1194–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, W.; Centeno, M.V.; Wei, X.; Wickersham, I.; Martina, M.; Apkarian, A.V.; Surmeier, D.J. Adaptive alterations in the mesoaccumbal network after peripheral nerve injury. Pain 2021, 162, 895–906. [Google Scholar] [CrossRef]
- Cardoso-Cruz, H.; Dourado, M.; Monteiro, C.; Galhardo, V. Blockade of dopamine D2 receptors disrupts intrahippocampal connectivity and enhances pain-related working memory deficits in neuropathic pain rats. Eur. J. Pain 2018, 22, 1002–1015. [Google Scholar] [CrossRef]
- Huang, S.; Borgland, S.; Zamponi, G.W. Peripheral nerve injury-induced alterations in VTA neuron firing properties. Mol. Brain 2019, 12, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tye, K.M.; Mirzabekov, J.J.; Warden, M.R.; Ferenczi, E.A.; Tsai, H.-C.; Finkelstein, J.; Kim, S.-Y.; Adhikari, A.; Thompson, K.R.; Andalman, A.S.; et al. Dopamine neurons modulate neural encoding and expression of depression-related behaviour. Nature 2013, 493, 537–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, D.M.; Hoffman, D.; Benabid, A.L. Simultaneous recording of spontaneous activities and nociceptive responses from neurons in the pars compacta of substantia nigra and in the lateral habenula. Eur. J. Neurosci. 1996, 8, 1474–1478. [Google Scholar] [CrossRef] [PubMed]
- Benabid, A.; Jeaugey, L. Cells of the rat lateral habenula respond to high-threshold somatosensory inputs. Neurosci. Lett. 1989, 96, 289–294. [Google Scholar] [CrossRef]
- Erpelding, N.; Sava, S.; Simons, L.E.; Lebel, A.; Serrano, P.; Becerra, L.; Borsook, D. Habenula functional resting-state connectivity in pediatric CRPS. J. Neurophysiol. 2014, 111, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Khalilzadeh, E.; Saiah, G.V. The possible mechanisms of analgesia produced by microinjection of morphine into the lateral habenula in the acute model of trigeminal pain in rats. Res. Pharm. Sci. 2017, 12, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Du, Y.; Wu, Y.-X.; Guo, F.; Qu, F.-H.; Hu, T.-T.; Tan, B.; Wang, Y.; Hu, W.-W.; Chen, Z.; Zhang, S.-H. Lateral Habenula Serves as a Potential Therapeutic Target for Neuropathic Pain. Neurosci. Bull. 2021, 37, 1339–1344. [Google Scholar] [CrossRef]
- Cui, W.-Q.; Zhang, W.-W.; Chen, T.; Li, Q.; Xu, F.; Mao-Ying, Q.-L.; Mi, W.-L.; Wang, Y.-Q.; Chu, Y.-X. Tacr3 in the lateral habenula differentially regulates orofacial allodynia and anxiety-like behaviors in a mouse model of trigeminal neuralgia. Acta Neuropathol. Commun. 2020, 8, 44. [Google Scholar] [CrossRef] [Green Version]
- Markovic, T.; Pedersen, C.E.; Massaly, N.; Vachez, Y.M.; Ruyle, B.; Murphy, C.A.; Abiraman, K.; Shin, J.H.; Garcia, J.J.; Yoon, H.J.; et al. Pain induces adaptations in ventral tegmental area dopamine neurons to drive anhedonia-like behavior. Nat. Neurosci. 2021, 24, 1601–1613. [Google Scholar] [CrossRef]
- Huang, S.; Zhang, Z.; Gambeta, E.; Xu, S.C.; Thomas, C.; Godfrey, N.; Chen, L.; M’Dahoma, S.; Borgland, S.L.; Zamponi, G.W. Dopamine Inputs from the Ventral Tegmental Area into the Medial Prefrontal Cortex Modulate Neuropathic Pain-Associated Behaviors in Mice. Cell Rep. 2020, 33, 108393. [Google Scholar] [CrossRef]
- Ko, M.Y.; Jang, E.Y.; Lee, J.Y.; Kim, S.P.; Whang, S.H.; Lee, B.H.; Kim, H.Y.; Yang, C.H.; Cho, H.J.; Gwak, Y.S. The Role of Ventral Tegmental Area Gamma-Aminobutyric Acid in Chronic Neuropathic Pain after Spinal Cord Injury in Rats. J. Neurotrauma. 2018, 35, 1755–1764. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.M.; Murphy, N.P.; Evans, C.J.; Cahill, C.M. Correlation between ventral striatal catecholamine content and nociceptive thresholds in neuropathic mice. J. Pain 2014, 15, 878–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, H.; Ku, S.M.; Juarez, B.; Morel, C.; Tzavaras, N.; Montgomery, S.; Hodes, G.E.; Brancato, A.; Russo, S.J.; et al. Sex differences in the neuroadaptations of reward-related circuits in response to subchronic variable stress. Neuroscience 2018, 376, 108–116. [Google Scholar] [CrossRef] [PubMed]
- LaPrairie, J.L.; Murphy, A.Z. Long-term impact of neonatal injury in male and female rats: Sex differences, mechanisms and clinical implications. Front. Neuroendocrinol. 2010, 31, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983, 16, 109–110. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent or Resource | Source | Identifier |
|---|---|---|
| Anti-GAD67 | Merck Millipore | Cat. No. MAB5406 |
| Anti-GAD65/67 | Thermofisher Invitrogen | Cat. No. PA1-84572 |
| Anti-TH | Merck Millipore | Cat. No. MAB318 |
| Anti-D2r | Merck Millipore | Cat. No. MABN53 |
| Anti-DAT | Thermofisher Invitrogen | Cat. No. MA5-24796 |
| Alexa-488 goat anti-mouse | Thermofisher Invitrogen | Cat. No. A-32723 |
| Alexa-568 goat anti-mouse | Thermofisher Invitrogen | Cat. No. A-11004 |
| Alexa-488 goat anti-rabbit | Thermofisher Invitrogen | Cat. No. A-11034 |
| Alexa-647 goat anti-rabbit | Thermofisher Invitrogen | Cat. No. A-32733 |
| AAV5-hSyn-eNpHR3.0-mCherry | NC Univ., USA. | n/a |
| AAV5-CaMKIIα-mCherry | NC Univ., USA. | n/a |
| Complete Freud’s adjuvant | Sigma Aldrich | Cat. No. F5881 |
| 45 mg sucrose pellets | Bioserv, USA. | Cat. No. F0023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alemi, M.; Pereira, A.R.; Cerqueira-Nunes, M.; Monteiro, C.; Galhardo, V.; Cardoso-Cruz, H. Role of Glutamatergic Projections from Lateral Habenula to Ventral Tegmental Area in Inflammatory Pain-Related Spatial Working Memory Deficits. Biomedicines 2023, 11, 820. https://doi.org/10.3390/biomedicines11030820
Alemi M, Pereira AR, Cerqueira-Nunes M, Monteiro C, Galhardo V, Cardoso-Cruz H. Role of Glutamatergic Projections from Lateral Habenula to Ventral Tegmental Area in Inflammatory Pain-Related Spatial Working Memory Deficits. Biomedicines. 2023; 11(3):820. https://doi.org/10.3390/biomedicines11030820
Chicago/Turabian StyleAlemi, Mobina, Ana Raquel Pereira, Mariana Cerqueira-Nunes, Clara Monteiro, Vasco Galhardo, and Helder Cardoso-Cruz. 2023. "Role of Glutamatergic Projections from Lateral Habenula to Ventral Tegmental Area in Inflammatory Pain-Related Spatial Working Memory Deficits" Biomedicines 11, no. 3: 820. https://doi.org/10.3390/biomedicines11030820
APA StyleAlemi, M., Pereira, A. R., Cerqueira-Nunes, M., Monteiro, C., Galhardo, V., & Cardoso-Cruz, H. (2023). Role of Glutamatergic Projections from Lateral Habenula to Ventral Tegmental Area in Inflammatory Pain-Related Spatial Working Memory Deficits. Biomedicines, 11(3), 820. https://doi.org/10.3390/biomedicines11030820