
A Comprehensive Review of the Genetic and Epigenetic Contributions to the Development of Fibromyalgia
 ,
,
Abstract
:1. Introduction
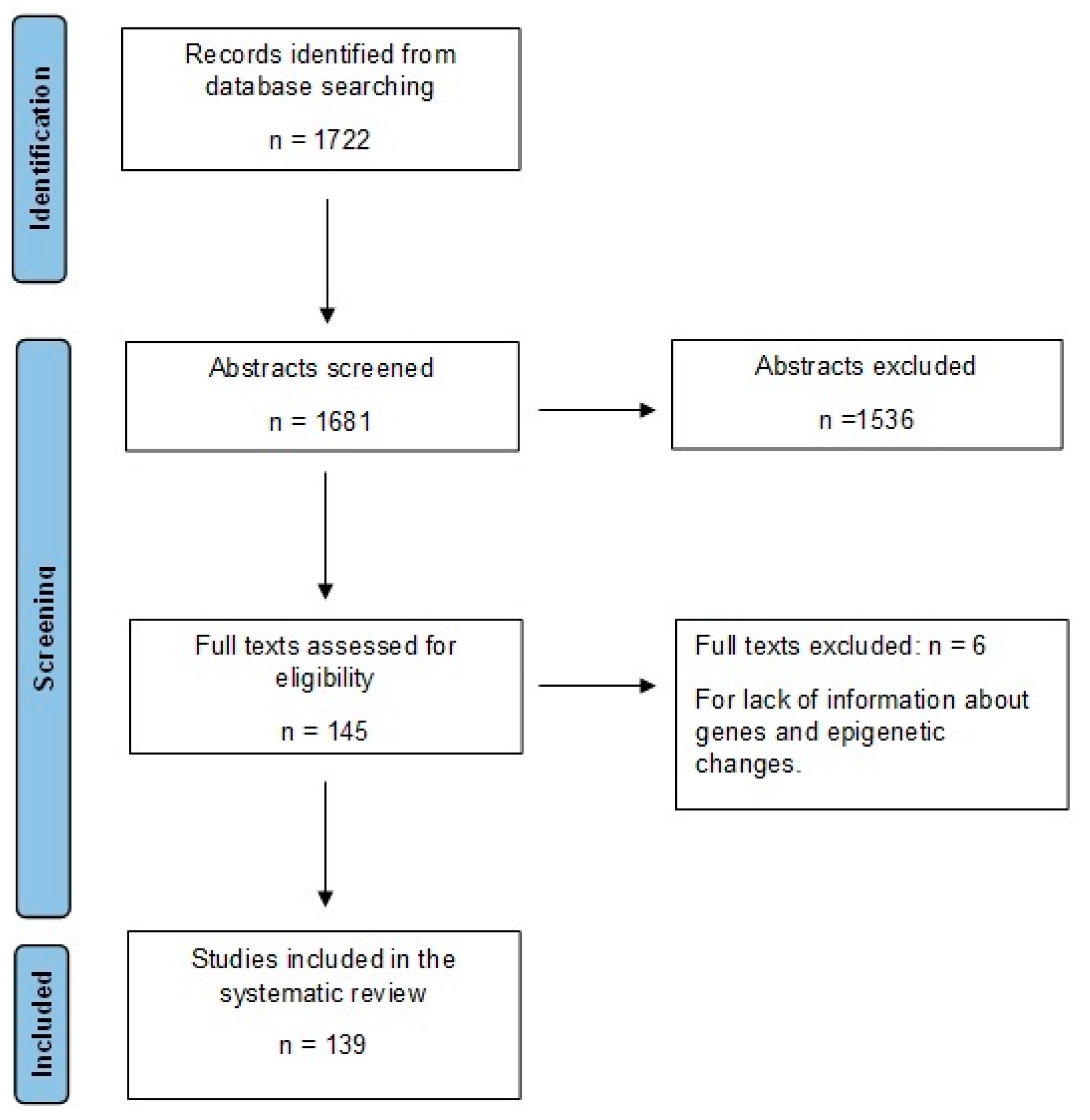
2. Methods
2.1. Literature Search Strategy
2.2. Study Selection
3. FM Pathophysiology
4. Genetic Contributions to the Development of FM
4.1. Catechol-O-Methyltransferase (COMT) Polymorphisms and FM
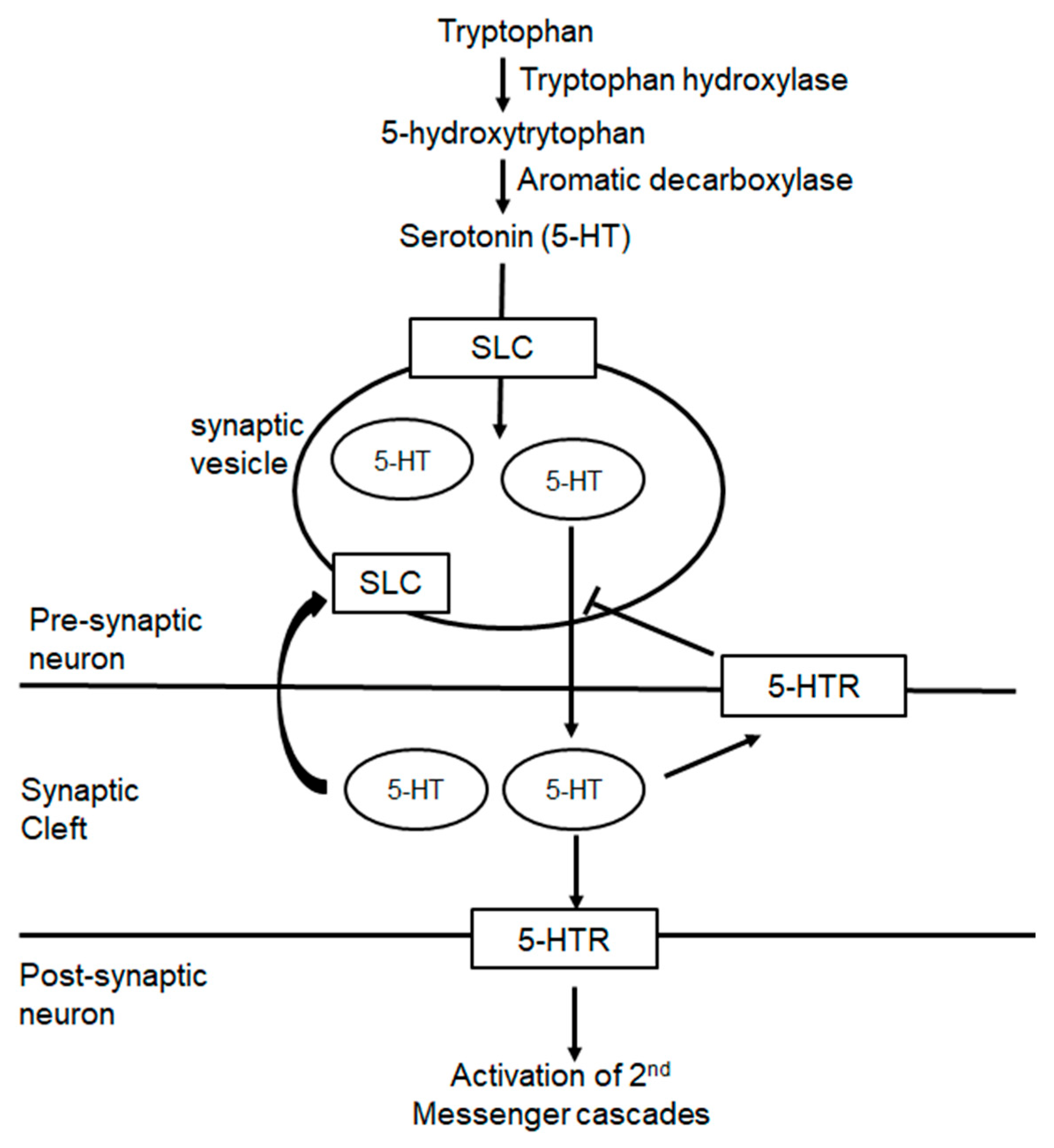
4.2. Polymorphisms in 5-HT Processing and FM
4.3. Polymorphisms in Pain Processing and FM
4.4. Inflammatory Genes/Proteins and FM
4.5. Polymorphisms in Mitochondrial DNA and Vascular Genes and FM
5. Epigenetic Contributions to the Development of FM
5.1. Associations of DNA Methylation Changes and FM
5.2. Micro-RNA and FM
5.3. Upregulation/Over-Expression of Genes and FM

{kind=link}
{kind=link}
| Study | Genes/Proteins | Main Results |
|---|---|---|
| [68] | SNP (rs1800541) EDN1 gene | FM patients had higher plasma levels of EDN1, a potent vasoconstrictor, compared to controls |
| [119] | METTL18, IGL3-25, IL1RAP, IGHV1OR21-1 | Increased METTL18, IGL3-25, IL1RAP, and IGHV1OR21-1 expression can differentiate FM individuals from healthy controls. |
| [120] | Th-17 related genes (14 total), Type 1 IFN related genes (15 total) | Genes involved in the immunologic pathway connected to interleukin-17 and type I interferon were overexpressed in FM individuals compared to controls. |
| [121] | 421 genes exhibited differential expression in FM patient compared to healthy controls | The genes identified are involved in pain processing and axonal development. |
| [122] | TNF-α, interleukin-1β, interleukin-6 | Significant positive correlation between TNF-α, interleukin-1β, and interleukin-6 overexpression and FM symptoms. |
| [123] | TNF-α, interleukin-1β, interleukin-6 | TNF-α, interleukin-1β, and interleukin-6 were upregulated in the skin tissue of FM individuals. |
| [124] | Interleukin-17A, interleukin-2, interleukin-4, interleukin-10, TNF, interferon necrosis-gamma | IL-17A levels were significantly higher in FM individuals. |
| [125] | Interleukin-1Ra, interleukin-8, interleukin-6 | FM individuals had lower serum Interleukin-1Ra levels and normal interleukin-8 and interleukin-6 levels. |
| [126] | 33 genes were overexpressed | Interplay between inflammation, coagulation, and complement cascades contributes to an inflammatory state in FM individuals compared to controls. |
| [128] | NLR, PLR | Systemic inflammatory response marker PLP levels correlated with the number of tender points in FM individuals. |
| [129] | Oleoylethanolamide, palmitoylethanolamide | Plasma levels of oleoylethanolamide and palmitoylethanolamide were significantly higher in FM individuals than in controls. Both are potential indicators of systemic inflammatory state in chronic widespread pain individuals. |
| [130] | CGRP, CLR, RCP | CGRP, CLR, and RCP were elevated in FM versus controls. |
| [132] | S100A8, S100A9, VCAM, CD163, SERPINA1, ANXA1, interleukin-8, AXIN1, SIRT2, STAMBP | Overexpression of identified proteins are associated with an interferon signature in B cells and increased inflammation in FM individuals. |
| [133] | Abeta1-42 | Abeta1-42 was significantly higher in FM individuals. |
| [134] | Neural cell adhesion molecule L1, complement C4-A, lysozyme C, receptor-type tyrosine-protein, phosphatase zeta, apolipoprotein D, alpha-1-antichymotrypsin granulins, calcium/calmodulin-dependent protein kinase type II subunit alpha, mast/stem cell growth factor receptor, prolow-density lipoprotein receptor-related protein 1 | Identified proteins were higher in FM and rheumatoid arthritis patients than controls. These proteins are related to synaptic transmission, inflammatory responses, neuropeptide signaling, and hormonal activity. |
| [135] | Catecholamines, anandamide | Plasma levels of catecholamines and anandamide are higher in FM individuals. |
| [136] | Delta-opioid receptor, Kappa-opioid receptor, mu-opioid receptor | Upregulation of delta and kappa receptor and downregulation of mu receptors in FM individuals compared to controls. |
| [137] | Alpha 2-adrenergic receptors | Upregulated alpha 2-adrenergic receptors in FM individuals positively correlate with receptor number and vasospastic symptoms. |
| [138] | 57 genes linked to hepatic stellate cell activation, oxidative phosphorylation, and airway pathology related to COPD | Expression of these genes can differentiate FM individuals from healthy controls with high validation accuracy. |
| [139] | Eotaxin, MCP-1 | Elevated levels of Eotaxin and MCP-1 in FM individuals versus controls. |
| [140] | Eotaxin-2 | Higher levels of Eotaxin-2 in FM versus controls, but no correlation between marker levels and FM disease severity. |
| [143] | HSP99AA1 | High levels are expressed in FM individuals; higher plasma levels are associated with an increased number of pain sites, fatigue, and decreased motivation. |
| [144] | HSP99AA1 | HSP99AA1 was significantly upregulated in FM individuals. |
| [145] | 12 genes identified, including CENPK, HSP99AA1 | CENPK and HSP99AA1 were significantly elevated in FM women. |
| [147] | 6-SMT | 6-SMT was significantly elevated in FM, but there was no correlation with disease severity. |
| [148] | CatS, CysC | Serum levels of CatS and CysC were higher in FM individuals than controls. |
5.4. Associations of Down-Regulated Genes/Proteins and FM
| Study | Genes/Proteins | Main Results |
|---|---|---|
| [133] | TNF-alpha | Elevated TNF-alpha levels in FM individuals are associated with higher levels of mitochondrial reactive oxygen species and reduced coenzyme Q10 activity. |
| [149] | GABA | FM individuals had reduced GABA levels in the right anterior insula compared to controls. This is positively correlated with lower pressure pain thresholds. |
| [150] | Interleukin-13 | FM individuals had lower interleukin-13 levels than controls. |
| [151] | Interleukin-4, Interleukin-10 | FM individuals had decreased expressions of interleukin -4 and interleukin-10 compared to controls. |
| [154] | EGR1 | FM individuals had lower serum EGR1 compared to controls. |
| [155] | HSP99AA1 | Gender and race may alter levels of HSP99AA1in FM individuals; HSP99AA1 levels lower in non-Caucasian FM individuals; and HSP99AA1 levels lower in Caucasian men with FM compared to controls |
| [155] | Interleukin -4, Interleukin -10, Opioid receptor | FM individuals had reduced opioid receptor binding to F-18-fluoro-ethyl-diprenorphine in the mid cingulate cortex compared to controls. FM individuals had low interleukin-4, interleukin-10 gene expression levels. |
| [156] | Natural killer cells | FM individuals had less natural killer cells than controls, but it was more responsive when exposed to human leukocyte antigen null target cells. |
| [122] | Catalase, glutathione peroxidase, glutathione reductase | Identified enzymes were significantly lower in FM; expression levels correlated with higher oxidative stress parameters compared to controls; and correlated with severity of FM related symptoms. |
6. Conclusions
Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Actual Search Strategy
Appendix A.1. OVID
| # | Searches |
|---|---|
| 1 | Fibromyalgia/ge |
| 2 | Fibromyalgia*/ |
| 3 | fibromyalg*.ti,ab. |
| 4 | 2 or 3 |
| 5 | exp Genetics/ |
| 6 | exp genotype/ |
| 7 | exp genetic phenomena/ |
| 8 | Genetic Markers/ |
| 9 | Phenotype/ |
| 10 | exp Genetic Association Studies/ |
| 11 | Genetic Predisposition to Disease/ |
| 12 | Gene Expression/ |
| 13 | Gene Expression Regulation/ |
| 14 | exp Genetic Variation/ |
| 15 | exp DNA/ |
| 16 | exp RNA/ |
| 17 | exp Genomics/ |
| 18 | exp Epigenomics/ |
| 19 | exp Mutation/ |
| 20 | (gene or genes or genetic* or genotyp* or genom* or epigenetic* or epigenom* or dna* or rna* or mirna* or mrna or polymorphism* or mutation or mutate or sequenc* or proteomic*).ti,ab,kf. |
| 21 | ge.fs. |
| 22 | or/5-21 |
| 23 | 4 and 22 |
| 24 | 1 or 23 |
| 25 | limit 24 to english language [Limit not valid in CDSR; records were retained] |
| 26 | remove duplicates from 25 |
Appendix A.2. SCOPUS
References
- Clauw, D.J. Fibromyalgia: A clinical review. JAMA 2014, 311, 1547–1555. [Google Scholar] [CrossRef]
- Bair, M.J.; Krebs, E.E. Fibromyalgia. Ann. Intern. Med. 2020, 172, ITC33–ITC48. [Google Scholar] [CrossRef]
- Siracusa, R.; Paola, R.D.; Cuzzocrea, S.; Impellizzeri, D. Fibromyalgia: Pathogenesis, Mechanisms, Diagnosis and Treatment Options Update. Int. J. Mol. Sci. 2021, 22, 3891. [Google Scholar] [CrossRef]
- Chinn, S.; Caldwell, W.; Gritsenko, K. Fibromyalgia Pathogenesis and Treatment Options Update. Curr. Pain Headache Rep. 2016, 20, 25. [Google Scholar] [CrossRef]
- Heidari, F.; Afshari, M.; Moosazadeh, M. Prevalence of fibromyalgia in general population and patients, a systematic review and meta-analysis. Rheumatol. Int. 2017, 37, 1527–1539. [Google Scholar] [CrossRef]
- Häuser, W.; Ablin, J.; Fitzcharles, M.A.; Littlejohn, G.; Luciano, J.V.; Usui, C.; Walitt, B. Fibromyalgia. Nat. Rev. Dis. Primers 2015, 1, 15022. [Google Scholar] [CrossRef]
- Mease, P.J. Fibromyalgia, a missed comorbidity in spondyloarthritis: Prevalence and impact on assessment and treatment. Curr. Opin. Rheumatol. 2017, 29, 304–310. [Google Scholar] [CrossRef]
- Løge-Hagen, J.S.; Sæle, A.; Juhl, C.; Bech, P.; Stenager, E.; Mellentin, A.I. Prevalence of depressive disorder among patients with fibromyalgia: Systematic review and meta-analysis. J. Affect. Disord. 2019, 245, 1098–1105. [Google Scholar] [CrossRef]
- Dadabhoy, D.; Crofford, L.J.; Spaeth, M.; Russell, I.J.; Clauw, D.J. Biology and therapy of fibromyalgia. Evidence-based biomarkers for fibromyalgia syndrome. Arthritis Res. Ther. 2008, 10, 211. [Google Scholar] [CrossRef] [Green Version]
- Russell, I.J.; Vaeroy, H.; Javors, M.; Nyberg, F. Cerebrospinal fluid biogenic amine metabolites in fibromyalgia/fibrositis syndrome and rheumatoid arthritis. Arthritis Rheum. 1992, 35, 550–556. [Google Scholar] [CrossRef]
- Wolfe, F.; Russell, I.J.; Vipraio, G.; Ross, K.; Anderson, J. Serotonin levels, pain threshold, and fibromyalgia symptoms in the general population. J. Rheumatol. 1997, 24, 555–559. [Google Scholar] [PubMed]
- Martínez-Jauand, M.; Sitges, C.; Rodríguez, V.; Picornell, A.; Ramon, M.; Buskila, D.; Montoya, P. Pain sensitivity in fibromyalgia is associated with catechol-O-methyltransferase (COMT) gene. Eur. J. Pain 2013, 17, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Ablin, J.N.; Buskila, D. Update on the genetics of the fibromyalgia syndrome. Best Pract. Res. Clin. Rheumatol. 2015, 29, 20–28. [Google Scholar] [CrossRef]
- Haviland, M.G.; Morton, K.R.; Oda, K.; Fraser, G.E. Traumatic experiences, major life stressors, and self-reporting a physician-given fibromyalgia diagnosis. Psychiatry Res. 2010, 177, 335–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, S.; Schweinhardt, P. Dysfunctional neurotransmitter systems in fibromyalgia, their role in central stress circuitry and pharmacological actions on these systems. Pain Res. Treat. 2012, 2012, 741746. [Google Scholar] [CrossRef]
- Crofford, L.J.; Pillemer, S.R.; Kalogeras, K.T.; Cash, J.M.; Michelson, D.; Kling, M.A.; Sternberg, E.M.; Gold, P.W.; Chrousos, G.P.; Wilder, R.L. Hypothalamic-pituitary-adrenal axis perturbations in patients with fibromyalgia. Arthritis Rheum. 1994, 37, 1583–1592. [Google Scholar] [CrossRef]
- Balkarli, A.; Akyol, M.; Tepeli, E.; Balkarli, H.; Temel, S.; Cobankara, V. Haplotypes of vitamin d receptor gene: Both risky and protective for fibromiyalgia (FMS). Ann. Rheum. Dis. 2016, 75, 391–392. [Google Scholar] [CrossRef]
- Sommer, C. CHAPTER 3.11—Serotonin in Pain and Pain Control. In Handbook of Behavioral Neuroscience; Müller, C.P., Jacobs, B.L., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; Volume 21, pp. 457–471. [Google Scholar]
- Chubanov, V.; Waldegger, S.; Mederos y Schnitzler, M.; Vitzthum, H.; Sassen, M.C.; Seyberth, H.W.; Konrad, M.; Gudermann, T. Disruption of TRPM6/TRPM7 complex formation by a mutation in the TRPM6 gene causes hypomagnesemia with secondary hypocalcemia. Proc. Natl. Acad. Sci. USA 2004, 101, 2894–2899. [Google Scholar] [CrossRef] [Green Version]
- Micaroni, M.; Giacchetti, G.; Plebani, R.; Xiao, G.; Federici, L. ATP2C1 gene mutations in Hailey–Hailey disease and possible roles of SPCA1 isoforms in membrane trafficking. Cell Death Dis. 2016, 7, e2259. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.S.; Winsvold, B.S.; Chavez Chavez, S.O.; Borte, S.; Tsepilov, Y.A.; Sharapov, S.Z.; Pain, H.A.-I.; Aulchenko, Y.S.; Hagen, K.; Fors, E.A.; et al. Genome-wide association study identifies RNF123 locus as associated with chronic widespread musculoskeletal pain. Ann. Rheum. Dis. 2021, 80, 1227–1235. [Google Scholar] [CrossRef]
- Sturgill, J.; McGee, E.; Menzies, V. Unique cytokine signature in the plasma of patients with fibromyalgia. J. Immunol. Res. 2014, 2014, 938576. [Google Scholar] [CrossRef] [Green Version]
- Mendieta, D.; De la Cruz-Aguilera, D.L.; Barrera-Villalpando, M.I.; Becerril-Villanueva, E.; Arreola, R.; Hernandez-Ferreira, E.; Perez-Tapia, S.M.; Perez-Sanchez, G.; Garces-Alvarez, M.E.; Aguirre-Cruz, L.; et al. IL-8 and IL-6 primarily mediate the inflammatory response in fibromyalgia patients. J. Neuroimmunol. 2016, 290, 22–25. [Google Scholar] [CrossRef]
- Matsumoto, M.; Weickert, C.S.; Akil, M.; Lipska, B.K.; Hyde, T.M.; Herman, M.M.; Kleinman, J.E.; Weinberger, D.R. Catechol O-methyltransferase mRNA expression in human and rat brain: Evidence for a role in cortical neuronal function. Neuroscience 2003, 116, 127–137. [Google Scholar] [CrossRef]
- Lachman, H.M.; Papolos, D.F.; Saito, T.; Yu, Y.M.; Szumlanski, C.L.; Weinshilboum, R.M. Human catechol-O-methyltransferase pharmacogenetics: Description of a functional polymorphism and its potential application to neuropsychiatric disorders. Pharmacogenetics 1996, 6, 243–250. [Google Scholar] [CrossRef]
- Desmeules, J.; Chabert, J.; Rebsamen, M.; Rapiti, E.; Piguet, V.; Besson, M.; Dayer, P.; Cedraschi, C. Central pain sensitization, COMT Val158Met polymorphism, and emotional factors in fibromyalgia. J. Pain 2014, 15, 129–135. [Google Scholar] [CrossRef]
- Inanir, A.; Karakus, N.; Ates, O.; Sezer, S.; Bozkurt, N.; Inanir, S.; Yigit, S. Clinical symptoms in fibromyalgia are associated to catechol-O-methyltransferase (COMT) gene Val158Met polymorphism. Xenobiotica 2014, 44, 952–956. [Google Scholar] [CrossRef]
- Ferrera, D.; Mercado, F.; Pelaez, I.; Martinez-Inigo, D.; Fernandes-Magalhaes, R.; Barjola, P.; Ecija, C.; Diaz-Gil, G.; Gomez-Esquer, F. Fear of pain moderates the relationship between self-reported fatigue and methionine allele of catechol-O-methyltransferase gene in patients with fibromyalgia. PLoS ONE 2021, 16, e0250547. [Google Scholar] [CrossRef]
- Ferrera, D.; Gomez-Esquer, F.; Pelaez, I.; Barjola, P.; Fernandes-Magalhaes, R.; Carpio, A.; De Lahoz, M.E.; Diaz-Gil, G.; Mercado, F. Effects of COMT Genotypes on Working Memory Performance in Fibromyalgia Patients. J. Clin. Med. 2020, 9, 2479. [Google Scholar] [CrossRef]
- da Silveira Alves, C.F.; Caumo, W.; Silvestri, J.M.; Zortea, M.; Dos Santos, V.S.; Cardoso, D.F.; Regner, A.; de Souza, A.H.; Simon, D. Pain catastrophizing is associated with the Val66Met polymorphism of the brain-derived neurotrophic factor in fibromyalgia. Adv. Rheumatol. 2020, 60, 39. [Google Scholar] [CrossRef]
- Zubieta, J.K.; Heitzeg, M.M.; Smith, Y.R.; Bueller, J.A.; Xu, K.; Xu, Y.; Koeppe, R.A.; Stohler, C.S.; Goldman, D. COMT val158met genotype affects mu-opioid neurotransmitter responses to a pain stressor. Science 2003, 299, 1240–1243. [Google Scholar] [CrossRef]
- Barbosa, F.R.; Matsuda, J.B.; Mazucato, M.; de Castro Franca, S.; Zingaretti, S.M.; da Silva, L.M.; Martinez-Rossi, N.M.; Junior, M.F.; Marins, M.; Fachin, A.L. Influence of catechol-O-methyltransferase (COMT) gene polymorphisms in pain sensibility of Brazilian fibromialgia patients. Rheumatol. Int. 2012, 32, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Neumann, L.; Glazer, Y.; Ebstein, R.P.; Buskila, D. The relationship between a common catechol-O-methyltransferase (COMT) polymorphism val(158) met and fibromyalgia. Clin. Exp. Rheumatol. 2009, 27, S51–S56. [Google Scholar] [PubMed]
- Gursoy, S.; Erdal, E.; Herken, H.; Madenci, E.; Alasehirli, B.; Erdal, N. Significance of catechol-O-methyltransferase gene polymorphism in fibromyalgia syndrome. Rheumatol. Int. 2003, 23, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, J.B.; Barbosa, F.R.; Morel, L.J.; Franca Sde, C.; Zingaretti, S.M.; da Silva, L.M.; Pereira, A.M.; Marins, M.; Fachin, A.L. Serotonin receptor (5-HT 2A) and catechol-O-methyltransferase (COMT) gene polymorphisms: Triggers of fibromyalgia? Rev. Bras. Reumatol. 2010, 50, 141–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas-Alarcon, G.; Fragoso, J.M.; Cruz-Robles, D.; Vargas, A.; Vargas, A.; Lao-Villadoniga, J.I.; Garcia-Fructuoso, F.; Ramos-Kuri, M.; Hernandez, F.; Springall, R.; et al. Catechol-O-methyltransferase gene haplotypes in Mexican and Spanish patients with fibromyalgia. Arthritis Res. Ther. 2007, 9, R110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.H.; Kim, J.H.; Song, G.G. Association between the COMT Val158Met polymorphism and fibromyalgia susceptibility and fibromyalgia impact questionnaire score: A meta-analysis. Rheumatol. Int. 2015, 35, 159–166. [Google Scholar] [CrossRef]
- Park, D.J.; Kim, S.H.; Nah, S.S.; Lee, J.H.; Kim, S.K.; Lee, Y.A.; Hong, S.J.; Kim, H.S.; Lee, H.S.; Kim, H.A.; et al. Association between catechol-O-methyl transferase gene polymorphisms and fibromyalgia in a Korean population: A case-control study. Eur. J. Pain 2016, 20, 1131–1139. [Google Scholar] [CrossRef]
- Potvin, S.; Larouche, A.; Normand, E.; de Souza, J.B.; Gaumond, I.; Grignon, S.; Marchand, S. DRD3 Ser9Gly polymorphism is related to thermal pain perception and modulation in chronic widespread pain patients and healthy controls. J. Pain 2009, 10, 969–975. [Google Scholar] [CrossRef]
- Tander, B.; Gunes, S.; Boke, O.; Alayli, G.; Kara, N.; Bagci, H.; Canturk, F. Polymorphisms of the serotonin-2A receptor and catechol-O-methyltransferase genes: A study on fibromyalgia susceptibility. Rheumatol. Int. 2008, 28, 685–691. [Google Scholar] [CrossRef]
- Estevez-Lopez, F.; Guerrero-Gonzalez, J.M.; Salazar-Tortosa, D.; Camiletti-Moiron, D.; Gavilan-Carrera, B.; Aparicio, V.A.; Acosta-Manzano, P.; Alvarez-Gallardo, I.C.; Segura-Jimenez, V.; Soriano-Maldonado, A.; et al. Interplay between genetics and lifestyle on pain susceptibility in women with fibromyalgia: The al-Andalus project. Rheumatology 2022, 61, 3180–3191. [Google Scholar] [CrossRef]
- Lee, C.; Liptan, G.; Kantorovich, S.; Sharma, M.; Brenton, A. Association of Catechol-O-methyltransferase single nucleotide polymorphisms, ethnicity, and sex in a large cohort of fibromyalgia patients. BMC Rheumatol. 2018, 2, 38. [Google Scholar] [CrossRef]
- Estevez-Lopez, F.; Camiletti-Moiron, D.; Aparicio, V.A.; Segura-Jimenez, V.; Alvarez-Gallardo, I.C.; Soriano-Maldonado, A.; Borges-Cosic, M.; Acosta-Manzano, P.; Geenen, R.; Delgado-Fernandez, M.; et al. Identification of candidate genes associated with fibromyalgia susceptibility in southern Spanish women: The al-Andalus project. J. Transl. Med. 2018, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, T.A.; Nguyen, J.C.; Polglaze, K.E.; Bertrand, P.P. Influence of Tryptophan and Serotonin on Mood and Cognition with a Possible Role of the Gut-Brain Axis. Nutrients 2016, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Al-Nimer, M.S.M.; Mohammad, T.A.M.; Alsakeni, R.A. Serum levels of serotonin as a biomarker of newly diagnosed fibromyalgia in women: Its relation to the platelet indices. J. Res. Med. Sci. 2018, 23, 71. [Google Scholar] [CrossRef]
- Juhl, J.H. Fibromyalgia and the serotonin pathway. Altern. Med. Rev. 1998, 3, 367–375. [Google Scholar]
- Sangkuhl, K.; Klein, T.E.; Altman, R.B. Selective serotonin reuptake inhibitors pathway. Pharm. Genom. 2009, 19, 907–909. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar] [CrossRef] [Green Version]
- Albert, P.R.; Vahid-Ansari, F. The 5-HT1A receptor: Signaling to behavior. Biochimie 2019, 161, 34–45. [Google Scholar] [CrossRef]
- Cohen, H.; Buskila, D.; Neumann, L.; Ebstein, R.P. Confirmation of an association between fibromyalgia and serotonin transporter promoter region (5-HTTLPR) polymorphism, and relationship to anxiety-related personality traits. Arthritis Rheum. 2002, 46, 845–847. [Google Scholar] [CrossRef] [Green Version]
- Offenbaecher, M.; Bondy, B.; de Jonge, S.; Glatzeder, K.; Kruger, M.; Schoeps, P.; Ackenheil, M. Possible association of fibromyalgia with a polymorphism in the serotonin transporter gene regulatory region. Arthritis Rheum. 1999, 42, 2482–2488. [Google Scholar] [CrossRef]
- Klein, R.; Berg, P.A. High incidence of antibodies to 5-hydroxytryptamine, gangliosides and phospholipids in patients with chronic fatigue and fibromyalgia syndrome and their relatives: Evidence for a clinical entity of both disorders. Eur. J. Med. Res. 1995, 1, 21–26. [Google Scholar]
- Ellerbrock, I.; Sandstrom, A.; Tour, J.; Fanton, S.; Kadetoff, D.; Schalling, M.; Jensen, K.B.; Sitnikov, R.; Kosek, E. Serotonergic gene-to-gene interaction is associated with mood and GABA concentrations but not with pain-related cerebral processing in fibromyalgia subjects and healthy controls. Mol. Brain 2021, 14, 81. [Google Scholar] [CrossRef] [PubMed]
- Ledermann, K.; Hasler, G.; Jenewein, J.; Sprott, H.; Schnyder, U.; Martin-Soelch, C. 5’UTR polymorphism in the serotonergic receptor HTR3A gene is differently associated with striatal Dopamine D2/D3 receptor availability in the right putamen in Fibromyalgia patients and healthy controls-Preliminary evidence. Synapse 2020, 74, e22147. [Google Scholar] [CrossRef] [PubMed]
- Frank, B.; Niesler, B.; Bondy, B.; Spath, M.; Pongratz, D.E.; Ackenheil, M.; Fischer, C.; Rappold, G. Mutational analysis of serotonin receptor genes: HTR3A and HTR3B in fibromyalgia patients. Clin. Rheumatol. 2004, 23, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Andolina, G.; Arico, A.; Caccamo, D. TRPM6 gene polymorphisms are highly frequent in patients with fibromyalgia. Clin. Exp. Rheumatol. 2019, 37, S136. [Google Scholar]
- Gloor, Y.; Matthey, A.; Sobo, K.; Mouterde, M.; Kosek, E.; Pickering, G.; Poloni, E.S.; Cedraschi, C.; Ehret, G.; Desmeules, J.A. Uncovering a Genetic Polymorphism Located in Huntingtin Associated Protein 1 in Modulation of Central Pain Sensitization Signaling Pathways. Front. Neurosci. 2022, 16, 807773. [Google Scholar] [CrossRef]
- Du, Q.; Liao, Q.; Chen, C.; Yang, X.; Xie, R.; Xu, J. The role of transient receptor potential vanilloid 1 in common diseases of the digestive tract and the cardiovascular and respiratory system. Front. Physiol. 2019, 10, 1064. [Google Scholar] [CrossRef]
- Park, D.J.; Kim, S.H.; Nah, S.S.; Lee, J.H.; Kim, S.K.; Lee, Y.A.; Hong, S.J.; Kim, H.S.; Lee, H.S.; Kim, H.A.; et al. Polymorphisms of the TRPV2 and TRPV3 genes associated with fibromyalgia in a Korean population. Rheumatology 2016, 55, 1518–1527. [Google Scholar] [CrossRef] [Green Version]
- Gerra, M.C.; Gonzalez-Villar, A.; Arendt-Nielsen, L.; Sokilde Pedersen, I.; Trinanes, Y.; Donnini, C.; Manfredini, M.; Walther, D.; Moeller, G.L.; Pidal-Miranda, M.; et al. A family-based study to identify genetic biomarkers of fibromyalgia: Consideration of patients’ subgroups. Clin. Exp. Rheumatol. 2021, 39 (Suppl. 130), 144–152. [Google Scholar] [CrossRef]
- Alnefeesi, Y.; Tamura, J.K.; Lui, L.M.W.; Jawad, M.Y.; Ceban, F.; Ling, S.; Nasri, F.; Rosenblat, J.D.; McIntyre, R.S. Trace amine-associated receptor 1 (TAAR1): Potential application in mood disorders: A systematic review. Neurosci. Biobehav. Rev. 2021, 131, 192–210. [Google Scholar] [CrossRef]
- Xiao, Y.; He, W.; Russell, I.J. Genetic polymorphisms of the ss2-Adrenergic receptor relate to guanosine protein-coupled stimulator receptor dysfunction in fibromyalgia syndrome. J. Rheumatol. 2011, 38, 1095–1103. [Google Scholar] [CrossRef]
- Vargas-Alarcon, G.; Alvarez-Leon, E.; Fragoso, J.M.; Vargas, A.; Martinez, A.; Vallejo, M.; Martinez-Lavin, M. A SCN9A gene-encoded dorsal root ganglia sodium channel polymorphism associated with severe fibromyalgia. BMC Musculoskelet Disord. 2012, 13, 23. [Google Scholar] [CrossRef] [Green Version]
- Estevez-Lopez, F.; Salazar-Tortosa, D.F.; Camiletti-Moiron, D.; Gavilan-Carrera, B.; Aparicio, V.A.; Acosta-Manzano, P.; Segura-Jimenez, V.; Alvarez-Gallardo, I.C.; Carbonell-Baeza, A.; Munguia-Izquierdo, D.; et al. Fatigue in Women with Fibromyalgia: A Gene-Physical Activity Interaction Study. J. Clin. Med. 2021, 10, 1902. [Google Scholar] [CrossRef]
- Frassoni, C.; Inverardi, F.; Coco, S.; Ortino, B.; Grumelli, C.; Pozzi, D.; Verderio, C.; Matteoli, M. Analysis of SNAP-25 immunoreactivity in hippocampal inhibitory neurons during development in culture and in situ. Neuroscience 2005, 131, 813–823. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, S.H.; Nah, S.S.; Lee, J.H.; Hong, S.J.; Kim, H.S.; Lee, H.S.; Kim, H.A.; Joung, C.I.; Bae, J.; et al. Association of guanosine triphosphate cyclohydrolase 1 gene polymorphisms with fibromyalgia syndrome in a Korean population. J. Rheumatol. 2013, 40, 316–322. [Google Scholar] [CrossRef]
- Ellerbrock, I.; Sandström, A.; Tour, J.; Kadetoff, D.; Schalling, M.; Jensen, K.B.; Kosek, E. Polymorphisms of the μ-opioid receptor gene influence cerebral pain processing in fibromyalgia. Eur. J. Pain 2021, 25, 398–414. [Google Scholar] [CrossRef]
- Nah, S.S.; Lee, H.; Hong, Y.; Im, J.; Won, H.; Chang, S.H.; Kim, H.K.; Kwon, J.T.; Kim, H.J. Association between endothelin-1 and fibromyalgia syndrome. Mol. Med. Rep. 2017, 16, 6234–6239. [Google Scholar] [CrossRef] [Green Version]
- Coskun Benlidayi, I. Role of inflammation in the pathogenesis and treatment of fibromyalgia. Rheumatol. Int. 2019, 39, 781–791. [Google Scholar] [CrossRef]
- Mizel, S.B. The interleukins. FASEB J. 1989, 3, 2379–2388. [Google Scholar] [CrossRef]
- Feng, J.; Zhang, Z.; Wu, X.; Mao, A.; Chang, F.; Deng, X.; Gao, H.; Ouyang, C.; Dery, K.J.; Le, K.; et al. Discovery of potential new gene variants and inflammatory cytokine associations with fibromyalgia syndrome by whole exome sequencing. PLoS ONE 2013, 8, e65033. [Google Scholar] [CrossRef] [Green Version]
- Yigit, S.; Inanir, A.; Tekcan, A.; Inanir, S.; Dural, S.; Ates, O. Association between fibromyalgia syndrome and polymorphism of the IL-4 gene in a Turkish population. Gene 2013, 527, 62–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Yang, Y.K.; Chen, T.; Zhang, H.; Yang, W.W.; Song, S.S.; Zhai, Z.H.; Chen, D.Y. RNF123 has an E3 ligase-independent function in RIG-I-like receptor-mediated antiviral signaling. EMBO Rep. 2016, 17, 1155–1168. [Google Scholar] [CrossRef] [Green Version]
- Park, D.J.; Kim, S.H.; Nah, S.S.; Lee, J.H.; Kim, S.K.; Lee, Y.A.; Hong, S.J.; Kim, H.S.; Lee, H.S.; Kim, H.A.; et al. Association between brain-derived neurotrophic factor gene polymorphisms and fibromyalgia in a Korean population: A multicenter study. Arthritis Res. Ther. 2018, 20, 220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casarotto, P.C.; dos Santos, P.C.; Lucas, G.A.; Biojone, C.; Pobbe, R.L.; Vilela-Costa, H.H.; Joca, S.R.; Guimarães, F.S.; Zangrossi Jr, H. BDNF-TRKB signaling system of the dorsal periaqueductal gray matter is implicated in the panicolytic-like effect of antidepressant drugs. Eur. Neuropsychopharmacol. 2015, 25, 913–922. [Google Scholar] [CrossRef] [PubMed]
- Sikandar, S.; Minett, M.S.; Millet, Q.; Santana-Varela, S.; Lau, J.; Wood, J.N.; Zhao, J. Brain-derived neurotrophic factor derived from sensory neurons plays a critical role in chronic pain. Brain 2018, 141, 1028–1039. [Google Scholar] [CrossRef] [Green Version]
- Smith, Z.D.; Meissner, A. DNA methylation: Roles in mammalian development. Nat. Rev. Genet. 2013, 14, 204–220. [Google Scholar] [CrossRef]
- Nugraha, B.; Anwar, S.L.; Gutenbrunner, C.; Korallus, C. Polymorphisms of brain-derived neurotrophic factor genes are associated with anxiety and body mass index in fibromyalgia syndrome patients. BMC Res. Notes 2020, 13, 402. [Google Scholar] [CrossRef]
- Xiao, Y.; Russell, I.J.; Liu, Y.G. A brain-derived neurotrophic factor polymorphism Val66Met identifies fibromyalgia syndrome subgroup with higher body mass index and C-reactive protein. Rheumatol. Int. 2012, 32, 2479–2485. [Google Scholar] [CrossRef]
- Atherton, K.; Berry, D.J.; Parsons, T.; Macfarlane, G.J.; Power, C.; Hyppönen, E. Vitamin D and chronic widespread pain in a white middle-aged British population: Evidence from a cross-sectional population survey. Ann. Rheum. Dis. 2009, 68, 817–822. [Google Scholar] [CrossRef]
- Achenbach, J.; Rhein, M.; Glahn, A.; Frieling, H.; Karst, M. Leptin promoter methylation in female patients with painful multisomatoform disorder and chronic widespread pain. Clin. Epigenet. 2022, 14, 13. [Google Scholar] [CrossRef]
- van Tilburg, M.A.L.; Parisien, M.; Boles, R.G.; Drury, G.L.; Smith-Voudouris, J.; Verma, V.; Khoury, S.; Chabot-Dore, A.J.; Nackley, A.G.; Smith, S.B.; et al. A genetic polymorphism that is associated with mitochondrial energy metabolism increases risk of fibromyalgia. Pain 2020, 161, 2860–2871. [Google Scholar] [CrossRef]
- Deveci, K.; Deveci, H. Evaluation of the association of MTHFR gene polymorphism with the family history of fibromyalgia syndrome (FMS). Gazi Med. J. 2019, 30, P55. [Google Scholar]
- Blanco, I.; Canto, H.; De Serres, F.J.; Fernandez-Bustillo, E.; Rodriguez, M.C. Alpha1-antitrypsin replacement therapy controls fibromyalgia symptoms in 2 patients with PI ZZ alpha1-antitrypsin deficiency. J. Rheumatol. 2004, 31, 2082–2085. [Google Scholar]
- Yaliwal, L.V.; Desai, R.M. Methylenetetrahydrofolate reductase mutations, a genetic cause for familial recurrent neural tube defects. Indian J. Hum. Genet. 2012, 18, 122–124. [Google Scholar] [CrossRef] [Green Version]
- Gerra, M.C.; Carnevali, D.; Pedersen, I.S.; Donnini, C.; Manfredini, M.; González-Villar, A.; Triñanes, Y.; Pidal-Miranda, M.; Arendt-Nielsen, L.; Carrillo-de-la-Peña, M.T. DNA methylation changes in genes involved in inflammation and depression in fibromyalgia: A pilot study. Scand. J. Pain 2021, 21, 372–383. [Google Scholar] [CrossRef]
- Larbalestier, H.; Keatinge, M.; Watson, L.; White, E.; Gowda, S.; Wei, W.; Koler, K.; Semenova, S.A.; Elkin, A.M.; Rimmer, N.; et al. GCH1 Deficiency Activates Brain Innate Immune Response and Impairs Tyrosine Hydroxylase Homeostasis. J. Neurosci. 2022, 42, 702–716. [Google Scholar] [CrossRef]
- Botto, L.D.; Yang, Q. 5,10-Methylenetetrahydrofolate reductase gene variants and congenital anomalies: A HuGE review. Am. J. Epidemiol. 2000, 151, 862–877. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Zhang, Z.; Li, W.; Shen, X.; Song, W.; Yang, C.; Chang, F.; Longmate, J.; Marek, C.; St Amand, R.P.; et al. Missense mutations in the MEFV gene are associated with fibromyalgia syndrome and correlate with elevated IL-1beta plasma levels. PLoS ONE 2009, 4, e8480. [Google Scholar] [CrossRef]
- Polli, A.; Ghosh, M.; Bakusic, J.; Ickmans, K.; Monteyne, D.; Velkeniers, B.; Bekaert, B.; Godderis, L.; Nijs, J. DNA Methylation and Brain-Derived Neurotrophic Factor Expression Account for Symptoms and Widespread Hyperalgesia in Patients With Chronic Fatigue Syndrome and Comorbid Fibromyalgia. Arthritis Rheumatol. 2020, 72, 1936–1944. [Google Scholar] [CrossRef]
- Menzies, V.; Lyon, D.E.; Archer, K.J.; Zhou, Q.; Brumelle, J.; Jones, K.H.; Gao, G.; York, T.P.; Jackson-Cook, C. Epigenetic alterations and an increased frequency of micronuclei in women with fibromyalgia. Nurs. Res. Pract. 2013, 2013, 795784. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Mullis, A.; Yun, S.; Isla, K.; Estrada, J.; Lukkahatai, N.; Saligan, L.; Walitt, B. A Pilot Methylated DNA Immunoprecipitation of the Death-Associate Protein 3 (Dap3) and Mir2110 in Fibromyalgia Associated with Child Abuse History among African American Females. Eur. Neuropsychopharmacol. 2019, 29, S916–S917. [Google Scholar] [CrossRef]
- Vargas-Alarcon, G.; Fragoso, J.-M.; Cruz-Robles, D.; Vargas, A.; Martinez, A.; Lao-Villadoniga, J.-I.; Garcia-Fructuoso, F.; Vallejo, M.; Martinez-Lavin, M. Association of adrenergic receptor gene polymorphisms with different fibromyalgia syndrome domains. Arthritis Rheum. 2009, 60, 2169–2173. [Google Scholar] [CrossRef] [PubMed]
- Balkarli, A.; Sengul, C.; Tepeli, E.; Balkarli, H.; Cobankara, V. Synaptosomal-associated protein 25 (Snap-25) gene polymorphism frequency in fibromyalgia syndrome and relationship with clinical symptoms. BMC Musculoskelet Disord. 2014, 15, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rush, A.M.; Dib-Hajj, S.D.; Liu, S.; Cummins, T.R.; Black, J.A.; Waxman, S.G. A single sodium channel mutation produces hyper-or hypoexcitability in different types of neurons. Proc. Natl. Acad. Sci. USA 2006, 103, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.B.; Maixner, D.W.; Fillingim, R.B.; Slade, G.; Gracely, R.H.; Ambrose, K.; Zaykin, D.V.; Hyde, C.; John, S.; Tan, K.; et al. Large candidate gene association study reveals genetic risk factors and therapeutic targets for fibromyalgia. Arthritis Rheum. 2012, 64, 584–593. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Ghil, S. Regulators of G-protein signaling, RGS2 and RGS4, inhibit protease-activated receptor 4-mediated signaling by forming a complex with the receptor and Galpha in live cells. Cell Commun. Signal. 2020, 18, 86. [Google Scholar] [CrossRef]
- Zhou, H.; Cheng, Z.; Bass, N.; Krystal, J.H.; Farrer, L.A.; Kranzler, H.R.; Gelernter, J. Genome-wide association study identifies glutamate ionotropic receptor GRIA4 as a risk gene for comorbid nicotine dependence and major depression. Transl. Psychiatry 2018, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Feng, J.; Mao, A.; Le, K.; La Placa, D.; Wu, X.; Longmate, J.; Marek, C.; St Amand, R.P.; Neuhausen, S.L.; et al. SNPs in inflammatory genes CCL11, CCL4 and MEFV in a fibromyalgia family study. PLoS ONE 2018, 13, e0198625. [Google Scholar] [CrossRef]
- Bergin, D.A.; Hurley, K.; McElvaney, N.G.; Reeves, E.P. Alpha-1 antitrypsin: A potent anti-inflammatory and potential novel therapeutic agent. Arch. Immunol. Et Ther. Exp. 2012, 60, 81–97. [Google Scholar] [CrossRef]
- Blanco, I.; de Serres, F.; Janciauskiene, S.; Arbesú, D.; Fernández-Bustillo, E.; Cárcaba, V.; Nita, I.; Astudillo, A. Estimates of the prevalence and number of fibromyalgia syndrome patients and their alpha-1 antitrypsin phenotypic distribution in ten countries. J. Musculoskelet. Pain 2007, 15, 9–23. [Google Scholar] [CrossRef]
- Santos, S.K.F.S.; Fernandes, K.B.P.; Zicarelli, C.A.M.; Santana, A.V.; Perrucini, P.D.D.O.; Poli-Frederico, R.C. Evaluation of ApaI and FokI polymorphism of VDR gene and functional characterization in patients with fibromyalgia. Fisioter. Mov. 2022, 35, e35122. [Google Scholar] [CrossRef]
- Inanir, A.; Yigit, S.; Tekcan, A.; Pinarli, F.A.; Inanir, S.; Karakus, N. Angiotensin converting enzyme and methylenetetrahydrofolate reductase gene variations in fibromyalgia syndrome. Gene 2015, 564, 188–192. [Google Scholar] [CrossRef]
- Ciampi de Andrade, D.; Maschietto, M.; Galhardoni, R.; Gouveia, G.; Chile, T.; Victorino Krepischi, A.C.; Dale, C.S.; Brunoni, A.R.; Parravano, D.C.; Cueva Moscoso, A.S.; et al. Epigenetics insights into chronic pain: DNA hypomethylation in fibromyalgia—A controlled pilot-study. Pain 2017, 158, 1473–1480. [Google Scholar] [CrossRef]
- Burri, A.; Marinova, Z.; Robinson, M.D.; Kühnel, B.; Waldenberger, M.; Wahl, S.; Kunze, S.; Gieger, C.; Livshits, G.; Williams, F. Are Epigenetic Factors Implicated in Chronic Widespread Pain? PLoS ONE 2016, 11, e0165548. [Google Scholar] [CrossRef] [Green Version]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.; Fathy, W.; Abdelaleem, E.A.; Nasser, M.; Yehia, A.; Elanwar, R. The Impact of Micro RNA-320a Serum Level on Severity of Symptoms and Cerebral Processing of Pain in Patients with Fibromyalgia. Pain Med. 2022, 23, 2061–2072. [Google Scholar] [CrossRef]
- Akaslan, E.; Güvener, O.; Görür, A.; Çelikcan, D.H.; Tamer, L.; Biçer, A. The plasma microRNA levels and their relationship with the general health and functional status in female patients with fibromyalgia syndrome. Arch. Rheumatol. 2021, 36, 482–492. [Google Scholar] [CrossRef]
- Bjersing, J.L.; Bokarewa, M.I.; Mannerkorpi, K. Profile of circulating microRNAs in fibromyalgia and their relation to symptom severity: An exploratory study. Rheumatol. Int. 2015, 35, 635–642. [Google Scholar] [CrossRef]
- Iannuccelli, C.; Guzzo, M.P.; Baldassarre, A.; Di Lorro, A.C.; Masotti, A.; Valesini, G.; Di Franco, M. A Microrna Signature in Liquid Biopsies for The Diagnosis of fibromyalgia. Ann. Rheum. Dis. 2016, 75, 1189–1190. [Google Scholar] [CrossRef]
- Erbacher, C.; Vaknine, S.; Moshitzky, G.; Lobentanzer, S.; Eisenberg, L.; Evdokimov, D.; Sommer, C.; Greenberg, D.S.; Soreq, H.; Üçeyler, N. Distinct CholinomiR Blood Cell Signature as a Potential Modulator of the Cholinergic System in Women with Fibromyalgia Syndrome. Cells 2022, 11, 1276. [Google Scholar] [CrossRef]
- Sommer, C.; Leinders, M.; Uceyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Braun, A.; Evdokimov, D.; Frank, J.; Sommer, C.; Üçeyler, N. MiR103a-3p and miR107 are related to adaptive coping in a cluster of fibromyalgia patients. PLoS ONE 2020, 15, e0239286. [Google Scholar] [CrossRef] [PubMed]
- Masotti, A.; Baldassarre, A.; Guzzo, M.P.; Iannuccelli, C.; Barbato, C.; Di Franco, M. Circulating microRNA Profiles as Liquid Biopsies for the Characterization and Diagnosis of Fibromyalgia Syndrome. Mol. Neurobiol. 2017, 54, 7129–7136. [Google Scholar] [CrossRef] [PubMed]
- Leinders, M.; Doppler, K.; Klein, T.; Deckart, M.; Rittner, H.; Sommer, C.; Üçeyler, N. Increased cutaneous miR-let-7d expression correlates with small nerve fiber pathology in patients with fibromyalgia syndrome. Pain 2016, 157, 2493–2503. [Google Scholar] [CrossRef]
- Ide, C. Peripheral nerve regeneration. Neurosci. Res. 1996, 25, 101–121. [Google Scholar] [CrossRef]
- Cerdá-Olmedo, G.; Mena-Durán, A.V.; Monsalve, V.; Oltra, E. Identification of a microRNA signature for the diagnosis of fibromyalgia. PLoS ONE 2015, 10, e0121903. [Google Scholar] [CrossRef] [Green Version]
- Bjersing, J.L.; Lundborg, C.; Bokarewa, M.I.; Mannerkorpi, K. Profile of cerebrospinal microRNAs in fibromyalgia. PLoS ONE 2013, 8, e78762. [Google Scholar] [CrossRef] [Green Version]
- Han, C.L.; Sheng, Y.C.; Wang, S.Y.; Chen, Y.H.; Kang, J.H. Serum proteome profiles revealed dysregulated proteins and mechanisms associated with fibromyalgia syndrome in women. Sci. Rep. 2020, 10, 12347. [Google Scholar] [CrossRef]
- Dolcino, M.; Tinazzi, E.; Puccetti, A.; Lunardi, C. Gene Expression Profiling in Fibromyalgia Indicates an Autoimmune Origin of the Disease and Opens New Avenues for Targeted Therapy. J. Clin. Med. 2020, 9, 1814. [Google Scholar] [CrossRef]
- Jones, K.D.; Gelbart, T.; Whisenant, T.C.; Waalen, J.; Mondala, T.S.; Iklé, D.N.; Salomon, D.R.; Bennett, R.M.; Kurian, S.M. Genome-wide expression profiling in the peripheral blood of patients with fibromyalgia. Clin. Exp. Rheumatol. 2016, 34 (Suppl. 96), S89–S98. [Google Scholar]
- Fatima, G.; Das, S.K.; Mahdi, A.A.; Sarwan, P.K. Deciphering the role of inflammatory cytokines and their correlation with clinical manifestations in women with fibromyalgia syndrome. Intern. Med. J. 2016, 46 (Suppl. 4), 26–27. [Google Scholar] [CrossRef] [Green Version]
- Salemi, S.; Rethage, J.; Wollina, U.; Michel, B.A.; Gay, R.E.; Gay, S.; Sprott, H. Detection of interleukin 1beta (IL-1beta), IL-6, and tumor necrosis factor-alpha in skin of patients with fibromyalgia. J. Rheumatol. 2003, 30, 146–150. [Google Scholar]
- Pernambuco, A.P.; Schetino, L.P.; Alvim, C.C.; Murad, C.M.; Viana, R.S.; Carvalho, L.S.; Reis, D. Increased levels of IL-17A in patients with fibromyalgia. Clin. Exp. Rheumatol. 2013, 31, S60–S63. [Google Scholar]
- Surendran, S.; Mithun, C.B.; Chandran, V.S.; Balan, S.; Tiwari, A. Serum interleukin-6, interleukin-8, and interleukin-1 receptor antagonist levels in South Indian fibromyalgia patients and its correlation with disease severity. J. Rheumatol. 2021, 16, 381–387. [Google Scholar]
- Ramírez-Tejero, J.A.; Martínez-Lara, E.; Rus, A.; Camacho, M.V.; Del Moral, M.L.; Siles, E. Insight into the biological pathways underlying fibromyalgia by a proteomic approach. J. Proteom. 2018, 186, 47–55. [Google Scholar] [CrossRef]
- Balta, S.; Ozturk, C. The platelet-lymphocyte ratio: A simple, inexpensive and rapid prognostic marker for cardiovascular events. Platelets 2015, 26, 680–681. [Google Scholar] [CrossRef]
- Ilgun, E.; Akyürek, Ö.; Kalkan, A.O.; Demir, F.; Demirayak, M.; Bilgi, M. Neutrophil/lymphocyte ratio and platelet/lymphocyte ratio in fibromyalgia. Eur. J. Gen. Med. 2016, 13, 100–104. [Google Scholar]
- Stensson, N.; Ghafouri, B.; Gerdle, B.; Ghafouri, N. Alterations of anti-inflammatory lipids in plasma from women with chronic widespread pain—A case control study. Lipids Health Dis. 2017, 16, 112. [Google Scholar] [CrossRef] [Green Version]
- Korucu, R.U.; Karadağ, A.; Taş, A.; Özmen, E.; Hayta, E.; Siliğ, Y. Serum Calcitonin Gene-Related Peptide and Receptor Protein Levels in Patients with Fibromyalgia Syndrome: A Cross-Sectional Study. Arch. Rheumatol. 2020, 35, 463–467. [Google Scholar] [CrossRef]
- Walsh, D.A.; McWilliams, D.F. CGRP and Painful Pathologies Other than Headache. Handb. Exp. Pharmacol. 2019, 255, 141–167. [Google Scholar] [CrossRef]
- Fineschi, S.; Klar, J.; Gustafsson, K.A.; Jonsson, K.; Karlsson, B.; Dahl, N. Inflammation and Interferon Signatures in Peripheral B-Lymphocytes and Sera of Individuals with Fibromyalgia. Front. Immunol. 2022, 13, 874490. [Google Scholar] [CrossRef] [PubMed]
- Lo, Y.C.; Li, T.J.T.; Lin, T.C.; Chen, Y.Y.; Kang, J.H. Microstructural Evidence of Neuroinflammation for Psychological Symptoms and Pain in Patients with Fibromyalgia. J. Rheumatol. 2022, 49, 942–947. [Google Scholar] [CrossRef] [PubMed]
- Khoonsari, P.E.; Ossipova, E.; Lengqvist, J.; Svensson, C.I.; Kosek, E.; Kadetoff, D.; Jakobsson, P.J.; Kultima, K.; Lampa, J. The human CSF pain proteome. J. Proteom. 2019, 190, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, I.; Schelling, G.; Eisner, C.; Richter, H.P.; Krauseneck, T.; Vogeser, M.; Hauer, D.; Campolongo, P.; Chouker, A.; Beyer, A.; et al. Anandamide and neutrophil function in patients with fibromyalgia. Psychoneuroendocrinology 2008, 33, 676–685. [Google Scholar] [CrossRef]
- Salemi, S.; Aeschlimann, A.; Wollina, U.; Gay, R.E.; Michel, B.A.; Gay, S.; Sprott, H. Up-regulation of delta-opioid receptors and kappa-opioid receptors in the skin of fibromyalgia patients. Arthritis Rheum. 2007, 56, 2464–2466. [Google Scholar] [CrossRef]
- Bennett, R.M.; Clark, S.R.; Campbell, S.M.; Ingram, S.B.; Burckhardt, C.S.; Nelson, D.L.; Porter, J.M. Symptoms of Raynaud’s syndrome in patients with fibromyalgia. A study utilizing the Nielsen test, digital photoplethysmography, and measurements of platelet alpha 2-adrenergic receptors. Arthritis Rheum. 1991, 34, 264–269. [Google Scholar] [CrossRef]
- Lukkahatai, N.; Walitt, B.; Deandrés-Galiana, E.J.; Fernández-Martínez, J.L.; Saligan, L.N. A predictive algorithm to identify genes that discriminate individuals with fibromyalgia syndrome diagnosis from healthy controls. J. Pain Res. 2018, 11, 2981–2990. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Cherryholmes, G.; Mao, A.; Marek, C.; Longmate, J.; Kalos, M.; Amand, R.P.S.T.; Shively, J.E. High plasma levels of MCP-1 and eotaxin provide evidence for an immunological basis of fibromyalgia. Exp. Biol. Med. 2008, 233, 1171–1180. [Google Scholar] [CrossRef]
- Furer, V.; Hazan, E.; Mor, A.; Segal, M.; Katav, A.; Aloush, V.; Elkayam, O.; George, J.; Ablin, J.N. Elevated Levels of Eotaxin-2 in Serum of Fibromyalgia Patients. Pain Res. Manag. 2018, 2018, 7257681. [Google Scholar] [CrossRef]
- Shan, Q.; Ma, F.; Wei, J.; Li, H.; Ma, H.; Sun, P. Physiological Functions of Heat Shock Proteins. Curr. Protein Pept. Sci. 2020, 21, 751–760. [Google Scholar] [CrossRef]
- Zugel, U.; Kaufmann, S.H. Role of heat shock proteins in protection from and pathogenesis of infectious diseases. Clin. Microbiol. Rev. 1999, 12, 19–39. [Google Scholar] [CrossRef] [Green Version]
- Kawi, J.; Lukkahatai, N.; Espina, A.; Walitt, B.; Saligan, L. Associations between heat shock protein and symptom burden in Fibromyalgia Syndrome (FMS). J. Pain 2016, 17 (Suppl. 1), S30–S31. [Google Scholar] [CrossRef]
- Majors, B.; Saligan, L.N.; Wang, D.; Walitt, B. Differential expression of HSP90AA1 associated with fibromyalgia. Clin. Transl. Sci. 2012, 5, 183–184. [Google Scholar]
- Lukkahatai, N.; Walitt, B.; Espina, A.; Wang, D.; Saligan, L.N. Comparing Genomic Profiles of Women with and without Fibromyalgia. Biol. Res. Nurs. 2015, 17, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Lukkahatai, N.; Kaur, K.; Walitt, B.; Kawi, J.; Espina, A.; Saligan, L. Differences in plasma levels of heat shock proteins between persons with and without fibromyalgia. J. Pain 2016, 17 (Suppl. 1), S31–S32. [Google Scholar] [CrossRef]
- Pernambuco, A.P.; Schetino, L.P.; Viana, R.S.; Carvalho, L.S.; d’Ávila Reis, D. The involvement of melatonin in the clinical status of patients with fibromyalgia syndrome. Clin. Exp. Rheumatol. 2015, 33, S14–S19. [Google Scholar]
- Kocak, I.; Hizmetli, S.; Tas, A.; Karadag, A.; Zontul, C.; Silig, Y. High levels of cathepsin S and cystatin C in patients with fibromyalgia syndrome. Int. J. Rheum. Dis. 2020, 23, 966–969. [Google Scholar] [CrossRef]
- Foerster, B.R.; Petrou, M.; Edden, R.A.; Sundgren, P.C.; Schmidt-Wilcke, T.; Lowe, S.E.; Harte, S.E.; Clauw, D.J.; Harris, R.E. Reduced insular γ-aminobutyric acid in fibromyalgia. Arthritis Rheum. 2012, 64, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Keskin, G.; İnal, A.; Keskin, D.; Muşabak, U.; Şengül, A.; Köse, K. Serum interleukin-13 (IL-13) levels in patients with fibromyalgia. Gulhane Med. J. 2008, 50, 257–260. [Google Scholar]
- Uçeyler, N.; Valenza, R.; Stock, M.; Schedel, R.; Sprotte, G.; Sommer, C. Reduced levels of antiinflammatory cytokines in patients with chronic widespread pain. Arthritis Rheum. 2006, 54, 2656–2664. [Google Scholar] [CrossRef]
- Cordero, M.D.; Díaz-Parrado, E.; Carrión, Á.M.; Alfonsi, S.; Sánchez Alcázar, J.A.; Miguel Rodríguez, M.D.; Battino, M. Is inflammation a mitochondrial dysfunction-dependent event in fibromyalgia? Antioxid. Redox Signal. 2013, 18, 800–807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fatima, G.; Das, S.K.; Mahdi, A.A. Some oxidative and antioxidative parameters and their relationship with clinical symptoms in women with fibromyalgia syndrome. Int. J. Rheum. Dis. 2017, 20, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Tas, A.; Ağbektaş, T.; Karadağ, A.; Hayta, E.; Siliğ, Y. Early growth response protein (EGR) levels in patients with fibromyalgia syndrome. Turk. J. Biochem. 2021, 46 (Suppl. 2), 63. [Google Scholar]
- Üçeyler, N.; Buchholz, H.G.; Kewenig, S.; Ament, S.J.; Birklein, F.; Schreckenberger, M.; Sommer, C. Cortical Binding Potential of Opioid Receptors in Patients with Fibromyalgia Syndrome and Reduced Systemic Interleukin-4 Levels—A Pilot Study. Front. Neurosci. 2020, 14, 512. [Google Scholar] [CrossRef]
- Verma, V.; Drury, G.L.; Parisien, M.; Özdağ Acarli, A.N.; Al-Aubodah, T.A.; Nijnik, A.; Wen, X.; Tugarinov, N.; Verner, M.; Klares, R.; et al. Unbiased immune profiling reveals a natural killer cell-peripheral nerve axis in fibromyalgia. Pain 2022, 163, e821–e836. [Google Scholar] [CrossRef]
- Kubota, T.; Miyake, K.; Hirasawa, T. Epigenetic understanding of gene-environment interactions in psychiatric disorders: A new concept of clinical genetics. Clin. Epigenet. 2012, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Lavín, M. Fibromyalgia in women: Somatisation or stress-evoked, sex-dimorphic neuropathic pain? Clin. Exp. Rheumatol. 2021, 39, 422–425. [Google Scholar] [CrossRef]
- D’Agnelli, S.; Arendt-Nielsen, L.; Gerra, M.C.; Zatorri, K.; Boggiani, L.; Baciarello, M.; Bignami, E. Fibromyalgia: Genetics and epigenetics insights may provide the basis for the development of diagnostic biomarkers. Mol. Pain 2019, 15, 1–12. [Google Scholar] [CrossRef]


| Study | Genes | Effects in FM |
|---|---|---|
| [12,32,33,34,35,36] | COMT Val-158-Met (rs4680) | Increased risk of developing FM |
| [12,36,42] | rs4818 of the COMT gene | No increased risk of FM development in diverse, Mexican, or Spanish populations |
| [32,36,38] | rs4818 of the COMT gene | Increased risk of FM development in Korean, Brazilian, and Spanish populations |
| [36] | rs6269 of the COMT gene | Increased risk of FM development in the Spanish population |
| [37,38,39,40] | COMT Val-158-Met (rs4680) | No increased risk of developing FM |
| [38] | rs4633 of the COMT gene | Increased risk of FM development in the Korean population |
| [42] | COMT Val-158-Met (rs4680) | Increased severity of symptoms |
| [43] | rs2097903 of the COMT gene | Increased risk of FM development |
| Study | Genes (Polymorphisms) | Effects In FM |
|---|---|---|
| [41] | 5-HTR2A, CT polymorphism genotype | Lower pain threshold |
| [50,51] | SLC6A4 “short allele” | Increased risk of FM |
| [53] | 5-HT1a, CC/G polymorphism alone and with 5-HTT-low | Increased depressive symptoms |
| 5HT1a, CC polymorphism with 5-HTT-high | Fewest depressive symptoms, highest response to an SSRI | |
| [54] | 5-HTR3a, SNP rs1062613 CC homozygote | Increased risk of FM |
| [55] | 5-HTR3a, 5-HTR3b | Unsure currently |
| Study | Genes | Effects in FM |
|---|---|---|
| [21] | ATP2C1 | Regulates calcium homeostasis through the hSPCA1 magnesium-dependent calcium channel and affects nerve function. |
| [60] | HAP1 | Regulates vesicular trafficking and lowers the nociceptive flexion reflex threshold. |
| [61] | SCN9A | Encodes the Nav1.7 sodium channel; the rs4796604 SNP associated with reduced motivation, reduced activity, and higher FM impact questionnaire scores. |
| [62] | OPRM1 | Modulates cerebral pain processing. |
| [63,64] | TRPV3 | Plays a role in the sensation of noxious stimuli; the rs395357 SNP increases the symptom severity of fatigue and mental health in FM. |
| [65] | TRPM6 | Regulates calcium and magnesium homeostasis in neurons. |
| [66,67] | SNAP25 | Regulates neurotransmitter release via vesicle docking and fusion; TC genotype is associated with higher pain and depression scores |
| [68] | GCH1 | Involved in dopamine and serotonin synthesis. rs3783641, rs84, rs752688, and rs4411417 SNPs are associated with lower pain sensitivity in FM. |
| [69] | TAAR1 | Encodes G-protein-coupled receptor and plays a role in reward and cognitive function. Increased levels decrease reward seeking behavior and increase attention and focus. |
| [70] | RGS4 | Encodes a GTPase that interacts with PAR4 GPCR and negatively regulates G-protein signaling. |
| [71] | GRIA4 | Encodes a glutamate receptor that mediates excitatory neurotransmission. |
| [72] | Beta2-Adrenergic Receptor gene | The Gly16Arg SNP increases the risk of FM and may cause sleep dysfunction in FM. |
| [73] | alpha(1A)-Adrenergic Receptor | rs1383914 and rs1048101 SNPs are associated with higher FM impact questionnaire scores; rs574584 is associated with a higher FM impact questionnaire score, increased stiffness, and increased fatigue. |
| [74] | CCL11 | Associated with increased susceptibility to FM due to higher levels of plasma chemokines and an increased inflammatory response. |
| [75] | VNTR | Encodes interleukin 4. A 70 bp polymorphism at this locus is associated with a higher risk for FM. |
| [76] | C11orf40 | Associated with higher levels of inflammatory cytokines. |
| [76] | ZNF77 | Associated with higher levels of inflammatory cytokines. |
| [77] | MEFV | Encodes a protein called pyrin, which suppresses inflammation; missense mutations of this gene correlated with higher levels of plasma IL-1beta. |
| [78] | RNF123 | Encodes E3 ubiquitin-protein-ligase, which plays a role in cell cycle progression, innate immunity, and the metabolism of proteins. rs1491985 SNP is associated with an increased risk of developing FM. |
| [79,80,81] | AAT | Encodes alpha-1-antitrypsin; the PI*Z polymorphism has increased prevalence in FM patients. ¼ to 1/36 of FM patients found to have AAT deficiency. |
| [32,82,83,84] | BDNF | rs12273539 SNP is associated with susceptibility and symptoms of FM; rs7124442 and rs2049046 SNPs are associated with body mass index and anxiety symptoms of FM. rs6265 polymorphism is associated with pain catastrophizing in FM. Val66Val SNP is associated with elevated plasma CRP and body mass index. |
| [67,85] | VDR | Encodes the vitamin D receptor; Apal polymorphism and Fokl polymorphism increase the risk of developing FM in women. |
| [86] | CNR1 | Encodes the cannabis receptor; the rs6454674 SNP increases the risk for depression in FM. |
| [87] | m.2352C | Mitochondrial DNA gene, which increases the risk of developing FM. |
| [88,89,90,91] | MTHFR | Encodes a key enzyme in folate metabolism and increases the risk of developing FM. |
| [92] | EDN-1 | Encodes endothelin-1, a potent vasoconstrictor; the rs1800541 SNP is associated with higher levels of endothelin-1 and susceptibility to FM. |
| [93] | ACE I/D | Encodes angiotensin-converting enzyme; the ACE I/D polymorphism increases susceptibility to FM. |
| Study | Genes | Main Results |
|---|---|---|
| [81] | Sp1 C/EBPalpha | DNA methylation at Sp1 and C/EBPalpha correlated with widespread pain syndrome and decreased leptin expression and serum leptin levels in FM individuals. |
| [86] | GRM2 | Hypermethylation of GRM2 in FM individuals compared to healthy controls. |
| [90] | BDNF | Hypomethylation at exon 9 of the BDNF gene. BDNF levels are increased in FM individuals. |
| [91] | BDNF, NAT15, HDAC4, PRKCA, RTN1, PRKG1 | Differentially methylated in these genes in women with FM compared to healthy controls. These genes are involved in neuron differentiation/nervous system development, skeletal/organ system development, and chromatin compaction. |
| [92] | DAP3 miR2100 | Hypermethylation at these sites in FM individuals with adverse childhood experiences compared to FM individuals without adverse childhood experiences. |
| [104] | 69% of the differentially methylated genes are in the MAPK signaling pathway, regulation of the actin cytoskeleton, endocytosis, and neuroactive ligand receptor pathways. | FM individuals had 1610 differentially methylated positions compared to healthy controls. |
| [105] | MDH2, CLEC3B, HSPB6 | In a 281-twin individual epigenome-wide analysis of DNA methylation, CpG loci with significant p-values were MDH2, tetranectin, and heat shock protein beta-6. |
| Study | micro-RNAs | Targeted Genes | Main Results |
|---|---|---|---|
| [107] | micro-RNA-320a | FM individuals had significantly increased levels of micro-RNA-320-a compared to controls. Within the FM population, increased micro-RNA-320-a levels are associated with insomnia, chronic fatigue syndrome, persistent depressive disorder, and primary headache disorder. | |
| [108] | micro-RNA-320a, micro-RNA-320b, micro-RNA-142-3p | The plasma levels of micro-RNA-320a, micro-RNA-320b, and micro-RNA-142-3p are positively correlated with the symptom severity score in general health, functional status, and mental symptoms in the FM impact questionnaire in women with FM. | |
| [109] | micro-RNA-320a | FM individuals with increased levels of micro-RNA-320-a had decreased pain. | |
| [110] | microRNA-320b | There is a significant negative correlation between micro-RNA-320-b and depression scores in women with FM compared to healthy controls. | |
| [111] | micro-RNA-182-5p | Bone morphogenic protein receptor 2, interleukin 6 | Micro-RNA-182-5p targets bone morphogenic protein receptor 2 and interleukin 6 in FM individuals. |
| [112] | micro-RNA103a/107 | Micro-RNA-103a/107 is correlated with pro-inflammation and pain severity. | |
| [113] | micro-RNA103a, micro-RNA107 | Positive association between micro-RNA103a and micro-RNA-107 expressions and adaptive coping in FM individuals. | |
| [114] | micro-RNA-23a-3p, micro-RNA-1, micro-RNA-133a, micro-RNA-346, micro-RNA-139-5p, and micro-RNA-320b | micro-RNA-23a-3p, micro-RNA-1, micro-RNA-133a, micro-RNA-346, micro-RNA-139-5p, and micro-RNA-320b were downregulated in FM individuals. | |
| [115] | micro-RNA-let-7d | Insulin-like growth factor-1 | Higher levels of micro-RNA-let-07d correlated with reduced small nerve fiber density in FM individuals. Insulin-like growth factor-1 is a downstream target of micro-RNA-let-07d that may lead to small nerve fiber impairment. |
| [117] | hsa-micro-RNA223-3p, hsa-micro-R451a, hsa-micro-RNA338-3p, hsa-micro-RNA143-3p, hsa-micro-RNA145-5p, and hsa-micro-RNA-21-5p | hsa-micro-RNA223-3p, hsa-micro-R451a, hsa-micro-RNA338-3p, hsa-micro-RNA143-3p, hsa-micro-RNA145-5p, and hsa-micro-RNA-21-5p are significantly downregulated in FM individuals. | |
| [118] | micro-RNA-21-5p, micro-RNA-145-5p, micro-RNA-29a-3p, micro-RNA-99b-5p, micro-RNA-125b-5p, micro-RNA-23a-3p, 23b-3p, micro-RNA-195-5p, and micro-RNA-223-3p | Expressions of micro-RNA-21-5p, micro-RNA-145-5p, micro-RNA-29a-3p, micro-RNA-99b-5p, micro-RNA-125b-5p, micro-RNA-23a-3p, 23b-3p, micro-RNA-195-5p, and micro-RNA-223-3p are significantly lower in FM individuals compared with healthy controls. MiR-145 is associated with pain and fatigue. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ovrom, E.A.; Mostert, K.A.; Khakhkhar, S.; McKee, D.P.; Yang, P.; Her, Y.F. A Comprehensive Review of the Genetic and Epigenetic Contributions to the Development of Fibromyalgia. Biomedicines 2023, 11, 1119. https://doi.org/10.3390/biomedicines11041119
Ovrom EA, Mostert KA, Khakhkhar S, McKee DP, Yang P, Her YF. A Comprehensive Review of the Genetic and Epigenetic Contributions to the Development of Fibromyalgia. Biomedicines. 2023; 11(4):1119. https://doi.org/10.3390/biomedicines11041119
Chicago/Turabian StyleOvrom, Erik A., Karson A. Mostert, Shivani Khakhkhar, Daniel P. McKee, Padao Yang, and Yeng F. Her. 2023. "A Comprehensive Review of the Genetic and Epigenetic Contributions to the Development of Fibromyalgia" Biomedicines 11, no. 4: 1119. https://doi.org/10.3390/biomedicines11041119
APA StyleOvrom, E. A., Mostert, K. A., Khakhkhar, S., McKee, D. P., Yang, P., & Her, Y. F. (2023). A Comprehensive Review of the Genetic and Epigenetic Contributions to the Development of Fibromyalgia. Biomedicines, 11(4), 1119. https://doi.org/10.3390/biomedicines11041119