
Neuromodulation for Peripheral Nerve Regeneration: Systematic Review of Mechanisms and In Vivo Highlights

Abstract
:1. Introduction
2. Materials and Methods
2.1. Search Strategy
2.2. Study Selection
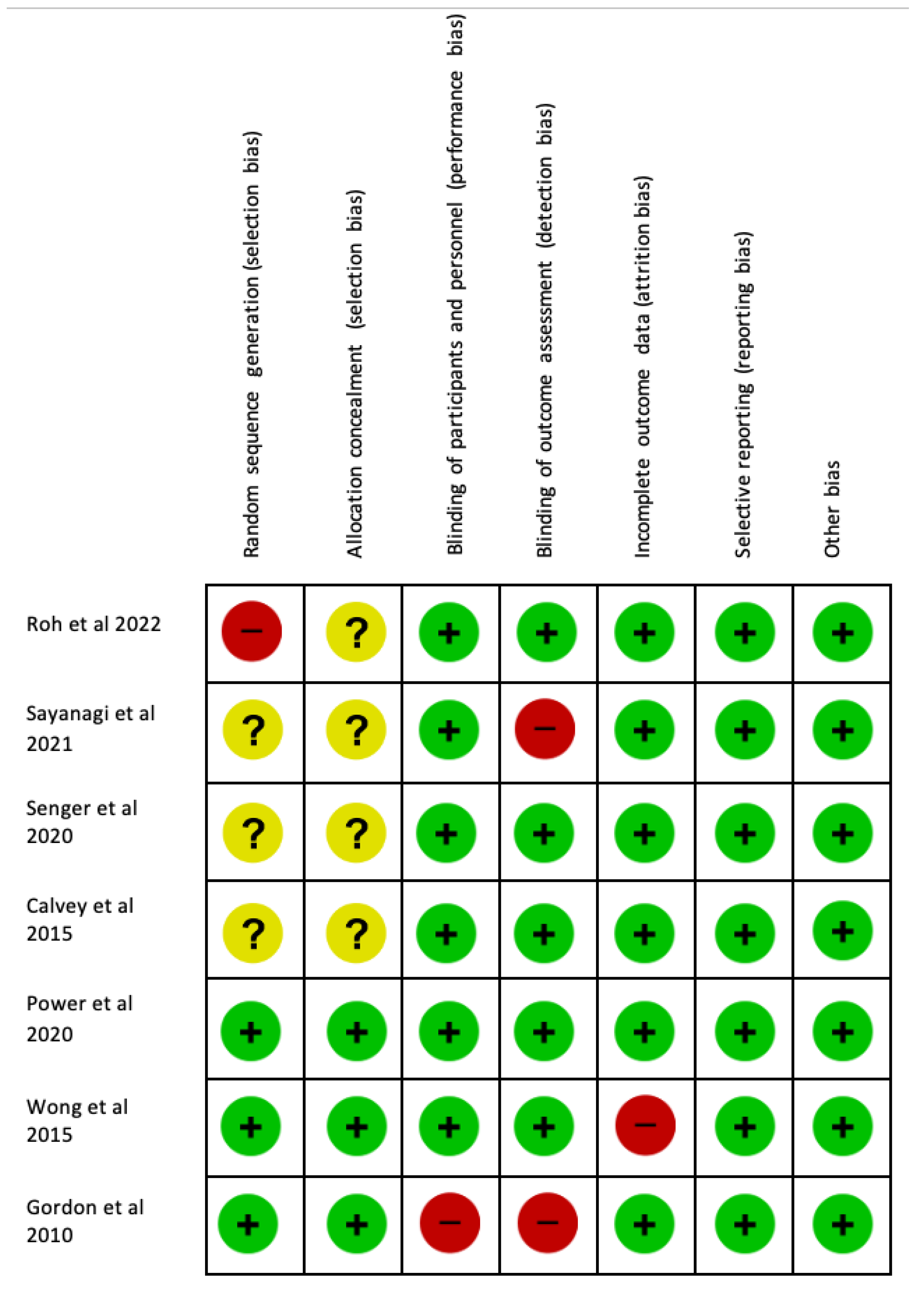
2.3. Bias Assessment
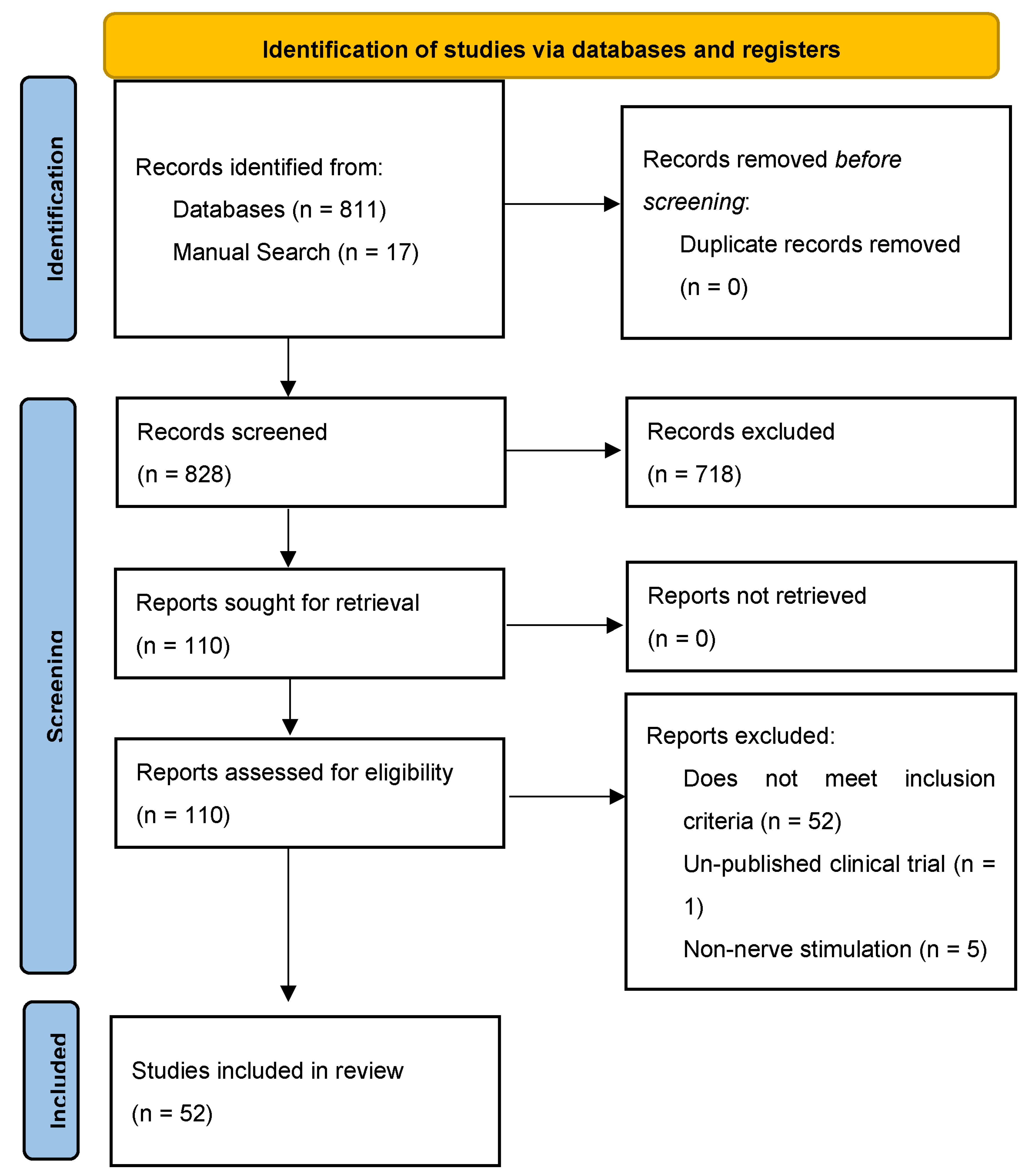
3. Results
3.1. Search Results
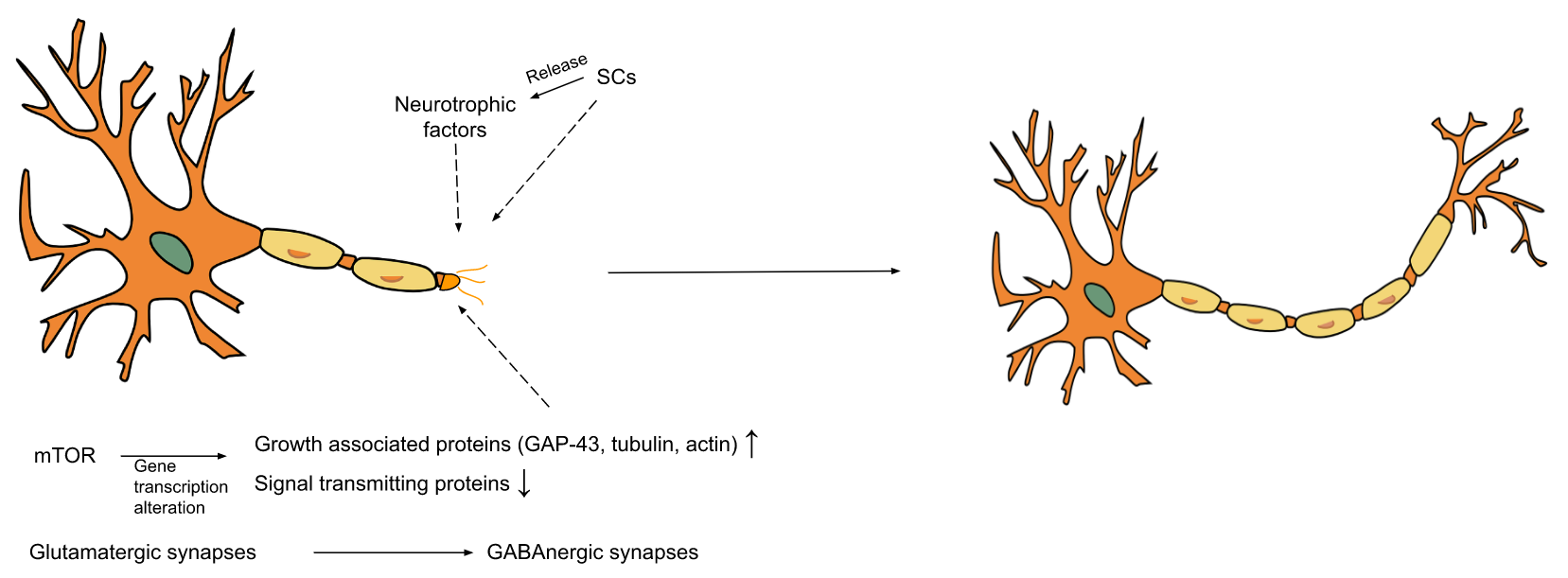
3.2. Mechanisms
3.2.1. Biological Mechanisms
3.2.2. ES Mechanisms
3.2.3. Other Neuromodulation Mechanisms
3.3. In Vivo Highlights
3.4. Bias Assessment
4. Discussion
5. Importance of Animal Models
6. Future Directions
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soendenbroe, C.; Andersen, J.L.; Mackey, A.L. Muscle-Nerve Communication and the Molecular Assessment of Human Skeletal Muscle Denervation with Aging. Am. J. Physiol.-Cell Physiol. 2021, 321, C317–C329. [Google Scholar] [CrossRef] [PubMed]
- Narayan, S.K.; Arumugam, M.; Chittoria, R. Outcome of Human Peripheral Nerve Repair Interventions Using Conduits: A Systematic Review. J. Neurol. Sci. 2019, 396, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Agata, V.; Trovato, B.; Roggio, F.; Castorina, A.; Vecchio, M.; Di Rosa, M.; Musumeci, G. The Role of Exercise on Peripheral Nerve Regeneration: From Animal Model to Clinical Application. Heliyon 2021, 7, e08281. [Google Scholar] [CrossRef] [PubMed]
- Osborne, N.R.; Anastakis, D.J.; Davis, K.D. Peripheral Nerve Injuries, Pain, and Neuroplasticity. J. Hand Ther. 2018, 31, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Busch, C.; Smith, O.; Weaver, T.; Vallabh, J.; Abd-Elsayed, A. Peripheral Nerve Stimulation for Lower Extremity Pain. Biomedicines 2022, 10, 1666. [Google Scholar] [CrossRef]
- Ring, D. Symptoms and Disability after Major Peripheral Nerve Injury. Hand Clin. 2013, 29, 421–425. [Google Scholar] [CrossRef]
- Padovano, W.M.; Dengler, J.; Patterson, M.M.; Yee, A.; Snyder-Warwick, A.K.; Wood, M.D.; Moore, A.M.; Mackinnon, S.E. Incidence of Nerve Injury after Extremity Trauma in the United States. Hand 2022, 17, 615–623. [Google Scholar] [CrossRef]
- Gordon, T. Peripheral Nerve Regeneration and Muscle Reinnervation. Int. J. Mol. Sci. 2020, 21, 8652. [Google Scholar] [CrossRef]
- Rau, C.-S.; Kuo, P.-J.; Wu, S.-C.; Huang, L.-H.; Lu, T.-H.; Wu, Y.-C.; Wu, C.-J.; Lin, C.-W.; Tsai, C.-W.; Hsieh, C.-H. Enhanced Nerve Regeneration by Exosomes Secreted by Adipose-Derived Stem Cells with or without FK506 Stimulation. Int. J. Mol. Sci. 2021, 22, 8545. [Google Scholar] [CrossRef]
- Char, S.; Jin, M.Y.; Francio, V.T.; Hussain, N.; Wang, E.J.; Morsi, M.; Orhurhu, V.; Prokop, L.J.; Fink, A.; D’Souza, R.S. Implantable Peripheral Nerve Stimulation for Peripheral Neuropathic Pain: A Systematic Review of Prospective Studies. Biomedicines 2022, 10, 2606. [Google Scholar] [CrossRef]
- Chu, X.-L.; Song, X.-Z.; Li, Q.; Li, Y.-R.; He, F.; Gu, X.-S.; Ming, D. Basic Mechanisms of Peripheral Nerve Injury and Treatment via Electrical Stimulation. Neural Regen. Res. 2022, 17, 2185–2193. [Google Scholar]
- Luo, L.; He, Y.; Jin, L.; Zhang, Y.; Guastaldi, F.P.; Albashari, A.A.; Hu, F.; Wang, X.; Wang, L.; Xiao, J. Application of Bioactive Hydrogels Combined with Dental Pulp Stem Cells for the Repair of Large Gap Peripheral Nerve Injuries. Bioact. Mater. 2021, 6, 638–654. [Google Scholar] [CrossRef]
- Gordon, T. Nerve Regeneration: Understanding Biology and Its Influence on Return of Function after Nerve Transfers. Hand Clin. 2016, 32, 103–117. [Google Scholar] [CrossRef]
- Abd-Elsayed, A. Wireless Peripheral Nerve Stimulation for Treatment of Peripheral Neuralgias. Neuromodulation Technol. Neural Interface 2020, 23, 827–830. [Google Scholar] [CrossRef]
- Fiala, K.J.; Kim, R.B.; Martens, J.M.; Abd-Elsayed, A. Lumbar Level Peripheral Nerve Stimulation for Low Back Pain. Ochsner J. 2022, 22, 265–272. [Google Scholar] [CrossRef]
- Gildenberg, P.L. History of Electrical Neuromodulation for Chronic Pain. Pain Med. 2006, 7, S7–S13. [Google Scholar] [CrossRef] [Green Version]
- Strand, N.H.; D’Souza, R.; Wie, C.; Covington, S.; Maita, M.; Freeman, J.; Maloney, J. Mechanism of Action of Peripheral Nerve Stimulation. Curr. Pain Headache Rep. 2021, 25, 47. [Google Scholar] [CrossRef]
- Kent, A.R.; Min, X.; Hogan, Q.H.; Kramer, J.M. Mechanisms of Dorsal Root Ganglion Stimulation in Pain Suppression: A Computational Modeling Analysis. Neuromodulation Technol. Neural Interface 2018, 21, 234–246. [Google Scholar] [CrossRef]
- Abd-Elsayed, A.; D’Souza, R.S. Peripheral Nerve Stimulation: The Evolution in Pain Medicine. Biomedicines 2022, 10, 18. [Google Scholar] [CrossRef]
- Labaran, L.; Jain, N.; Puvanesarajah, V.; Jain, A.; Buchholz, A.L.; Hassanzadeh, H. A Retrospective Database Review of the Indications, Complications, and Incidence of Subsequent Spine Surgery in 12,297 Spinal Cord Stimulator Patients. Neuromodulation Technol. Neural Interface 2020, 23, 634–638. [Google Scholar] [CrossRef]
- Scarfo, K.-A.; Demian, P.S.; Strand, N.; Hunter, C.; Deer, T.R. Introduction to Dorsal Root Ganglion Stimulation an Overview of the Field. In Neuromodulation Techniques for Pain Treatment: A Step-by-Step Guide to Interventional Procedures and Managing Complications; Freitas, T.d.S., Assumpcao de Monaco, B., Golovac, S., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 81–90. ISBN 978-3-030-84778-4. [Google Scholar]
- D’Souza, R.S.; Kubrova, E.; Her, Y.F.; Barman, R.A.; Smith, B.J.; Alvarez, G.M.; West, T.E.; Abd-Elsayed, A. Dorsal Root Ganglion Stimulation for Lower Extremity Neuropathic Pain Syndromes: An Evidence-Based Literature Review. Adv. Ther. 2022, 39, 4440–4473. [Google Scholar] [CrossRef] [PubMed]
- Strand, N.; D’Souza, R.S.; Hagedorn, J.M.; Pritzlaff, S.; Sayed, D.; Azeem, N.; Abd-Elsayed, A.; Escobar, A.; Huntoon, M.A.; Lam, C.M. Evidence-Based Clinical Guidelines from the American Society of Pain and Neuroscience for the Use of Implantable Peripheral Nerve Stimulation in the Treatment of Chronic Pain. J. Pain Res. 2022, 15, 2483–2504. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.Y.; D’Souza, R.S.; Abd-Elsayed, A.A. Efficacy of Neuromodulation Interventions for the Treatment of Sexual Dysfunction: A Systematic Review. Neuromodulation Technol. Neural Interface 2022. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Sun, Z.; Wu, J.; Rana, M.; Garza, J.; Zhu, A.C.; Chakravarthy, K.V.; Abd-Elsayed, A.; Rosenquist, E.; Basi, H. Peripheral Nerve Stimulation in Pain Management: A Systematic Review. Pain Physician 2021, 24, E131. [Google Scholar] [PubMed]
- Zuo, K.J.; Gordon, T.; Chan, K.M.; Borschel, G.H. Electrical Stimulation to Enhance Peripheral Nerve Regeneration: Update in Molecular Investigations and Clinical Translation. Exp. Neurol. 2020, 332, 113397. [Google Scholar] [CrossRef]
- ElAbd, R.; Alabdulkarim, A.; AlSabah, S.; Hazan, J.; Alhalabi, B.; Thibaudeau, S. Role of Electrical Stimulation in Peripheral Nerve Regeneration: A Systematic Review. Plast. Reconstr. Surg. Glob. Open 2022, 10, e4115. [Google Scholar] [CrossRef]
- Jo, S.; Pan, D.; Halevi, A.E.; Roh, J.; Schellhardt, L.; Hunter RA, D.A.; Snyder-Warwick, A.K.; Moore, A.M.; Mackinnon, S.E.; Wood, M.D. Comparing Electrical Stimulation and Tacrolimus (FK506) to Enhance Treating Nerve Injuries. Muscle Nerve 2019, 60, 629–636. [Google Scholar] [CrossRef]
- Page, M.J.; Shamseer, L.; Tricco, A.C. Registration of Systematic Reviews in PROSPERO: 30,000 Records and Counting. Syst. Rev. 2018, 7, 32. [Google Scholar] [CrossRef]
- Higgins, J.P.; Altman, D.G.; Gøtzsche, P.C.; Jüni, P.; Moher, D.; Oxman, A.D.; Savović, J.; Schulz, K.F.; Weeks, L.; Sterne, J.A. The Cochrane Collaboration’s Tool for Assessing Risk of Bias in Randomised Trials. Bmj 2011, 343, d5928. [Google Scholar] [CrossRef] [Green Version]
- English, A.W.; Wilhelm, J.C.; Ward, P.J. Exercise, Neurotrophins, and Axon Regeneration in the PNS. Physiology 2014, 29, 437–445. [Google Scholar] [CrossRef] [Green Version]
- Gordon, T.; English, A.W. Strategies to Promote Peripheral Nerve Regeneration: Electrical Stimulation and/or Exercise. Eur. J. Neurosci. 2016, 43, 336–350. [Google Scholar] [CrossRef] [Green Version]
- van Niekerk, E.A.; Tuszynski, M.H.; Lu, P.; Dulin, J.N. Molecular and Cellular Mechanisms of Axonal Regeneration after Spinal Cord Injury. Mol. Cell. Proteom. 2016, 15, 394–408. [Google Scholar] [CrossRef] [Green Version]
- Zigmond, R.E.; Echevarria, F.D. Macrophage Biology in the Peripheral Nervous System after Injury. Prog. Neurobiol. 2019, 173, 102–121. [Google Scholar] [CrossRef]
- Willand, M.P.; Nguyen, M.-A.; Borschel, G.H.; Gordon, T. Electrical Stimulation to Promote Peripheral Nerve Regeneration. Neurorehabil. Neural Repair 2016, 30, 490–496. [Google Scholar] [CrossRef] [Green Version]
- Gordon, T. Electrical Stimulation to Enhance Axon Regeneration after Peripheral Nerve Injuries in Animal Models and Humans. Neurotherapeutics 2016, 13, 295–310. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.J.; Kim, H.W.; Gonzalez-Cano, R.; Choi, J.; Back, S.K.; Roh, S.E.; Johnson, E.; Gabriac, M.; Kim, M.-S.; Lee, J. Natural Killer Cells Degenerate Intact Sensory Afferents Following Nerve Injury. Cell 2019, 176, 716–728. [Google Scholar] [CrossRef] [Green Version]
- Richner, M.; Ulrichsen, M.; Elmegaard, S.L.; Dieu, R.; Pallesen, L.T.; Vaegter, C.B. Peripheral Nerve Injury Modulates Neurotrophin Signaling in the Peripheral and Central Nervous System. Mol. Neurobiol. 2014, 50, 945–970. [Google Scholar] [CrossRef]
- Ward, P.J.; Davey, R.A.; Zajac, J.D.; English, A.W. Neuronal Androgen Receptor Is Required for Activity Dependent Enhancement of Peripheral Nerve Regeneration. Dev. Neurobiol. 2021, 81, 411–423. [Google Scholar] [CrossRef]
- Thompson, N.J.; Sengelaub, D.R.; English, A.W. Enhancement of Peripheral Nerve Regeneration Due to Treadmill Training and Electrical Stimulation Is Dependent on Androgen Receptor Signaling. Dev. Neurobiol. 2014, 74, 531–540. [Google Scholar] [CrossRef]
- Singh, B.; Krishnan, A.; Micu, I.; Koshy, K.; Singh, V.; Martinez, J.; Koshy, D.; Xu, F.; Chandrasekhar, A.; Dalton, C. Peripheral Neuron Plasticity Is Enhanced by Brief Electrical Stimulation and Overrides Attenuated Regrowth in Experimental Diabetes. Neurobiol. Dis. 2015, 83, 134–151. [Google Scholar] [CrossRef]
- Zhu, H.; Xue, C.; Yao, M.; Wang, H.; Zhang, P.; Qian, T.; Zhou, S.; Li, S.; Yu, B.; Wang, Y. MiR-129 Controls Axonal Regeneration via Regulating Insulin-like Growth Factor-1 in Peripheral Nerve Injury. Cell Death Dis. 2018, 9, 720. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, Y.; Lu, L.; Hu, X.; Luo, Z. Electrical Stimulation Accelerates Nerve Regeneration and Functional Recovery in Delayed Peripheral Nerve Injury in Rats. Eur. J. Neurosci. 2013, 38, 3691–3701. [Google Scholar] [CrossRef] [PubMed]
- English, A.W.; Liu, X.; Mistretta, O.C.; Ward, P.J.; Ye, K. Asparagine Endopeptidase (δ Secretase), an Enzyme Implicated in Alzheimer’s Disease Pathology, Is an Inhibitor of Axon Regeneration in Peripheral Nerves. Eneuro 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Pei, B.; Zi, J.; Wu, L.; Zhang, C.; Chen, Y. Pulsed Electrical Stimulation Protects Neurons in the Dorsal Root and Anterior Horn of the Spinal Cord after Peripheral Nerve Injury. Neural Regen. Res. 2015, 10, 1650. [Google Scholar] [PubMed]
- Zhou, S.; Gao, R.; Hu, W.; Qian, T.; Wang, N.; Ding, G.; Ding, F.; Yu, B.; Gu, X. MiR-9 Inhibits Schwann Cell Migration by Targeting Cthrc1 Following Sciatic Nerve Injury. J. Cell Sci. 2014, 127, 967–976. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Yamamoto, T.; Honjo, K.; Ichioka, H.; Oseko, F.; Kishida, T.; Mazda, O.; Kanamura, N. Electrical Stimulation with Periodic Alternating Intervals Stimulates Neuronal Cells to Produce Neurotrophins and Cytokines through Activation of Mitogen-Activated Protein Kinase Pathways. Eur. J. Oral Sci. 2015, 123, 403–408. [Google Scholar] [CrossRef]
- Yi, S.; Wang, S.; Zhao, Q.; Yao, C.; Gu, Y.; Liu, J.; Gu, X.; Li, S. MiR-Sc3, a Novel MicroRNA, Promotes Schwann Cell Proliferation and Migration by Targeting Astn1. Cell Transplant. 2016, 25, 973–982. [Google Scholar] [CrossRef] [Green Version]
- Wakatsuki, S.; Araki, T.; Sehara-Fujisawa, A. Neuregulin-1/Glial Growth Factor Stimulates S Chwann Cell Migration by Inducing A5 Β1 Integrin–ErbB2–Focal Adhesion Kinase Complex Formation. Genes Cells 2014, 19, 66–77. [Google Scholar] [CrossRef]
- Gu, X.; Fu, J.; Bai, J.; Wang, J.; Pan, W.; Zhang, C. Low-Frequency Electrical Stimulation Induces the Proliferation and Differentiation of Peripheral Blood Stem Cells into Schwann Cells. Am. J. Med. Sci. 2015, 349, 157–161. [Google Scholar] [CrossRef]
- Stassart, R.M.; Fledrich, R.; Velanac, V.; Brinkmann, B.G.; Schwab, M.H.; Meijer, D.; Sereda, M.W.; Nave, K.-A. A Role for Schwann Cell–Derived Neuregulin-1 in Remyelination. Nat. Neurosci. 2013, 16, 48–54. [Google Scholar] [CrossRef]
- Chen, T.-J.; Kukley, M. Glutamate Receptors and Glutamatergic Signalling in the Peripheral Nerves. Neural Regen. Res. 2020, 15, 438. [Google Scholar]
- Alvarez, F.J.; Rotterman, T.M.; Akhter, E.T.; Lane, A.R.; English, A.W.; Cope, T.C. Synaptic Plasticity on Motoneurons after Axotomy: A Necessary Change in Paradigm. Front. Mol. Neurosci. 2020, 13, 68. [Google Scholar] [CrossRef]
- Liu, Q.-Y.; Miao, Y.; Wang, X.-H.; Wang, P.; Cheng, Z.-C.; Qian, T.-M. Increased Levels of MiR-3099 Induced by Peripheral Nerve Injury Promote Schwann Cell Proliferation and Migration. Neural Regen. Res. 2019, 14, 525. [Google Scholar]
- Wang, D.; Chen, Y.; Liu, M.; Cao, Q.; Wang, Q.; Zhou, S.; Wang, Y.; Mao, S.; Gu, X.; Luo, Z. The Long Noncoding RNA Arrl1 Inhibits Neurite Outgrowth by Functioning as a Competing Endogenous RNA during Neuronal Regeneration in Rats. J. Biol. Chem. 2020, 295, 8374–8386. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.; Wang, Q.; Wang, Y.; Wu, J.; Cao, X.; Lu, Y.; Chen, Y.; Feng, W.; Gu, X.; Dun, X. Loc680254 Regulates Schwann Cell Proliferation through Psrc1 and Ska1 as a MicroRNA Sponge Following Sciatic Nerve Injury. Glia 2021, 69, 2391–2403. [Google Scholar] [CrossRef]
- Yao, C.; Wang, Y.; Zhang, H.; Feng, W.; Wang, Q.; Shen, D.; Qian, T.; Liu, F.; Mao, S.; Gu, X. LncRNA TNXA-PS1 Modulates Schwann Cells by Functioning as a Competing Endogenous RNA Following Nerve Injury. J. Neurosci. 2018, 38, 6574–6585. [Google Scholar] [CrossRef]
- Wang, D.; Zheng, T.; Ge, X.; Xu, J.; Feng, L.; Jiang, C.; Tao, J.; Chen, Y.; Liu, X.; Yu, B. Unfolded Protein Response-Induced Expression of Long Noncoding RNA Ngrl1 Supports Peripheral Axon Regeneration by Activating the PI3K-Akt Pathway. Exp. Neurol. 2022, 352, 114025. [Google Scholar] [CrossRef]
- Gordon, T. Nerve Regeneration in the Peripheral and Central Nervous Systems. J. Physiol. 2016, 594, 3517. [Google Scholar] [CrossRef] [Green Version]
- Wagstaff, L.J.; Gomez-Sanchez, J.A.; Fazal, S.V.; Otto, G.W.; Kilpatrick, A.M.; Michael, K.; Wong, L.Y.; Ma, K.H.; Turmaine, M.; Svaren, J. Failures of Nerve Regeneration Caused by Aging or Chronic Denervation Are Rescued by Restoring Schwann Cell C-Jun. Elife 2021, 10, e62232. [Google Scholar] [CrossRef]
- Wood, M.D.; Kemp, S.W.; Weber, C.; Borschel, G.H.; Gordon, T. Outcome Measures of Peripheral Nerve Regeneration. Ann. Anat.-Anat. Anz. 2011, 193, 321–333. [Google Scholar] [CrossRef]
- Colley, K.J.; Kitajima, K.; Sato, C. Polysialic Acid: Biosynthesis, Novel Functions and Applications. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 498–532. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Qian, T.; Wang, Y.; Zhou, S.; Ding, G.; Ding, F.; Gu, X. MiR-182 Inhibits Schwann Cell Proliferation and Migration by Targeting FGF9 and NTM, Respectively at an Early Stage Following Sciatic Nerve Injury. Nucleic Acids Res. 2012, 40, 10356–10365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, S.; Yuan, Y.; Chen, Q.; Wang, X.; Gong, L.; Liu, J.; Gu, X.; Li, S. Regulation of Schwann Cell Proliferation and Migration by MiR-1 Targeting Brain-Derived Neurotrophic Factor after Peripheral Nerve Injury. Sci. Rep. 2016, 6, 29121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhang, R.; Yuan, Y.; Yi, S.; Chen, Q.; Gong, L.; Liu, J.; Ding, F.; Cao, Z.; Gu, X. MiR-340 Regulates Fibrinolysis and Axon Regrowth Following Sciatic Nerve Injury. Mol. Neurobiol. 2017, 54, 4379–4389. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Chen, C.; Yi, S.; Wang, S.; Gong, L.; Liu, J.; Gu, X.; Zhao, Q.; Li, S. MiR-Sc8 Inhibits Schwann Cell Proliferation and Migration by Targeting Egfr. PLoS ONE 2015, 10, e0145185. [Google Scholar] [CrossRef]
- Huang, J.; Ye, Z.; Hu, X.; Lu, L.; Luo, Z. Electrical Stimulation Induces Calcium-dependent Release of NGF from Cultured Schwann Cells. Glia 2010, 58, 622–631. [Google Scholar] [CrossRef]
- Luo, B.; Huang, J.; Lu, L.; Hu, X.; Luo, Z.; Li, M. Electrically Induced Brain-derived Neurotrophic Factor Release from Schwann Cells. J. Neurosci. Res. 2014, 92, 893–903. [Google Scholar] [CrossRef]
- Vingren, J.L.; Kraemer, W.J.; Ratamess, N.A.; Anderson, J.M.; Volek, J.S.; Maresh, C.M. Testosterone Physiology in Resistance Exercise and Training. Sports Med. 2010, 40, 1037–1053. [Google Scholar] [CrossRef]
- Ashida, Y.; Himori, K.; Tatebayashi, D.; Yamada, R.; Ogasawara, R.; Yamada, T. Effects of Contraction Mode and Stimulation Frequency on Electrical Stimulation-Induced Skeletal Muscle Hypertrophy. J. Appl. Physiol. 2018, 124, 341–348. [Google Scholar] [CrossRef]
- Juretic, N.; Jorquera, G.; Caviedes, P.; Jaimovich, E.; Riveros, N. Electrical Stimulation Induces Calcium-Dependent up-Regulation of Neuregulin-1β in Dystrophic Skeletal Muscle Cell Lines. Cell. Physiol. Biochem. 2012, 29, 919–930. [Google Scholar] [CrossRef]
- Quan, X.; Huang, L.; Yang, Y.; Ma, T.; Liu, Z.; Ge, J.; Huang, J.; Luo, Z. Potential Mechanism of Neurite Outgrowth Enhanced by Electrical Stimulation: Involvement of MicroRNA-363-5p Targeting DCLK1 Expression in Rat. Neurochem. Res. 2017, 42, 513–525. [Google Scholar] [CrossRef]
- Roh, J.; Schellhardt, L.; Keane, G.C.; Hunter, D.A.; Moore, A.M.; Snyder-Warwick, A.K.; Mackinnon, S.E.; Wood, M.D. Short-Duration, Pulsatile, Electrical Stimulation Therapy Accelerates Axon Regeneration and Recovery Following Tibial Nerve Injury and Repair in Rats. Plast. Reconstr. Surg. 2022, 149, 681e–690e. [Google Scholar] [CrossRef]
- Sayanagi, J.; Acevedo-Cintrón, J.A.; Pan, D.; Schellhardt, L.; Hunter, D.A.; Snyder-Warwick, A.K.; Mackinnon, S.E.; Wood, M.D. Brief Electrical Stimulation Accelerates Axon Regeneration and Promotes Recovery Following Nerve Transection and Repair in Mice. JBJS 2021, 103, e80. [Google Scholar] [CrossRef]
- Senger, J.-L.B.; Chan, K.M.; Webber, C.A. Conditioning Electrical Stimulation Is Superior to Postoperative Electrical Stimulation, Resulting in Enhanced Nerve Regeneration and Functional Recovery. Exp. Neurol. 2020, 325, 113147. [Google Scholar] [CrossRef]
- Calvey, C.; Zhou, W.; Stakleff, K.S.; Sendelbach-Sloan, P.; Harkins, A.B.; Lanzinger, W.; Willits, R.K. Short-Term Electrical Stimulation to Promote Nerve Repair and Functional Recovery in a Rat Model. J. Hand Surg. 2015, 40, 314–322. [Google Scholar] [CrossRef]
- Power, H.A.; Morhart, M.J.; Olson, J.L.; Chan, K.M. Postsurgical Electrical Stimulation Enhances Recovery Following Surgery for Severe Cubital Tunnel Syndrome: A Double-Blind Randomized Controlled Trial. Neurosurgery 2020, 86, 769–777. [Google Scholar] [CrossRef]
- Wong, J.N.; Olson, J.L.; Morhart, M.J.; Chan, K.M. Electrical Stimulation Enhances Sensory Recovery: A Randomized Controlled Trial. Ann. Neurol. 2015, 77, 996–1006. [Google Scholar] [CrossRef]
- Gordon, T.; Amirjani, N.; Edwards, D.C.; Chan, K.M. Brief Post-Surgical Electrical Stimulation Accelerates Axon Regeneration and Muscle Reinnervation without Affecting the Functional Measures in Carpal Tunnel Syndrome Patients. Exp. Neurol. 2010, 223, 192–202. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Sulaiman, W.; Gordon, T. Neurobiology of Peripheral Nerve Injury, Regeneration, and Functional Recovery: From Bench Top Research to Bedside Application. Ochsner J. 2013, 13, 100–108. [Google Scholar]
- Gordon, T.; Borschel, G.H. The Use of the Rat as a Model for Studying Peripheral Nerve Regeneration and Sprouting after Complete and Partial Nerve Injuries. Exp. Neurol. 2017, 287, 331–347. [Google Scholar] [CrossRef]
- Günter, C.; Delbeke, J.; Ortiz-Catalan, M. Safety of Long-Term Electrical Peripheral Nerve Stimulation: Review of the State of the Art. J. Neuroeng. Rehabil. 2019, 16, 13. [Google Scholar] [CrossRef] [Green Version]
- Senger, J.L.B.; Verge, V.M.K.; Macandili, H.S.J.; Olson, J.L.; Chan, K.M.; Webber, C.A. Electrical Stimulation as a Conditioning Strategy for Promoting and Accelerating Peripheral Nerve Regeneration. Exp. Neurol. 2018, 302, 75–84. [Google Scholar] [CrossRef]
- Friedman, L.M.; Furberg, C.D.; DeMets, D.L.; Reboussin, D.M.; Granger, C.B. Fundamentals of Clinical Trials; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 3-319-18539-X. [Google Scholar]
- Jaggi, A.S.; Jain, V.; Singh, N. Animal Models of Neuropathic Pain. Fundam. Clin. Pharmacol. 2011, 25, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.-P.; Shen, Y.-J.; Qiu, Y.-Q.; Li, J.; Shen, Y.-D.; Wang, X.-M.; Cong, M.; He, Q.-R.; Chen, Q.-Z.; Jiang, S. Comparative Effects of Implanted Electrodes with Differing Contact Patterns on Peripheral Nerve Regeneration and Functional Recovery. Neurosci. Res. 2019, 145, 22–29. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wallerian Degeneration |
|---|
|
| Regeneration |
|
| Intervention | Key Mechanisms |
|---|---|
| ES | |
| SCS |
|
| DRG-S |
| Author | Study Population | Injury Type | Stimulation Settings (Location) | Arms | Outcomes |
|---|---|---|---|---|---|
| Roh et al., 2022 [73] | 39 rats | Tibial nerve transection Ψ | 100 µs, 0.5 mA, 16 Hz (2 mm proximal to the cut/repair site and proximal to injury site) | 10 min ES (n = 13) vs. 60 min ES (n = 13) vs. no ES control (n = 13) | At 2 weeks, the 10 min ES group had greater axon outgrowth than the 60 min ES group which had greater outgrowth than the no ES groups. At 52 days, 10 min and 60 min ES groups both had similar TFI scores that were significantly greater than the control. The control group had a significantly greater number of myelinated axons than the 10 min ES group. |
| Sayanagi et al., 2021 [74] | 99 mice | Sciatic nerve transection Ψ | 0.5 mA, 16 Hz (2 mm proximal to the repair site) | 10 min ES (n = 33) vs. 60 min ES (n = 33) vs. no ES control (n = 33) | At 15 days, both ES groups had a significantly greater number of labeled motoneurons and myelinated axons than the control. At 56 days, both ES groups also outperformed the control on the grid walking test and on mechanical sensitivity. No significant differences in cold sensitivity were found for any of the groups. |
| Senger et al., 2020 [75] | 122 rats | Tibial nerve transection Ψ | 0.1 ms, 20 Hz (tibial nerve) | CES Ω (n = NR) vs. PES vs. CES + PES vs. control | At 1 week, CES group had significantly greater length and number of axons regenerated than all other groups. At 6 weeks, CES significantly outperformed all groups in sensory and motor recovery. |
| Calvey et al., 2015 [76] | 41 rats | 10-mm Sciatic nerve injury ± | 24 mV, ∼1.5 μA (proximal and distal nerve segments) | 10 min ES (n = 10) vs. 60 min ES (n = 11) vs. no ES control (n = 10) vs. Isograft control (n = 10) | SFI and extensor potential thrust at week 12 were greatest in 10 min ES group. The number of nerve fibers at the midline of the conduit was greatest in isograft control and at 2 mm distal to the repair conduit was equally greatest for both ES groups and isograft control |
| Power et al., 2020 [77] | 31 humans | Cubital tunnel syndrome * | <30 V, 0.1 ms, 20 Hz (ulnar nerve proximal to the site of compression) | 60 min ES (n = 20) vs. no ES control (n = 11) | At 3 years, the ES group demonstrated significantly higher MUNE and key pinch strength than control. Other functional and physiological outcomes were also significantly improved in the ES group compared to the control. |
| Wong et al., 2015 [78] | 36 humans | Complete digital nerve transection Ψ | <30 V, 0.1–0.4 ms, 20 Hz (proximal to surgical site) | 60 min ES (n = 16) vs. sham (n = 16) | ES group reported greater restoration of CDT and DASH scores. Based on the MRC Modified Highet Scale, 86.7% of the ES group experienced normal recovery in tactile discrimination and pressure detection as compared to 43.8% of the sham group. Sham group recovery plateaued at 3–4 months. |
| Gordon et al., 2010 [79] | 21 humans | Carpal tunnel syndrome * | 4–6 V, 0.1–0.8 ms, 20 Hz (medial nerve above the site of compression) | 60 min ES (n = 11) vs. no ES control (n = 10) | MUNE, terminal motor latency, and sensory nerve conduction significantly improved for the ES group but not control. MUNE of the ES group was not statistically different from a healthy hand. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, M.Y.; Weaver, T.E.; Farris, A.; Gupta, M.; Abd-Elsayed, A. Neuromodulation for Peripheral Nerve Regeneration: Systematic Review of Mechanisms and In Vivo Highlights. Biomedicines 2023, 11, 1145. https://doi.org/10.3390/biomedicines11041145
Jin MY, Weaver TE, Farris A, Gupta M, Abd-Elsayed A. Neuromodulation for Peripheral Nerve Regeneration: Systematic Review of Mechanisms and In Vivo Highlights. Biomedicines. 2023; 11(4):1145. https://doi.org/10.3390/biomedicines11041145
Chicago/Turabian StyleJin, Max Y., Tristan E. Weaver, Adam Farris, Mayank Gupta, and Alaa Abd-Elsayed. 2023. "Neuromodulation for Peripheral Nerve Regeneration: Systematic Review of Mechanisms and In Vivo Highlights" Biomedicines 11, no. 4: 1145. https://doi.org/10.3390/biomedicines11041145
APA StyleJin, M. Y., Weaver, T. E., Farris, A., Gupta, M., & Abd-Elsayed, A. (2023). Neuromodulation for Peripheral Nerve Regeneration: Systematic Review of Mechanisms and In Vivo Highlights. Biomedicines, 11(4), 1145. https://doi.org/10.3390/biomedicines11041145