

Knockdown SENP1 Suppressed the Angiogenic Potential of Mesenchymal Stem Cells by Impacting CXCR4-Regulated MRTF-A SUMOylation and CCN1 Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. CD31/KDR Double-Positive Cell Sorting and Flow Cytometry
2.3. Quantitative Real-Time PCR (qRT-PCR)
2.4. Plasmids and Cell Transfection
2.5. SENP1 Knockdown Cells in ECs Differentiation Medium (EDM)
2.6. Uptake of Acetylated Low-Density Lipoprotein (DiI-Ac-LDL)
2.7. Cell Migration Assay
2.8. Transwell Chamber Assay
2.9. Immunocytochemistry Assay
2.10. In Vitro Angiogenesis Assay
2.11. Western Blotting
2.12. Co-Immunoprecipitation
2.13. Luciferase Assay
2.14. Chromosomal Immunoprecipitation (ChIP) Assay
2.15. Statistical Analysis
3. Results
3.1. SENP1 Is Involved in the Differentiation of hMSCs into Artery-Specific Endothelial Cells
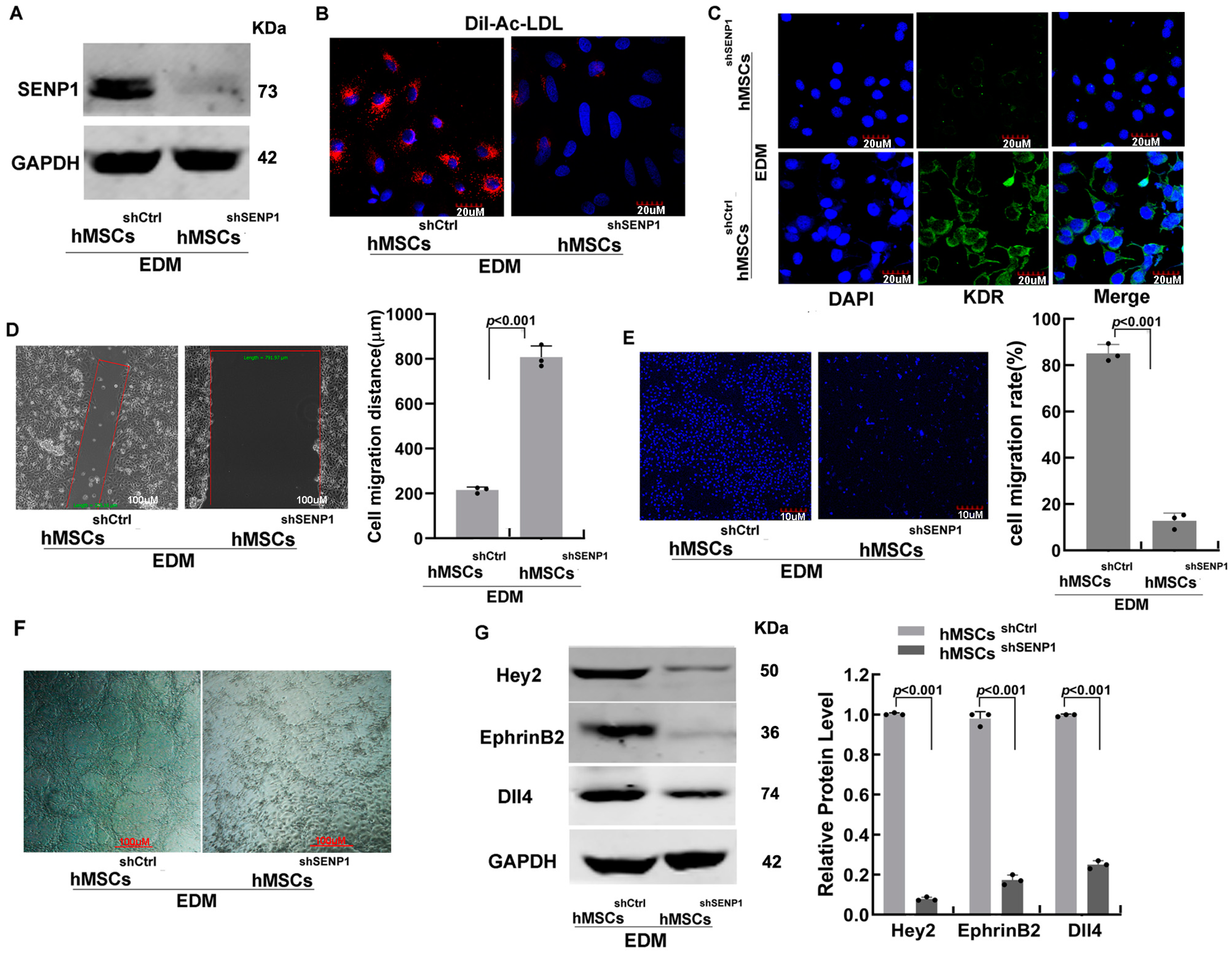
3.2. SENP1 Stable Knockdown Suppresses Endothelial-Specific Differentiation of hMSCs and Exhibits Weaker Migration and Angiogenesis Potential
3.3. SENP1 Knockdown Led to SUMO1 Modification of MRTF-A
3.4. SENP1 Knockdown Suppressed the Expression of CXCR4, Which Was Causatively Related to SUMO1 Modification of MRTF-A and CCN1 Expression
3.5. SUMOylation of MRTF-A Occurred on Three Lysine Residues and Played a Critical Role in Transactivating the Angiogenic Factor CCN1
3.6. SENP1 Knockdown Abolished the Synergetic Induction of CCN1 by the Coordination between MRTF-A and P300
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bagno, L.; Hatzistergos, K.E.; Balkan, W.; Hare, J.M. Mesenchymal Stem Cell-Based Therapy for Cardiovascular Disease: Progress and Challenges. Mol. Ther. 2018, 26, 1610–1623. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.M.; Hur, S.-M.; Park, K.-Y.; Kim, C.-K.; Kim, Y.-M.; Kim, H.-S.; Shin, H.-C.; Won, M.-H.; Ha, K.-S.; Kwon, Y.-G.; et al. Multiple Paracrine Factors Secreted by Mesenchymal Stem Cells Contribute to Angiogenesis. Vascul. Pharmacol. 2014, 63, 19–28. [Google Scholar] [CrossRef]
- Watt, S.M.; Gullo, F.; van der Garde, M.; Markeson, D.; Camicia, R.; Khoo, C.P.; Zwaginga, J.J. The Angiogenic Properties of Mesenchymal Stem/Stromal Cells and Their Therapeutic Potential. Br. Med. Bull. 2013, 108, 25–53. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory Mechanisms of Mesenchymal Stem and Stromal Cells in Inflammatory Diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef]
- Grochot-Przeczek, A.; Dulak, J.; Jozkowicz, A. Therapeutic Angiogenesis for Revascularization in Peripheral Artery Disease. Gene 2013, 525, 220–228. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, N.; Zhang, L.-N.; Huang, N.; Song, T.-F.; Li, Z.-Z.; Li, M.; Luo, X.-G.; Zhou, H.; He, H.-P.; et al. Knockdown of DNMT1 and DNMT3a Promotes the Angiogenesis of Human Mesenchymal Stem Cells Leading to Arterial Specific Differentiation. Stem Cells 2016, 34, 1273–1283. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-G.; Islam, R.; Cho, J.Y.; Jeong, H.; Cap, K.-C.; Park, Y.; Hossain, A.J.; Park, J.-B. Regulation of RhoA GTPase and Various Transcription Factors in the RhoA Pathway. J. Cell Physiol. 2018, 233, 6381–6392. [Google Scholar] [CrossRef]
- Esnault, C.; Stewart, A.; Gualdrini, F.; East, P.; Horswell, S.; Matthews, N.; Treisman, R. Rho-Actin Signaling to the MRTF Coactivators Dominates the Immediate Transcriptional Response to Serum in Fibroblasts. Genes Dev. 2014, 28, 943–958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miralles, F.; Posern, G.; Zaromytidou, A.-I.; Treisman, R. Actin Dynamics Control SRF Activity by Regulation of Its Coactivator MAL. Cell 2003, 113, 329–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medjkane, S.; Perez-Sanchez, C.; Gaggioli, C.; Sahai, E.; Treisman, R. Myocardin-Related Transcription Factors and SRF Are Required for Cytoskeletal Dynamics and Experimental Metastasis. Nat. Cell Biol. 2009, 11, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Vartiainen, M.K.; Guettler, S.; Larijani, B.; Treisman, R. Nuclear Actin Regulates Dynamic Subcellular Localization and Activity of the SRF Cofactor MAL. Science 2007, 316, 1749–1752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baarlink, C.; Wang, H.; Grosse, R. Nuclear Actin Network Assembly by Formins Regulates the SRF Coactivator MAL. Science 2013, 340, 864–867. [Google Scholar] [CrossRef] [PubMed]
- Muehlich, S.; Wang, R.; Lee, S.-M.; Lewis, T.C.; Dai, C.; Prywes, R. Serum-Induced Phosphorylation of the Serum Response Factor Coactivator MKL1 by the Extracellular Signal-Regulated Kinase 1/2 Pathway Inhibits Its Nuclear Localization. Mol. Cell Biol. 2008, 28, 6302–6313. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, K.; Kuzumaki, N. Transcriptional Activity of Megakaryoblastic Leukemia 1 (MKL1) Is Repressed by SUMO Modification. Genes Cells 2005, 10, 835–850. [Google Scholar] [CrossRef]
- Qin, Y.; Yuan, H.; Chen, X.; Yang, X.; Xing, Z.; Shen, Y.; Dong, W.; An, S.; Qi, Y.; Wu, H. SUMOylation Wrestles with the Occurrence and Development of Breast Cancer. Front. Oncol. 2021, 11, 659661. [Google Scholar] [CrossRef]
- Kunz, K.; Piller, T.; Müller, S. SUMO-Specific Proteases and Isopeptidases of the SENP Family at a Glance. J. Cell Sci. 2018, 131, jcs211904. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Ding, S.; Qiu, C.; Shi, Y.; Song, L.; Wang, Y.; Wang, Y.; Li, J.; Wang, Y.; Sun, Y.; et al. SUMOylation Negatively Regulates Angiogenesis by Targeting Endothelial NOTCH Signaling. Circ. Res. 2017, 121, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Jiang, X. SUMOylation of Vascular Endothelial Growth Factor Receptor 2 Inhibits the Proliferation, Migration, and Angiogenesis Signaling Pathway in Non-Small Cell Lung Cancer. Anti-Cancer Drugs 2020, 31, 492–499. [Google Scholar] [CrossRef]
- MacDonald, I.J.; Huang, C.-C.; Liu, S.-C.; Lin, Y.-Y.; Tang, C.-H. Targeting CCN Proteins in Rheumatoid Arthritis and Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 4340. [Google Scholar] [CrossRef]
- Li, J.; Ye, L.; Owen, S.; Weeks, H.P.; Zhang, Z.; Jiang, W.G. Emerging Role of CCN Family Proteins in Tumorigenesis and Cancer Metastasis (Review). Int. J. Mol. Med. 2015, 36, 1451–1463. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-Y.; Su, C.-M.; Hsu, C.-J.; Huang, C.-C.; Wang, S.-W.; Liu, S.-C.; Chen, W.-C.; Fuh, L.-J.; Tang, C.-H. CCN1 Promotes VEGF Production in Osteoblasts and Induces Endothelial Progenitor Cell Angiogenesis by Inhibiting miR-126 Expression in Rheumatoid Arthritis: CCN1 Increases VEGF Expression in Osteoblasts. J. Bone Miner. Res. 2017, 32, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Riese, D.J.; Shen, J. The Role of the CXCL12/CXCR4/CXCR7 Chemokine Axis in Cancer. Front. Pharmacol. 2020, 11, 574667. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Wang, Z.; Ye, D.; Feng, Y.; Liu, M.; Xu, Y.; Wang, M.; Zhang, J.; Liu, J.; Zhao, M.; et al. The Role of CXC Chemokines in Cardiovascular Diseases. Front. Pharmacol. 2022, 12, 765768. [Google Scholar] [CrossRef] [PubMed]
- Zagzag, D.; Lukyanov, Y.; Lan, L.; Ali, M.A.; Esencay, M.; Mendez, O.; Yee, H.; Voura, E.B.; Newcomb, E.W. Hypoxia-Inducible Factor 1 and VEGF Upregulate CXCR4 in Glioblastoma: Implications for Angiogenesis and Glioma Cell Invasion. Lab. Investig. 2006, 86, 1221–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niklason, L.; Dai, G. Arterial Venous Differentiation for Vascular Bioengineering. Annu. Rev. Biomed. Eng. 2018, 20, 431–447. [Google Scholar] [CrossRef]
- Aranguren, X.L.; Luttun, A.; Clavel, C.; Moreno, C.; Abizanda, G.; Barajas, M.A.; Pelacho, B.; Uriz, M.; Araña, M.; Echavarri, A.; et al. In Vitro and In Vivo Arterial Differentiation of Human Multipotent Adult Progenitor Cells. Blood 2007, 109, 2634–2642. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Liu, Z.; Sun, J.; Sun, F.-L.; Li, C.-Z.; Sun, J.-Z.; Xu, L.-Y. The MRTF-A/B Function as Oncogenes in Pancreatic Cancer. Oncol. Rep. 2016, 35, 127–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaqour, B. Caught between a “Rho” and a Hard Place: Are CCN1/CYR61 and CCN2/CTGF the Arbiters of Microvascular Stiffness? J. Cell Commun. Signal. 2020, 14, 21–29. [Google Scholar] [CrossRef]
- Hanna, M.; Liu, H.; Amir, J.; Sun, Y.; Morris, S.W.; Siddiqui, M.A.Q.; Lau, L.F.; Chaqour, B. Mechanical Regulation of the Proangiogenic Factor CCN1/CYR61 Gene Requires the Combined Activities of MRTF-A and CREB-Binding Protein Histone Acetyltransferase. J. Biol. Chem. 2009, 284, 23125–23136. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Wang, D.; Yao, H.; Wei, Z.; Lai, Y.; Hu, J.; Liu, X.; Wang, Y.; Zhou, H.; Wang, N.; et al. Transcriptional Factors P300 and MRTF-A Synergistically Enhance the Expression of Migration-Related Genes in MCF-7 Breast Cancer Cells. Biochem. Biophys. Res. Commun. 2015, 467, 813–820. [Google Scholar] [CrossRef]
- Meng, C.; He, Y.; Wei, Z.; Lu, Y.; Du, F.; Ou, G.; Wang, N.; Luo, X.-G.; Ma, W.; Zhang, T.-C.; et al. MRTF-A Mediates the Activation of COL1A1 Expression Stimulated by Multiple Signaling Pathways in Human Breast Cancer Cells. Biomed. Pharmacother. 2018, 104, 718–728. [Google Scholar] [CrossRef] [PubMed]
- Weinl, C.; Riehle, H.; Park, D.; Stritt, C.; Beck, S.; Huber, G.; Wolburg, H.; Olson, E.N.; Seeliger, M.W.; Adams, R.H.; et al. Endothelial SRF/MRTF Ablation Causes Vascular Disease Phenotypes in Murine Retinae. J. Clin. Investig. 2013, 123, 2193–2206. [Google Scholar] [CrossRef] [Green Version]
- Dorn, T.; Kornherr, J.; Parrotta, E.I.; Zawada, D.; Ayetey, H.; Santamaria, G.; Iop, L.; Mastantuono, E.; Sinnecker, D.; Goedel, A.; et al. Interplay of Cell-Cell Contacts and RhoA/MRTF-A Signaling Regulates Cardiomyocyte Identity. EMBO J. 2018, 37, e98133. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, X.; Wang, H.; Wang, Y.; Chen, P.; Wang, L. Heat Shock Protein 27 Enhances SUMOylation of Heat Shock Protein B8 to Accelerate the Progression of Breast Cancer. Am. J. Pathol. 2020, 190, 2464–2477. [Google Scholar] [CrossRef] [PubMed]
- Ashikari, D.; Takayama, K.; Tanaka, T.; Suzuki, Y.; Obinata, D.; Fujimura, T.; Urano, T.; Takahashi, S.; Inoue, S. Androgen Induces G3BP2 and SUMO-Mediated P53 Nuclear Export in Prostate Cancer. Oncogene 2017, 36, 6272–6281. [Google Scholar] [CrossRef] [PubMed]
- Pandareesh, M.D.; Kameshwar, V.H.; Byrappa, K. Prostate Carcinogenesis: Insights in Relation to Epigenetics and Inflammation. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 253–267. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, H.; Wang, H.; Xiao, F.; Seth, P.; Xu, W.; Jia, Q.; Wu, C.; Yang, Y.; Wang, L. SUMO-Specific Cysteine Protease 1 Promotes Epithelial Mesenchymal Transition of Prostate Cancer Cells via Regulating SMAD4 DeSUMOylation. Int. J. Mol. Sci. 2017, 18, 808. [Google Scholar] [CrossRef] [Green Version]
- Bawa-Khalfe, T.; Yeh, E.T.H. SUMO Losing Balance: SUMO Proteases Disrupt SUMO Homeostasis to Facilitate Cancer Development and Progression. Genes Cancer 2010, 1, 748–752. [Google Scholar] [CrossRef]
- Mo, F.-E.; Muntean, A.G.; Chen, C.-C.; Stolz, D.B.; Watkins, S.C.; Lau, L.F. CYR61 (CCN1) Is Essential for Placental Development and Vascular Integrity. Mol. Cell Biol. 2002, 22, 8709–8720. [Google Scholar] [CrossRef] [Green Version]
- Brigstock, D.R. Regulation of Angiogenesis and Endothelial Cell Function by Connective Tissue Growth Factor (CTGF) and Cysteine-Rich 61 (CYR61). Angiogenesis 2002, 5, 153–165. [Google Scholar] [CrossRef]
- Lee, S.; Elaskandrany, M.; Lau, L.F.; Lazzaro, D.; Grant, M.B.; Chaqour, B. Interplay between CCN1 and Wnt5a in Endothelial Cells and Pericytes Determines the Angiogenic Outcome in a Model of Ischemic Retinopathy. Sci. Rep. 2017, 7, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Ahad, A.; Luu, M.; Moon, S.; Caesar, J.; Cardoso, W.V.; Grant, M.B.; Chaqour, B. CCN1-Yes-Associated Protein Feedback Loop Regulates Physiological and Pathological Angiogenesis. Mol. Cell Biol. 2019, 39, e00107-19. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Liu, K.; Thorne, R.F.; Shi, R.; Zhang, Q.; Wu, M.; Liu, L. Mitochondrial SENP2 Regulates the Assembly of SDH Complex under Metabolic Stress. Cell Rep. 2023, 42, 112041. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, T.; Li, M.; Li, C.; Ma, Y.; Chen, G.; Sun, Y.; Zheng, H.; Wu, G.; Liao, W.; et al. Inhibition of SENP2-Mediated Akt DeSUMOylation Promotes Cardiac Regeneration via Activating Akt Pathway. Clin. Sci. 2021, 135, 811–828. [Google Scholar] [CrossRef]
- Yu, O.M.; Miyamoto, S.; Brown, J.H. Myocardin-Related Transcription Factor A and Yes-Associated Protein Exert Dual Control in G Protein-Coupled Receptor- and RhoA-Mediated Transcriptional Regulation and Cell Proliferation. Mol. Cell Biol. 2016, 36, 39–49. [Google Scholar] [CrossRef] [Green Version]
- Hinkel, R.; Trenkwalder, T.; Petersen, B.; Husada, W.; Gesenhues, F.; Lee, S.; Hannappel, E.; Bock-Marquette, I.; Theisen, D.; Leitner, L.; et al. MRTF-A Controls Vessel Growth and Maturation by Increasing the Expression of CCN1 and CCN2. Nat. Commun. 2014, 5, 3970. [Google Scholar] [CrossRef] [Green Version]
- Morein, D.; Erlichman, N.; Ben-Baruch, A. Beyond Cell Motility: The Expanding Roles of Chemokines and Their Receptors in Malignancy. Front. Immunol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Jiang, Q.; Huang, K.; Lu, F.; Deng, S.; Yang, Z.; Hu, S. Modifying Strategies for SDF-1/CXCR4 Interaction during Mesenchymal Stem Cell Transplantation. Gen. Thorac. Cardiovasc. Surg. 2022, 70, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Lu, P.; Li, L.; Jin, H.; He, X.; Mukaida, N.; Zhang, X. Critical Role of SDF-1α-Induced Progenitor Cell Recruitment and Macrophage VEGF Production in the Experimental Corneal Neovascularization. Mol. Vis. 2011, 17, 2129–2138. [Google Scholar] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, R.; Liu, Q.; Lyu, C.; Gao, X.; Ma, W. Knockdown SENP1 Suppressed the Angiogenic Potential of Mesenchymal Stem Cells by Impacting CXCR4-Regulated MRTF-A SUMOylation and CCN1 Expression. Biomedicines 2023, 11, 914. https://doi.org/10.3390/biomedicines11030914
Zhang R, Liu Q, Lyu C, Gao X, Ma W. Knockdown SENP1 Suppressed the Angiogenic Potential of Mesenchymal Stem Cells by Impacting CXCR4-Regulated MRTF-A SUMOylation and CCN1 Expression. Biomedicines. 2023; 11(3):914. https://doi.org/10.3390/biomedicines11030914
Chicago/Turabian StyleZhang, Rui, Qingxi Liu, Cuicui Lyu, Xing Gao, and Wenjian Ma. 2023. "Knockdown SENP1 Suppressed the Angiogenic Potential of Mesenchymal Stem Cells by Impacting CXCR4-Regulated MRTF-A SUMOylation and CCN1 Expression" Biomedicines 11, no. 3: 914. https://doi.org/10.3390/biomedicines11030914
APA StyleZhang, R., Liu, Q., Lyu, C., Gao, X., & Ma, W. (2023). Knockdown SENP1 Suppressed the Angiogenic Potential of Mesenchymal Stem Cells by Impacting CXCR4-Regulated MRTF-A SUMOylation and CCN1 Expression. Biomedicines, 11(3), 914. https://doi.org/10.3390/biomedicines11030914