
Genome–Environment Interactions and Psychiatric Disorders

Abstract
:1. Introduction
2. Genetic Basis of Psychiatric Disorders
3. Epigenetic Mechanisms of Gene Expression
4. Epigenetic Mechanisms and Environmental Factors
5. Environmental Factors and Psychiatric Disorders
6. Environmental Factors Acting Epigenetically in Psychiatric Disorders
6.1. Social Determinants of Mental Health Including Early Life Stress and Adversity
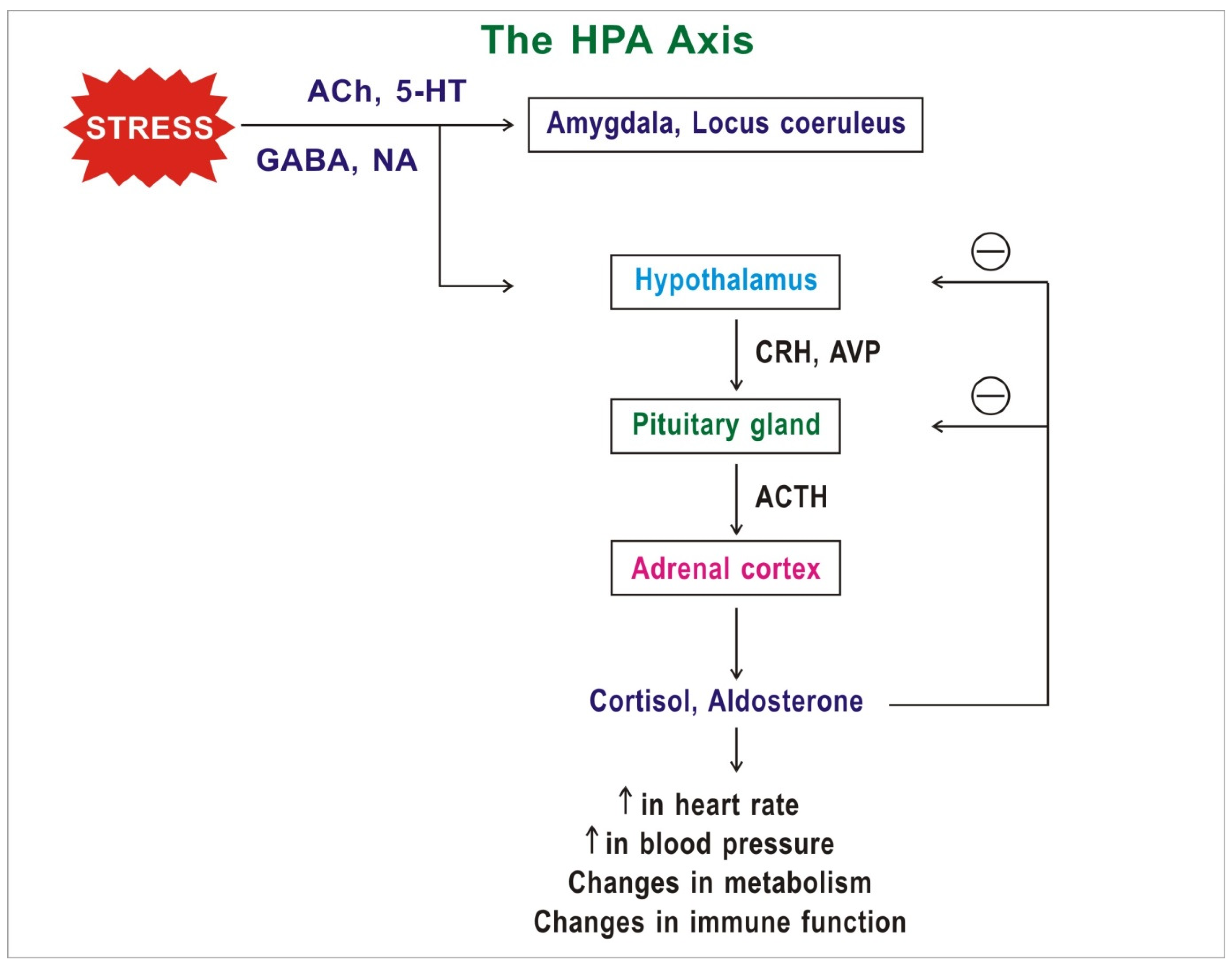
Mechanism of Action of Social Determinants of Mental Health
6.2. Maternal Prenatal Psychological Stress
6.3. Pregnancy and Birth Complications
6.4. Poverty
6.5. Migration
6.6. Urban Dwelling
6.7. Alcohol Use
6.8. Use of Substances Other Than Alcohol
6.9. Microbiota
6.10. Prenatal and Postnatal Infections
7. Reversal of Genome–Environment Interactions in the Treatment of Psychiatric Disorders
7.1. Drugs
7.2. Psychotherapy
7.3. Electroconvulsive Therapy
7.4. Physical Exercise
8. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. The ICD-11 Classification Of Mental And Behavioural Disorders: Diagnostic Criteria for Research; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Assary, E.; Vincent, J.P.; Keers, R.; Pluess, M. Gene-environment interaction and psychiatric disorders: Review and future directions. Semin. Cell Dev.Biol. 2018, 77, 133–143. [Google Scholar] [CrossRef]
- Murphy, C.M. Writing an effective review article. J. Med. Toxicol. 2012, 8, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Boland, R.; Verduin, M.L.; Ruiz, P. Kaplan & Sadock’s Synopsis of Psychiatry, 12th ed.; Wolters Kluwer: Philadelphia, PA, USA, 2022. [Google Scholar]
- Allis, C.D.; Caparros, M.-L.; Jenuwein, T.; Lachner, M.; Reinberg, D. Overview and concepts. In Epigenetics, 2nd ed.; Allis, C.D., Caparros, M.-L., Jenuwein, T., Reinberg, D., Eds.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2015; pp. 47–115. [Google Scholar]
- Luo, C.; Hajkova, P.; Ecker, J.R. Dynamic DNA methylation: In the right place at the right time. Science 2018, 361, 1336–1340. [Google Scholar] [CrossRef]
- Margot, J.B.; Ehrenhofer-Murray, A.E.; Leonhardt, H. Interactions within the mammalian DNA methyltransferase family. BMC Mol.Biol. 2003, 4, 7. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Bourc’his, D. The diverse roles of DNA methylation in mammalian development and disease. Nat. Rev. Mol. Cell Biol. 2019, 20, 590–607. [Google Scholar] [CrossRef]
- Santiago, M.; Antunes, C.; Guedes, M.; Sousa, N.; Marques, C.J. TET enzymes and DNA hydroxymethylation in neuronal development and function—How critical are they? Genomics 2014, 104, 334–340. [Google Scholar] [CrossRef]
- Stillman, B. Histone modifications: Insights into their influence on gene expression. Cell 2018, 175, 6–9. [Google Scholar] [CrossRef]
- Bhatti, G.K.; Khullar, N.; Sidhu, I.S.; Navik, U.S.; Reddy, A.P.; Reddy, P.H.; Bhatti, J.S. Emerging role of non-coding RNA in health and disease. Metabol. Brain Dis. 2021, 36, 1119–1134. [Google Scholar] [CrossRef]
- Seal, R.L.; Chen, L.-L.; Griffiths-Jones, S.; Lowe, T.M.; Mathews, M.B.; O’Reilly, D.; Pierce, A.J.; Stadler, P.F.; Ulitsky, I.; Wolin, S.L.; et al. A guide to naming human non-coding RNA genes. EMBO J. 2020, 39, e103777. [Google Scholar] [CrossRef]
- Kurumizaka, H.; Kujirai, T.; Takizawa, Y. Contributions of histone variants in nucleosome structure and function. J. Mol. Biol. 2021, 433, 166678. [Google Scholar] [CrossRef]
- Cavalli, G.; Heard, E. Advances in epigenetics link genetics to the environment and disease: Implications for genetic studies of complex disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef]
- Meaburn, E.L.; Schalkwyk, L.C.; Mill, J. Allele-specific methylation in the human genome. Epigenetics 2010, 5, 578–582. [Google Scholar] [CrossRef]
- Feil, R.; Fraga, M.F. Epigenetics and the environment: Emerging patterns and implications. Nat. Rev. Genet. 2012, 13, 97–109. [Google Scholar] [CrossRef]
- Feinberg, A.P.; Fallin, M.D. Epigenetics at the crossroads of genes and the environment. JAMA 2015, 314, 1129–1130. [Google Scholar] [CrossRef]
- Norouzitallab, P.; Baruah, K.; Vanrompay, D.; Bossier, P. Can epigenetics translate environmental cues into phenotypes? Sci. Total Environ. 2019, 647, 1281–1293. [Google Scholar] [CrossRef]
- Kaletsky, R.; Moore, R.S.; Vrla, G.D.; Parsons, L.R.; Gitai, Z.; Murphy, C.T. C. elegans interprets bacterial non-coding RNAs to learn pathogenic avoidance. Nature 2020, 586, 445–451. [Google Scholar] [CrossRef]
- Kendler, K.S. Incremental advances in psychiatric molecular genetics and nosology. World Psychiatry 2022, 21, 415–416. [Google Scholar] [CrossRef]
- Peedicayil, J.; Grayson, D.R. An epigenetic basis for an omnigenic model of psychiatric disorders. J. Theor. Biol. 2018, 443, 52–55. [Google Scholar] [CrossRef]
- Peedicayil, J.; Grayson, D.R. Some implications of an epigenetic-based omnigenic model of psychiatric disorders. J. Theor. Biol. 2018, 452, 81–84. [Google Scholar] [CrossRef]
- van Os, J.; Rutten, B.P.F.; Poulton, R. Gene-environment interactions in schizophrenia: Review of epidemiological findings and future directions. Schizophr. Bull. 2008, 34, 1066–1082. [Google Scholar] [CrossRef]
- Thomas, K.; Nilsson, E.; Festin, K.; Henriksson, P.; Lowén, M.; Löf, M.; Kristenson, M. Associations of psychosocial factors with multiple health behaviors: A population-based study of middle-aged men and women. Int. J. Environ. Res. Public Health 2020, 17, 1239. [Google Scholar] [CrossRef]
- Upton, J. Psychosocial factors. In Encyclopedia of Behavioral Medicine; Gelmann, M.D., Turner, J.R., Eds.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Amaresha, A.C.; Venkatasubramnian, G. Expressed emotion in schizophrenia: An overview. Indian J. Psychol. Med. 2012, 34, 12–20. [Google Scholar] [CrossRef]
- Butzlaff, R.L.; Hooley, J.M. Expressed emotion and psychiatric relapse. A meta-analysis. Arch. Gen. Psychiatry 1998, 55, 547–552. [Google Scholar] [CrossRef]
- Weaver, I.C.G.; Cervoni, N.; Champagne, F.A.; D’Alessio, A.C.; Sharma, S.; Seckl, J.R.; Dymov, S.; Szyf, M.; Meaney, M.J. Epigenetic programming by maternal behavior. Nat. Neurosci. 2004, 7, 847–854. [Google Scholar] [CrossRef]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef]
- Peedicayil, J. Psychosocial factors in the pathogenesis of psychiatric disorders. Br. J. Psychiatry 2004, 185, 520. [Google Scholar] [CrossRef]
- Peedicayil, J. Psychosocial factors may act via epigenetic mechanisms in the pathogenesis of mental disorders. Med. Hypotheses 2008, 70, 700–701. [Google Scholar] [CrossRef]
- Peedicayil, J. Epigenetics as a link between psychosocial factors and mental disorders. Indian J. Psychiatry 2015, 57, 218. [Google Scholar] [CrossRef]
- Thibaut, F. Epigenetics: The missing link between genes and psychiatric disorders? Dialogues Clin. Neurosci. 2019, 21, 337–338. [Google Scholar] [CrossRef]
- Dwyer, J.B.; Kaffman, A. Epigenetics in psychiatry: The promise for new biomarkers and treatments. In Kaplan & Sadock’s Comprehensive Textbook of Psychiatry; Sadock, B.J., Sadock, V.A., Ruiz, P., Eds.; Wolters Kluwer: Philadelphia, PA, USA, 2017. [Google Scholar]
- Peedicayil, J. The role of epigenetics in social psychiatry. Int. J. Soc. Psychiatry 2017, 63, 14–20. [Google Scholar] [CrossRef]
- Schiele, M.A.; Gottschalk, M.G.; Domschke, K. The applied implications of epigenetics in anxiety, affective and stress-related disorders—A review and synthesis on psychosocial stress, psychotherapy and prevention. Clin. Psychol. Rev. 2020, 77, 101830. [Google Scholar] [CrossRef]
- Hakamata, Y.; Suzuki, Y.; Kobashikawa, H.; Hori, H. Neurobiology of early life adversity: A systematic review of meta-analyses towards an integrative account of its neurobiological tragetories to mental disorders. Front. Neuroendocr. 2022, 65, 100994. [Google Scholar] [CrossRef]
- Jiménez, J.P.; Botto, A.; Herrera, L.; Leighton, C.; Rossi, J.L.; Quevedo, Y.; Silva, J.R.; Martínez, F.; Assar, R.; Salazar, L.A.; et al. Psychotherapy and genetic neuroscience: An emerging dialogue. Front. Genet. 2018, 9, 257. [Google Scholar] [CrossRef]
- Cannon, W.B. The interrelations of emotions as suggested by recent physiologic researches. Am. J. Psychol. 1914, 25, 256–282. [Google Scholar] [CrossRef]
- Selye, H. Stress and the general adaptation syndrome. Br. Med. J. 1950, 1, 1383–1392. [Google Scholar] [CrossRef]
- Jiang, S.; Postovit, L.; Cattaneo, A.; Binder, E.B.; Aitchison, K.J. Epigenetic modifications in stress response genes associated with childhood trauma. Front. Psychiatry 2019, 10, 808. [Google Scholar] [CrossRef]
- Criado-Marrero, M.; Rein, T.; Binder, E.B.; Porter, J.T.; Koren, J.; Blair, L.J. Hsp90 and FKBP51: Complex regulators of psychiatric diseases. Phil. Trans. R. Soc. B 2018, 373, 20160532. [Google Scholar] [CrossRef]
- Klengel, T.; Binder, E.B. Epigenetics of stress-related psychiatric disorders and gene x environment interactions. Neuron 2015, 86, 1343–1357. [Google Scholar] [CrossRef]
- Murgatroyd, C.; Patchev, A.V.; Wu, Y.; Micale, V.; Bockmühl, Y.; Fischer, D.; Holsboer, F.; Wotjak, C.T.; Almeida, O.F.X.; Spengler, D. Dynamic DNA methylation programs persistent adverse effects of early-life stress. Nat. Neurosci. 2009, 12, 1559–1566. [Google Scholar] [CrossRef]
- Murgatroyd, C.; Spengler, D. Polycomb binding precedes early-life stress responsive DNA methylation at the Avp enhancer. PLoS ONE 2014, 9, e90277. [Google Scholar] [CrossRef]
- Nuber, U.A.; Kriaucionis, S.; Roloff, T.C.; Guy, J.; Selfridge, J.; Steinhoff, C.; Schulz, R.; Lipkowitz, B.; Ropers, H.H.; Holmes, M.C.; et al. Up-regulation of glucocorticoid-regulated genes in a mouse model of Rett syndrome. Hum. Mol. Genet. 2005, 14, 2247–2256. [Google Scholar] [CrossRef] [PubMed]
- Ebert, D.H.; Gabel, H.W.; Robinson, N.D.; Kastan, N.R.; Hu, L.S.; Cohen, S.; Navarro, A.J.; Lyst, M.J.; Ekiert, R.; Bird, A.P.; et al. Activity-dependent phosphorylation of MeCP2 threonine 308 regulates interaction with NCoR. Nature 2013, 499, 341–345. [Google Scholar] [CrossRef]
- Elliott, E.; Ezra-Nevo, G.; Regev, L.; Neufeld-Cohen, A.; Chen, A. Resilience to social stress coincides with functional DNA methylation of the Crf gene in adult mice. Nat. Neurosci. 2010, 13, 1351–1353. [Google Scholar] [CrossRef]
- Laplant, Q.; Vialou, V.; Covington, H.E.; Dumitriu, D.; Feng, J.; Warren, B.L.; Maze, I.; Dietz, D.M.; Watts, E.L.; Iniguez, S.D.; et al. Dnmt3a regulates emotional behavior and spine plasticity in the nucleus accumbens. Nat. Neurosci. 2010, 13, 1137–1143. [Google Scholar] [CrossRef]
- Levine, A.; Worrell, T.R.; Zimnisky, R.; Schmauss, C. Early life stress triggers sustained changes in histone deacetylase expression and histone H4 modifications that alter responsiveness to adolescent antidepressant treatment. Neurobiol. Dis. 2012, 45, 488–498. [Google Scholar] [CrossRef]
- Brandeis, M.; Frank, D.; Keshet, L.; Siegfried, Z.; Mendelsohn, M.; Nemes, A.; Temper, V.; Razin, A.; Cedar, H. Sp1 elements protect a CpG island from de novo methylation. Nature 1994, 371, 435–438. [Google Scholar] [CrossRef]
- Ochi, S.; Dwivedi, Y. Dissecting early life stress-induced adolescent depression through epigenomic approach. Mol. Psychiatry 2023, 28, 141–153. [Google Scholar] [CrossRef]
- Zucchi, F.C.R.; Yao, Y.; Ward, I.D.; Ilnytskyy, Y.; Olson, D.M.; Benzies, K.; Kovalchuk, I.; Kovalchuk, O.; Metz, G.A.S. Maternal stress induces epigenetic signatures of psychiatric and neurological diseases in the offspring. PLoS ONE 2013, 8, e56967. [Google Scholar] [CrossRef]
- Guidotti, A.; Dong, E.; Tueting, P.; Grayson, D.R. Modeling the molecular epigenetic profile of psychosis in prenatally stressed mice. Prog. Mol. Biol. Transl. Sci. 2014, 128, 89–101. [Google Scholar]
- Grayson, D.R.; Guidotti, A. DNA methylation in animal models of psychosis. Prog. Mol. Biol. Transl. Sci. 2018, 157, 105–132. [Google Scholar]
- Matrisciano, F.; Tueting, P.; Dalal, I.; Kadriu, B.; Grayson, D.R.; Davis, J.M.; Nicoletti, F.; Guidotti, A. Epigenetic modifications of GABAergic interneurons are associated with the schizophrenia-like phenotype induced by prenatal stress in mice. Neuropharmacology 2013, 68, 184–194. [Google Scholar] [CrossRef]
- Nazzari, S.; Grumi, S.; Mambretti, F.; Villa, M.; Giorda, R.; Provenzi, L. MOM-COPE Study Group. Maternal and infant NR3C1 and SLC6A4 epigenetic signatures of the COVID-19 pandemic lockdown: When timing matters. Transl. Psychiatry 2022, 12, 386. [Google Scholar] [CrossRef]
- Jablensky, A.V.; Morgan, V.; Zubrick, S.R.; Bower, C.; Yellachich, L.-A. Pregnancy, delivery, and neonatal complications in a population cohort of women with schizophrenia and affective disorders. Am. J. Psychiatry 2005, 162, 79–91. [Google Scholar] [CrossRef]
- Palma-Gudiel, H.; Eixarch, E.; Crispi, F.; Morán, S.; Zannas, A.S.; Fananás, L. Prenatal adverse environment is associated with epigenetic age deceleration at birth and hypomethylation at the hypoxia-responsive EP300 gene. Clin. Epigenet. 2019, 11, 73. [Google Scholar] [CrossRef]
- Patel, V.; Kleinman, A. Poverty and common mental disorders in developing countries. Bull. World Health Organ. 2003, 81, 609–615. [Google Scholar]
- Murali, V.; Oyebode, F. Poverty, social inequality, and mental health. Adv. Psychiatr. Treat. 2004, 10, 216–224. [Google Scholar] [CrossRef]
- Hoare, J.; Stein, D.J.; Heany, S.J.; Fouche, J.-P.; Phillips, N.; Er, S.; Myer, L.; Zar, H.J.; Horvath, S.; Levine, A.J. Accelerated epigenetic aging in adolescents from low-income households is associated with altered development of brain structures. Metab. Brain Dis. 2020, 35, 1287–1298. [Google Scholar] [CrossRef]
- Cantor-Graae, E.; Pedersen, C.B. Full spectrum of psychiatric disorders related to foreign migration: A Danish population-based cohort study. JAMA Psychiatry 2013, 70, 427–435. [Google Scholar] [CrossRef]
- Stilo, S.A.; Murray, R.M. Non-genetic factors in schizophrenia. Curr. Psychiatry Rep. 2019, 21, 100. [Google Scholar] [CrossRef]
- Clausing, E.S.; Non, A.L. Epigenetics as a mechanism of developmental embodiment of stress. Front. Psychiatry 2021, 12, 696827. [Google Scholar] [CrossRef]
- Cronjé, H.T.; Elliot, H.R.; Nienaber-Rousseau, C.; Pieters, M. Leveraging the urban-rural divide for epigenetic research. Epigenomics 2020, 12, 1071–1081. [Google Scholar] [CrossRef]
- Cannon, M.; Clarke, M.C. Risk for schizophrenia-Broadening the concepts. Schizophr. Res. 2005, 79, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Whittier, A.B.; Gelaye, B.; Deyessa, N.; Bahretibeb, Y.; Kelkile, T.S.; Berhane, Y.; Wiliams, M.A. Major depressive disorder and suicidal behavior among urban dwelling Ethiopian outpatients at a general hospital. J. Affect. Disord. 2016, 197, 58–65. [Google Scholar]
- Beydoun, M.A.; Hossain, S.; Chitrala, K.N.; Tajuddin, S.M.; Bedoun, S.A.; Evans, M.K.; Zonderman, A.B. Association between epigenetic age acceleration and depressive symptoms in a prospective cohort study of urban-dwelling adults. J. Affect. Disord. 2019, 257, 64–73. [Google Scholar] [CrossRef]
- Gatta, E.; Saudagar, V.; Guidotti, A. Alcohol use disorder and associated alterations in brain epigenetic marks. In Epigenetics in Psychiatry; Peedicayil, J., Grayson, D.R., Avramopoulos, D., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 599–617. [Google Scholar]
- Milian, A.A.; Bendersky, C.J.; Walker, D. Epigenetic mechanisms of drug addiction vulnerability. In Epigenetics in Psychiatry; Peedicayil, J., Grayson, D.R., Avramopoulos, D., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 575–598. [Google Scholar]
- Peedicayil, J.; Santhosh, S. Epigenetic aspects of the microbiota and psychiatric disorders. In Epigenetics in Psychiatry; Peedicayil, J., Grayson, D.R., Avramopoulos, D., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 783–791. [Google Scholar]
- Li, J.; Ma, Y.; Bao, Z.; Gui, X.; Li, A.N.; Yang, Z.; Li, M.D. Clostridiales are predominant microbes that mediate psychiatric disorders. J. Psychiatr. Res. 2020, 130, 48–56. [Google Scholar] [CrossRef]
- Weber-Stadlbauer, U. Epigenetic and transgenerational mechanisms in infection-mediated neurodevelopmental disorders. Transl. Psychiatry 2017, 7, e1113. [Google Scholar] [CrossRef]
- Boks, M.P.; de Jong, N.M.; Kas, M.J.H.; Vinkers, C.H.; Fernandez, C.; Kahn, R.S.; Mill, J.; Ophoff, R.A. Current status and future prospects for epigenetic psychopharmacology. Epigenetics 2012, 7, 20–28. [Google Scholar] [CrossRef]
- Peedicayil, J.; Santhosh, S. Epigenetic effects of currently used psychotropic drugs. In Epigenetics in Psychiatry; Peedicayil, J., Grayson, D.R., Avramopoulos, D., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 621–637. [Google Scholar]
- Logan, R.W.; Ozburn, A.R.; Arey, R.N.; Ketchesin, K.D.; Winquist, A.; Crain, A.; Tobe, B.T.D.; Becker-Krail, D.; Jarpe, M.B.; Xue, X.; et al. Valproate reverses mania-like behaviors in mice by preferentially targeting of HDAC2. Mol. Psychiatry 2021, 26, 4066–4084. [Google Scholar] [CrossRef]
- Wu, M.-S.; Li, X.-J.; Liu, C.-Y.; Xu, Q.; Huang, J.-Q.; Gu, S.; Chen, J.-X. Effects of histone modification in major depressive disorder. Curr. Neuropharmacol. 2022, 20, 1261–1277. [Google Scholar]
- Meng, Y.; Du, J.; Liu, N.; Qiang, Y.; Xiao, L.; Lan, X.; Ma, L.; Yang, J.; Yu, J.; Lu, G. Epigenetic modulation: Research progress on histone acetylation levels in major depressive disorders. J. Drug Target. 2023, 31, 142–151. [Google Scholar] [CrossRef]
- Cuomo, A.; Crescenzi, B.B.; Bolognesi, S.; Goracci, A.; Koukouna, D.; Rossi, R.; Fagiolini, A. S-adenosylmethionine (SAME) in major depressive disorder (MDD): A clinician-oriented systematic review. Ann. Gen. Psychiatry 2020, 19, 50. [Google Scholar] [CrossRef]
- Lam, N.S.K.; Long, X.X.; Li, X.; Saad, M.; Lim, F.; Doery, J.C.; Griffin, R.C.; Galletly, C. The potential use of folate and its derivatives in treating psychiatric disorders: A systematic review. Biomed. Pharmacother. 2022, 146, 112541. [Google Scholar] [CrossRef]
- Roffman, J.L.; Petruzzi, L.J.; Tanner, A.S.; Brown, H.E.; Eryilmaz, H.; Ho, N.F.; Giegold, M.; Silverstein, N.J.; Bottiglieri, T.; Manoach, D.S.; et al. Biochemical, physiological and clinical effects of L-methylfolate in schizophrenia: A randomized controlled trial. Mol. Psychiatry 2018, 23, 316–322. [Google Scholar] [CrossRef]
- Nierenberg, A.A.; Montana, R.; Kinrys, G.; Deckersbach, T.; Dufour, S.; Baek, J.H. L-methylfolate for bipolar I depressive episodes: An open trial proof-of-concept registry. J. Affect. Disord. 2017, 207, 429–433. [Google Scholar] [CrossRef]
- Zeigler, C.; Richter, J.; Mahr, M.; Gajewska, A.; Schiele, M.A.; Gehrmann, A.; Schmidt, B.; Lesch, K.-P.; Lang, T.; Helbig-Lang, S.; et al. MAOA gene hypomethylation in panic disorder-Reversibility of an epigenetic risk pattern by psychotherapy. Transl. Psychiatry 2016, 6, e773. [Google Scholar] [CrossRef]
- Zeigler, C.; Grundner-Culemann, F.; Schiele, M.A.; Schlosser, P.; Kollert, L.; Mahr, M.; Gajewska, A.; Lesch, K.-P.; Deckert, J.; Köttgen, A.; et al. The DNA methylome in panic disorder: A case-control and longitudinal psychotherapy-epigenetic study. Transl. Psychiatry 2019, 9, 314. [Google Scholar] [CrossRef]
- Syed, S.A.; Zannas, A.S. Epigenetics in psychotherapy. In Epigenetics in Psychiatry; Peedicayil, J., Grayson, D.R., Avramopoulos, D., Eds.; Elsevier: San Diego, CA, USA, 2021; pp. 701–709. [Google Scholar]
- Schiele, M.A.; Thiel, C.; Deckert, J.; Zaudig, M.; Berberich, G.; Domschke, K. Monoamine oxidase A hypomethylation in obsessive compulsive disorder: Reversibility by successful psychotherapy? Int. J. Neuropsychopharmacol. 2020, 23, 319–323. [Google Scholar] [CrossRef]
- Jiang, J.; Wang, J.; Li, C. Potential mechanisms underlying the therapeutic effects of electroconvulsive therapy. Neurosci. Bull. 2017, 33, 339–347. [Google Scholar] [CrossRef]
- Sirignano, L.; Frank, J.; Kranaster, L.; Witt, S.H.; Streit, F.; Zillich, L.; Sartorius, A.; Rietschel, M.; Foo, J.C. Methylome-wide change associated with response to electroconvulsive therapy in depressed patients. Transl. Psychiatry 2021, 11, 347. [Google Scholar] [CrossRef]
- Patki, G.; Solanki, N.; Atrooz, F.; Ansari, A.; Allam, F.; Jannise, B.; Maturi, J.; Salim, S. Novel mechanistic insights into treadmill exercise based rescue of social defeat-induced anxiety-like behavior and memory impairment in rats. Physiol. Behav. 2014, 130, 135–144. [Google Scholar] [CrossRef]
- Chekroud, S.R.; Gueorguieva, R.; Zheutlin, A.B.; Paulus, M.; Krumholz, H.M.; Krystal, J.H.; Chekroud, A.M. Association between physical exercise and mental health in 1.2 million individuals in the USA between 2011 and 2015: A cross-sectional study. Lancet Psychiatry 2018, 5, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Grasdalsmoen, M.; Eriksen, H.R.; Lønning, K.J.; Sivertsen, B. Physical exercise, mental health problems, and suicide attempts in university students. BMC Psychiatry 2020, 20, 175. [Google Scholar] [CrossRef]
- Ntanasis-Stathopoulos, J.; Tzanninis, J.G.; Philippou, A.; Koutsilieris, M. Epigenetic regulation on gene expression induced by physical exercise. J. Musculoskelet. Neuronal Interact. 2013, 13, 133–146. [Google Scholar] [PubMed]
- Fernandez, J.; Arida, R.M.; Gomez-Pinilla, F. Physical exercise as an epigenetic modulator of brain plasticity and cognition. Neurosci. Biobehav. Rev. 2017, 80, 443–456. [Google Scholar] [CrossRef]


{kind=link}
| Social determinants of mental health including early life adversity and early life stress |
| Maternal prenatal stress |
| Poverty |
| Migration |
| Urban dwelling |
| Pregnancy and birth complications |
| Alcohol use |
| Substance use other than alcohol |
| Microbiota |
| Prenatal and postnatal infections |
| Psychotropic drugs |
| Psychotherapy |
| Electroconvulsive therapy |
| Physical exercise |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peedicayil, J. Genome–Environment Interactions and Psychiatric Disorders. Biomedicines 2023, 11, 1209. https://doi.org/10.3390/biomedicines11041209
Peedicayil J. Genome–Environment Interactions and Psychiatric Disorders. Biomedicines. 2023; 11(4):1209. https://doi.org/10.3390/biomedicines11041209
Chicago/Turabian StylePeedicayil, Jacob. 2023. "Genome–Environment Interactions and Psychiatric Disorders" Biomedicines 11, no. 4: 1209. https://doi.org/10.3390/biomedicines11041209
APA StylePeedicayil, J. (2023). Genome–Environment Interactions and Psychiatric Disorders. Biomedicines, 11(4), 1209. https://doi.org/10.3390/biomedicines11041209