Enhanced Proliferative and Osteogenic Potential of Periodontal Ligament Stromal Cells


 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
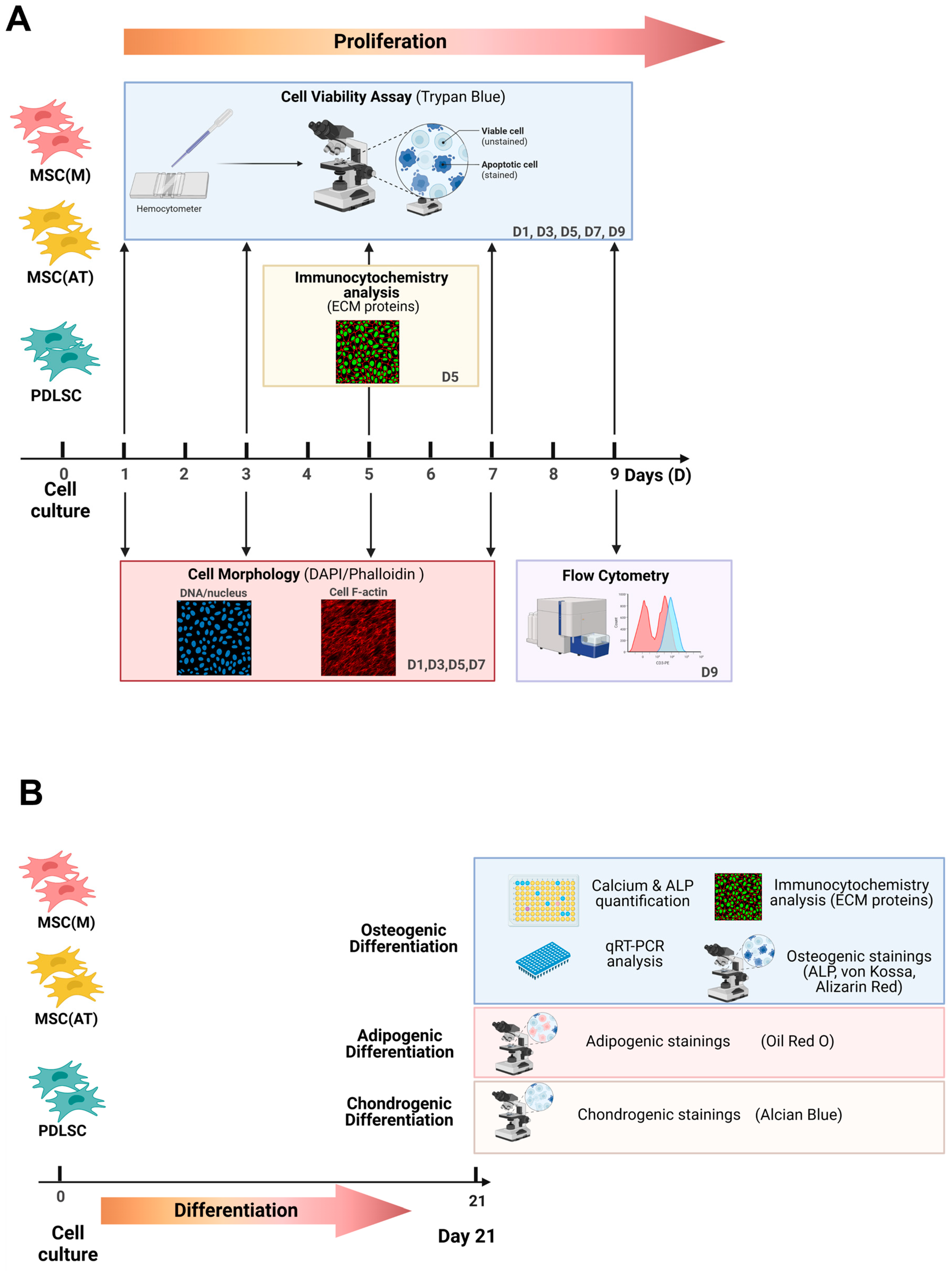
2. Materials and Methods
2.1. Isolation and Culture of Human PDLSC
2.2. Culture of MSC(M) and MSC(AT)
2.3. Multilineage Differentiation and Stainings
2.3.1. Osteogenic Differentiation
Alkaline Phosphatase (ALP) Activity Assay
Calcium Quantification Assay
2.3.2. Adipogenic Differentiation
2.3.3. Chondrogenic Differentiation
2.4. Flow Cytometry Analysis
2.5. Immunocytochemistry Analysis
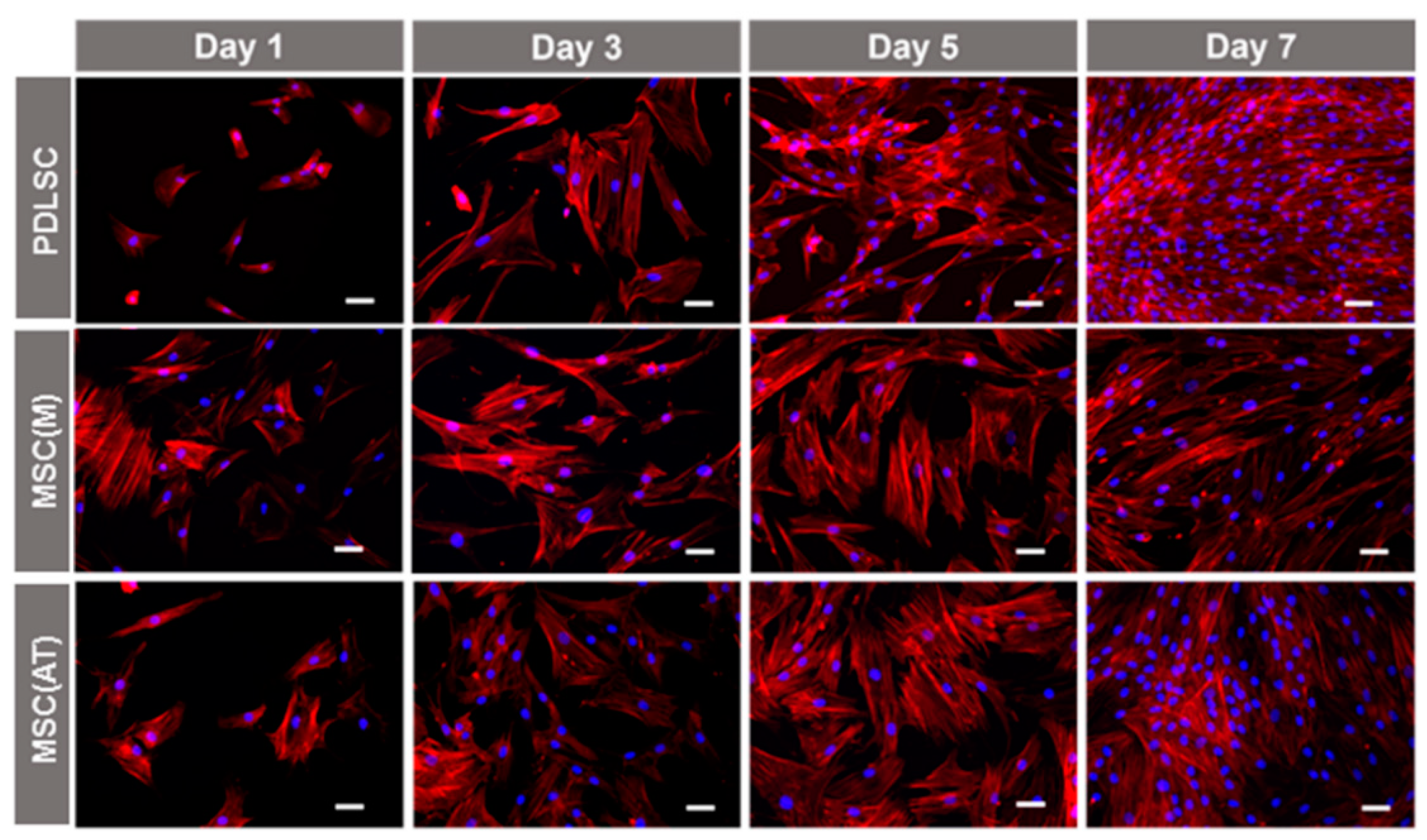
2.6. Cell Morphology Assays
2.7. Proliferation Assays
2.8. Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR) Analysis
2.9. Statistical Analysis
3. Results
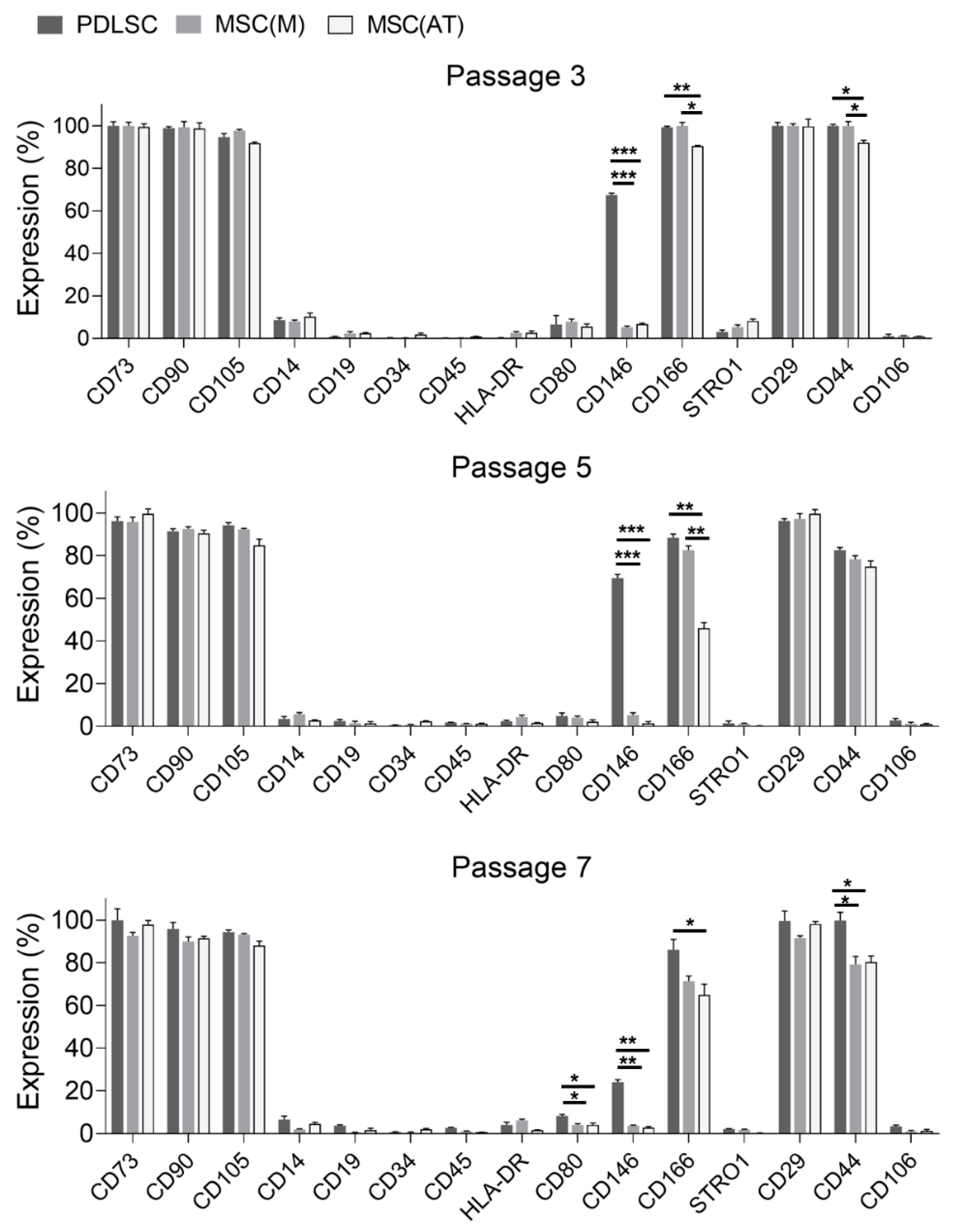
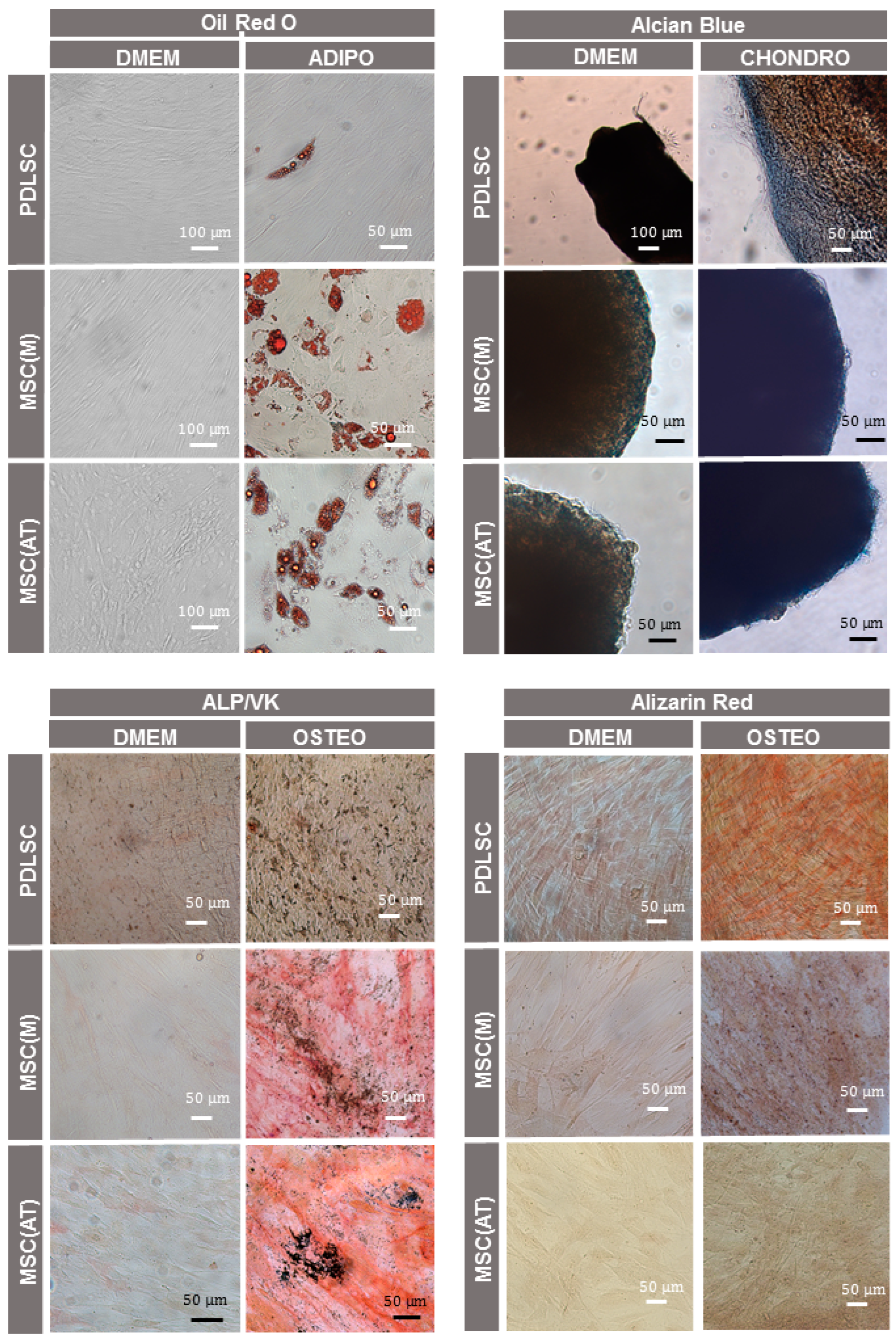
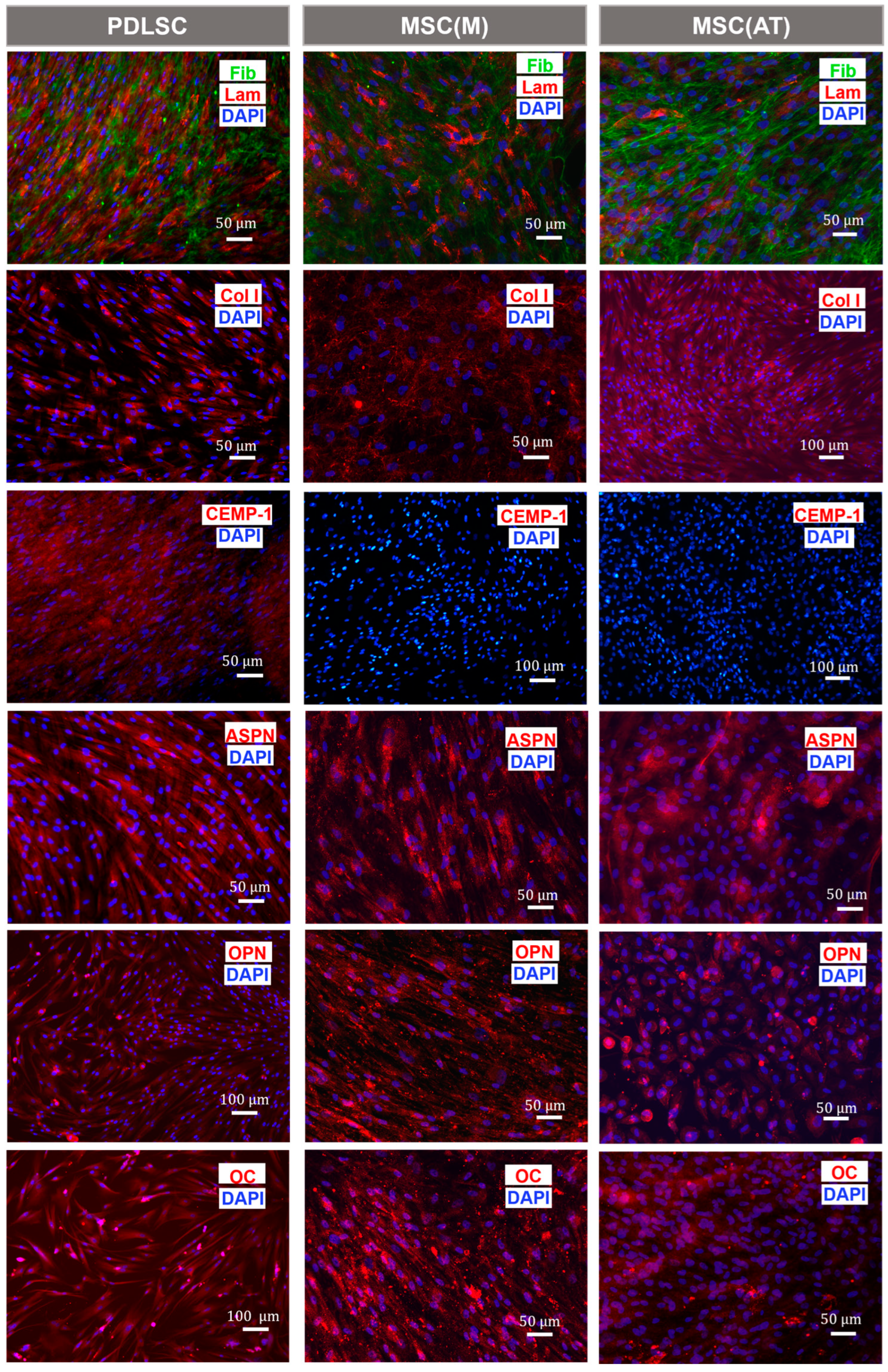
3.1. Characterization of MSC Derived from Different Adult Tissue Sources: Periodontal Ligament, Bone Marrow and Adipose Tissue
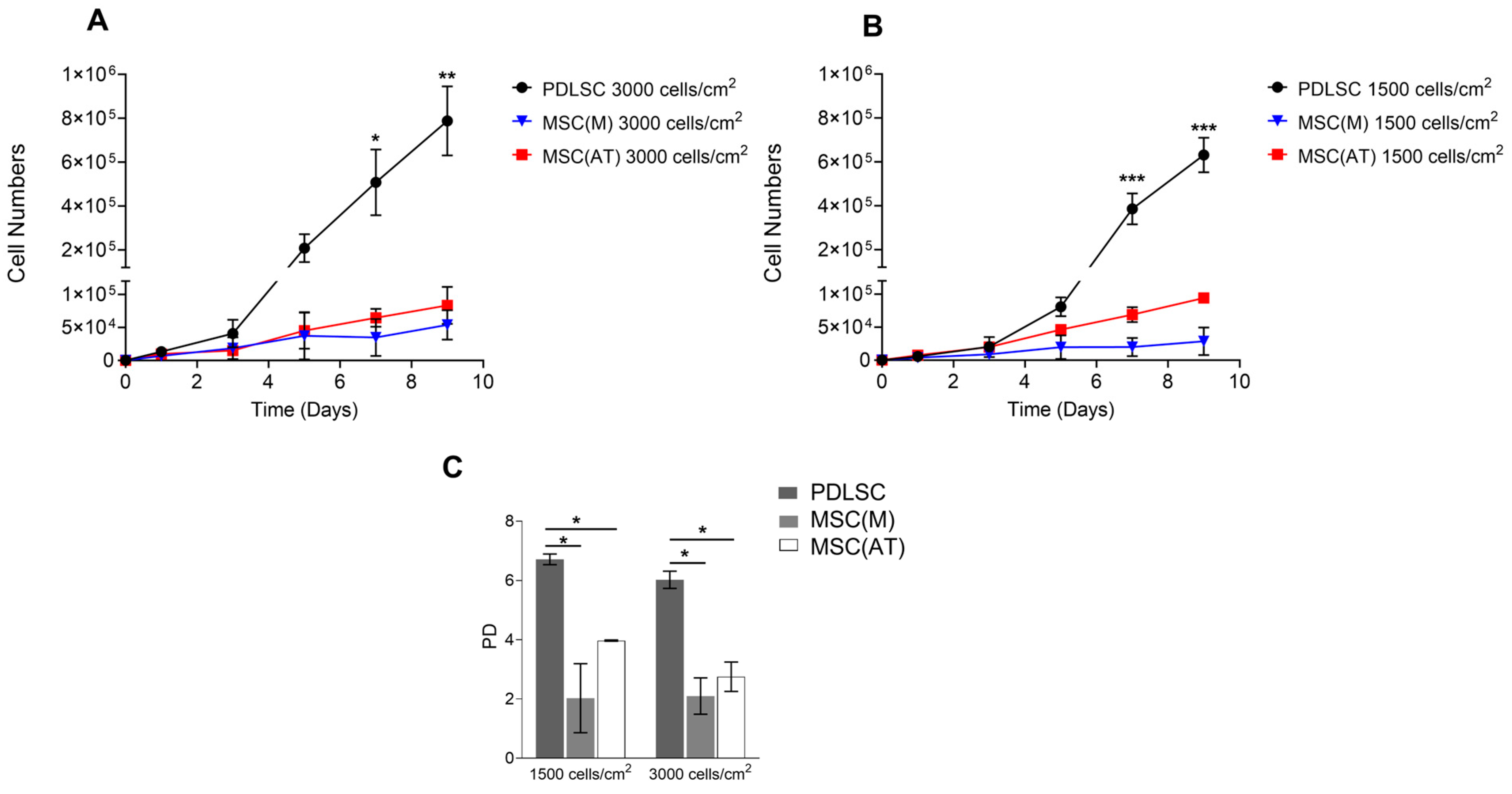
3.2. Proliferative Potential of MSC Derived from Periodontal Ligament, Bone Marrow and Adipose Tissue
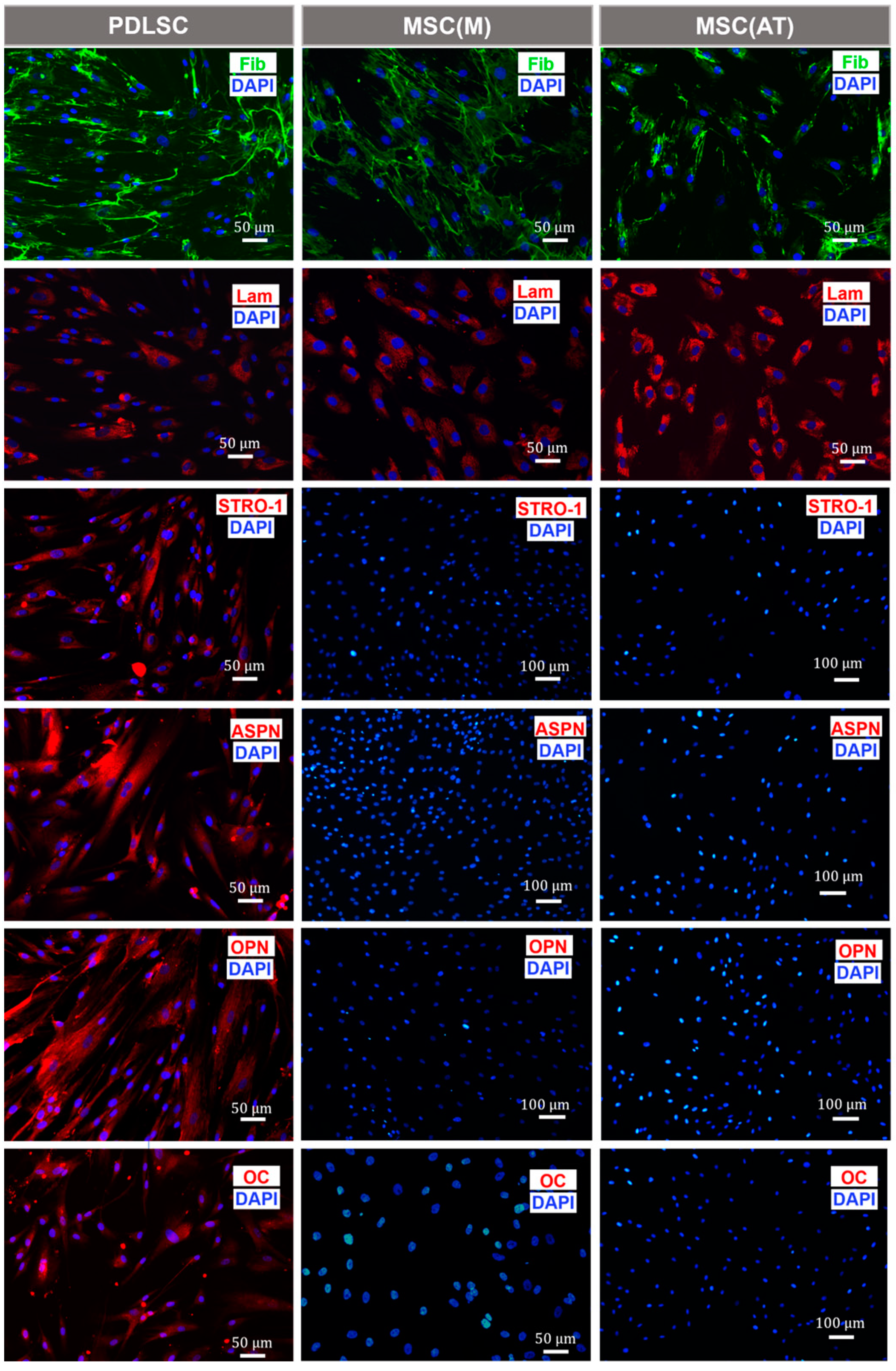
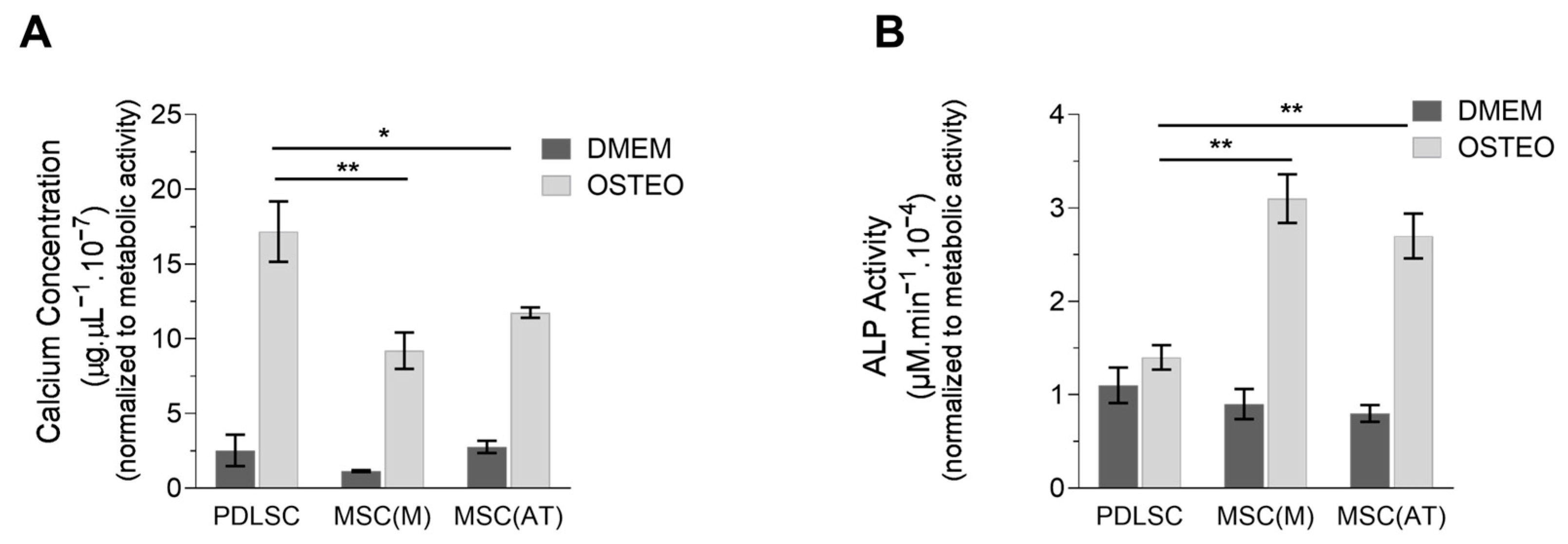
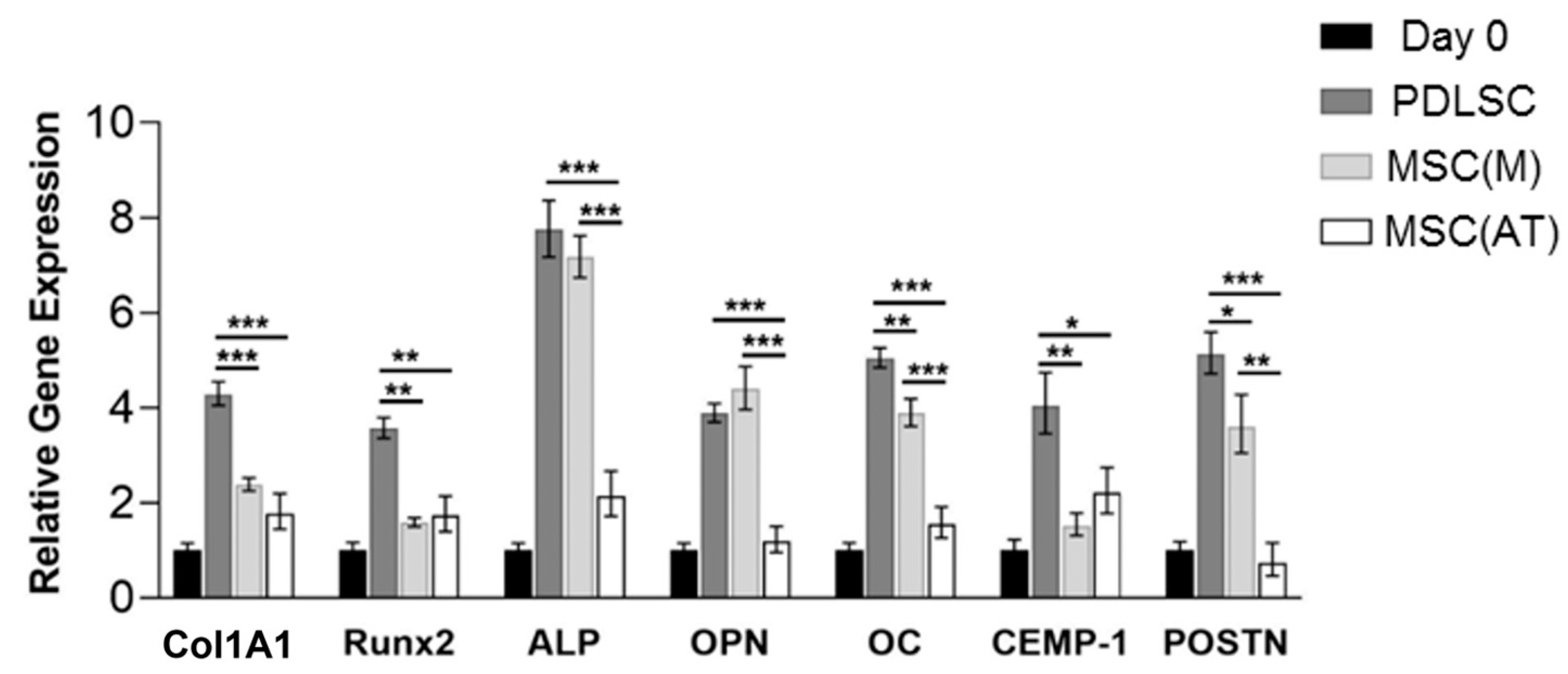
3.3. Osteogenic Potential of MSC Derived Periodontal Ligament, Bone Marrow and Adipose Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, M.X.; Zhong, Y.J.; Dong, Q.Q.; Wong, H.M.; Wen, Y.F. Global, Regional, and National Burden of Severe Periodontitis, 1990–2019: An Analysis of the Global Burden of Disease Study 2019. J. Clin. Periodontol. 2021, 48, 1165–1188. [Google Scholar] [CrossRef]
- Machado, V.; Botelho, J.; Escalda, C.; Hussain, S.B.; Luthra, S.; Mascarenhas, P.; Orlandi, M.; Mendes, J.J.; D’Aiuto, F. Serum C-Reactive Protein and Periodontitis: A Systematic Review and Meta-Analysis. Front. Immunol. 2021, 12, 706432. [Google Scholar] [CrossRef]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal Diseases. Nat. Rev. Dis. Prim. 2017, 3, 17038. [Google Scholar] [CrossRef]
- Ferreira, M.C.; Dias-Pereira, A.C.; Branco-de-Almeida, L.S.; Martins, C.C.; Paiva, S.M. Impact of Periodontal Disease on Quality of Life: A Systematic Review. J. Periodontal Res. 2017, 52, 651–665. [Google Scholar] [CrossRef]
- Botelho, J.; Machado, V.; Leira, Y.; Proença, L.; Chambrone, L.; Mendes, J.J. Economic Burden of Periodontitis in the United States and Europe—An Updated Estimation. J. Periodontol. 2021, 93, 373–379. [Google Scholar] [CrossRef]
- Preshaw, P.M.; Seymour, R.A.; Heasman, P.A. Current Concepts in Periodontal Pathogenesis. Dent. Update 2004, 31, 570–578. [Google Scholar] [CrossRef]
- Botelho, J.; Mascarenhas, P.; Viana, J.; Proença, L.; Orlandi, M.; Leira, Y.; Chambrone, L.; Mendes, J.J.; Machado, V. An Umbrella Review of the Evidence Linking Oral Health and Systemic Health: From the Prevalence to Clinical and Circulating Markers. medRxiv 2022. [Google Scholar] [CrossRef]
- Larsson, L.; Decker, A.M.; Nibali, L.; Pilipchuk, S.P.; Berglundh, T.; Giannobile, W.V. Regenerative Medicine for Periodontal and Peri-Implant Diseases. J. Dent. Res. 2016, 95, 255–266. [Google Scholar] [CrossRef]
- Du, J.; Shan, Z.; Ma, P.; Wang, S.; Fan, Z. Allogeneic Bone Marrow Mesenchymal Stem Cell Transplantation for Periodontal Regeneration. J. Dent. Res. 2014, 93, 183–188. [Google Scholar] [CrossRef]
- Citterio, F.; Gualini, G.; Fierravanti, L.; Aimetti, M. Stem Cells and Periodontal Regeneration: Present and Future. Plast. Aesthetic Res. 2020, 7, 41. [Google Scholar] [CrossRef]
- Carvalho, M.S.; Alves, L.; Bogalho, I.; Cabral, J.M.S.; da Silva, C.L. Impact of Donor Age on the Osteogenic Supportive Capacity of Mesenchymal Stromal Cell-Derived Extracellular Matrix. Front. Cell Dev. Biol. 2021, 9, 747521. [Google Scholar] [CrossRef]
- Viswanathan, S.; Ciccocioppo, R.; Galipeau, J.; Krampera, M.; Le Blanc, K.; Martin, I.; Moniz, K.; Nolta, J.; Phinney, D.G.; Shi, Y.; et al. Consensus International Council for Commonality in Blood Banking Automation–International Society for Cell & Gene Therapy Statement on Standard Nomenclature Abbreviations for the Tissue of Origin of Mesenchymal Stromal Cells. Cytotherapy 2021, 23, 1060–1063. [Google Scholar] [CrossRef]
- Seo, B.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.; Shi, S. Investigation of Multipotent Postnatal Stem Cells from Human Periodontal Ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef]
- Bartold, P.M.; Shi, S.; Gronthos, S. Stem Cells and Periodontal Regeneration. Periodontol. 2000 2006, 40, 164–172. [Google Scholar] [CrossRef]
- Qu, G.; Li, Y.; Chen, L.; Chen, Q.; Zou, D.; Yang, C.; Zhou, Q. Comparison of Osteogenic Differentiation Potential of Human Dental-Derived Stem Cells Isolated from Dental Pulp, Periodontal Ligament, Dental Follicle, and Alveolar Bone. Stem Cells Int. 2021, 2021, 6631905. [Google Scholar] [CrossRef]
- Simmons, P.J.; Torok-Storb, B. Identification of Stromal Cell Precursors in Human Bone Marrow by a Novel Monoclonal Antibody, STRO-1. Blood 1991, 78, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Nagatomo, K.; Komaki, M.; Sekiya, I.; Sakaguchi, Y.; Noguchi, K.; Oda, S.; Muneta, T.; Ishikawa, I. Stem Cell Properties of Human Periodontal Ligament Cells. J. Periodontal Res. 2006, 41, 303–310. [Google Scholar] [CrossRef]
- Itaya, T.; Kagami, H.; Okada, K.; Yamawaki, A.; Narita, Y.; Inoue, M.; Sumita, Y.; Ueda, M. Characteristic Changes of Periodontal Ligament-Derived Cells during Passage. J. Periodontal Res. 2009, 44, 425–433. [Google Scholar] [CrossRef]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-Derived Stem Cells for Regenerative Medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Dos Santos, F.; Andrade, P.Z.; Boura, J.S.; Abecasis, M.M.; da Silva, C.L.; Cabral, J.M.S. Ex Vivo Expansion of Human Mesenchymal Stem Cells: A More Effective Cell Proliferation Kinetics and Metabolism under Hypoxia. J. Cell. Physiol. 2009, 223, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Moreno Sancho, F.; Leira, Y.; Orlandi, M.; Buti, J.; Giannobile, W.V.; D’Aiuto, F. Cell-Based Therapies for Alveolar Bone and Periodontal Regeneration: Concise Review. Stem Cells Transl. Med. 2019, 8, 1286–1295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, T.; Nakagawa, T. Mesenchymal Stem Cell-Based Tissue Regeneration Therapies for Periodontitis. Regen. Ther. 2020, 14, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Winning, L.; El Karim, I.A.; Lundy, F.T. A Comparative Analysis of the Osteogenic Potential of Dental Mesenchymal Stem Cells. Stem Cells Dev. 2019, 28, 1050–1058. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Menicanin, D.; Shi, S.; Bartold, P.M.; Gronthos, S. Immunomodulatory Properties of Human Periodontal Ligament Stem Cells. J. Cell. Physiol. 2009, 219, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Banavar, S.R.; Rawal, S.Y.; Paterson, I.C.; Singh, G.; Davamani, F.; Khoo, S.P.; Tan, E.L. Establishing a Technique for Isolation and Characterization of Human Periodontal Ligament Derived Mesenchymal Stem Cells. Saudi Dent. J. 2020, 33, 693–701. [Google Scholar] [CrossRef]
- Xu, L.; Liu, Y.; Sun, Y.; Wang, B.; Xiong, Y.; Lin, W.; Wei, Q.; Wang, H.; He, W.; Wang, B.; et al. Tissue Source Determines the Differentiation Potentials of Mesenchymal Stem Cells: A Comparative Study of Human Mesenchymal Stem Cells from Bone Marrow and Adipose Tissue. Stem Cell Res. Ther. 2017, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-Derived and Bone Marrow Mesenchymal Stem Cells: A Donor-Matched Comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef]
- Kobolak, J.; Dinnyes, A.; Memic, A.; Khademhosseini, A.; Mobasheri, A. Mesenchymal Stem Cells: Identification, Phenotypic Characterization, Biological Properties and Potential for Regenerative Medicine through Biomaterial Micro-Engineering of Their Niche. Methods 2016, 99, 62–68. [Google Scholar] [CrossRef]
- Tamaki, Y.; Nakahara, T.; Ishikawa, H.; Sato, S. In Vitro Analysis of Mesenchymal Stem Cells Derived from Human Teeth and Bone Marrow. Odontology 2013, 101, 121–132. [Google Scholar] [CrossRef]
- Petrenko, Y.; Vackova, I.; Kekulova, K.; Chudickova, M.; Koci, Z.; Turnovcova, K.; Kupcova Skalnikova, H.; Vodicka, P.; Kubinova, S. A Comparative Analysis of Multipotent Mesenchymal Stromal Cells Derived from Different Sources, with a Focus on Neuroregenerative Potential. Sci. Rep. 2020, 10, 4290. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.-H.K.; Ogando, C.R.; Wang See, C.; Chang, T.-Y.; Barabino, G.A. Changes in Phenotype and Differentiation Potential of Human Mesenchymal Stem Cells Aging in Vitro. Stem Cell Res. Ther. 2018, 9, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, G.; Liu, G.; Banie, L.; Wang, G.; Ning, H.; Lue, T.F.; Lin, C.-S. Tissue Distribution of Mesenchymal Stem Cell Marker Stro-1. Stem Cells Dev. 2011, 20, 1747–1752. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.-J.; Tuan, R.S.; Cheung, K.M.C.; Leung, V.Y.L. Concise Review: The Surface Markers and Identity of Human Mesenchymal Stem Cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef] [PubMed]
- Guezguez, B.; Vigneron, P.; Lamerant, N.; Kieda, C.; Jaffredo, T.; Dunon, D. Dual Role of Melanoma Cell Adhesion Molecule (MCAM)/CD146 in Lymphocyte Endothelium Interaction: MCAM/CD146 Promotes Rolling via Microvilli Induction in Lymphocyte and Is an Endothelial Adhesion Receptor. J. Immunol. 2007, 179, 6673–6685. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Huang, Z.; Wu, D.; Kou, X.; Mao, X.; Shi, S. CD146 Controls the Quality of Clinical Grade Mesenchymal Stem Cells from Human Dental Pulp. Stem Cell Res. Ther. 2021, 12, 488. [Google Scholar] [CrossRef]
- Yamada, S.; Kitamura, M.; Murakami, S. PLAP-1: A Novel Molecule Regulating Homeostasis of Periodontal Tissues. Jpn. Dent. Sci. Rev. 2008, 44, 137–144. [Google Scholar] [CrossRef] [Green Version]
- Stein, G.S.; Lian, J.B.; Owen, T.A. Relationship of Cell Growth to the Regulation of Tissue-Specific Gene Expression during Osteoblast Differentiation. FASEB J. 1990, 4, 3111–3123. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Bai, J.; Ji, X.; Li, R.; Xuan, Y.; Wang, Y. Comprehensive Characterization of Four Different Populations of Human Mesenchymal Stem Cells as Regards Their Immune Properties, Proliferation and Differentiation. Int. J. Mol. Med. 2014, 34, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Raheja, L.F.; Genetos, D.C.; Yellowley, C.E. Hypoxic Osteocytes Recruit Human MSCs through an OPN/CD44-Mediated Pathway. Biochem. Biophys. Res. Commun. 2008, 366, 1061–1066. [Google Scholar] [CrossRef]
- Zou, C.; Luo, Q.; Qin, J.; Shi, Y.; Yang, L.; Ju, B.; Song, G. Osteopontin Promotes Mesenchymal Stem Cell Migration and Lessens Cell Stiffness via Integrin Β1, FAK, and ERK Pathways. Cell Biochem. Biophys. 2013, 65, 455–462. [Google Scholar] [CrossRef]
- Rogall, R.; Rabenstein, M.; Vay, S.; Bach, A.; Pikhovych, A.; Baermann, J.; Hoehn, M.; Couillard-Despres, S.; Fink, G.R.; Schroeter, M.; et al. Bioluminescence Imaging Visualizes Osteopontin-Induced Neurogenesis and Neuroblast Migration in the Mouse Brain after Stroke. Stem Cell Res. Ther. 2018, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Poggio, P.; Grau, J.B.; Field, B.C.; Sainger, R.; Seefried, W.F.; Rizzolio, F.; Ferrari, G. Osteopontin Controls Endothelial Cell Migration in Vitro and in Excised Human Valvular Tissue from Patients with Calcific Aortic Stenosis and Controls. J. Cell. Physiol. 2011, 226, 2139–2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, S.K.; Johnston, H.M.; Whitty, G.A.; Williams, B.; Webb, R.J.; Denhardt, D.T.; Bertoncello, I.; Bendall, L.J.; Simmons, P.J.; Haylock, D.N. Osteopontin, a Key Component of the Hematopoietic Stem Cell Niche and Regulator of Primitive Hematopoietic Progenitor Cells. Blood 2005, 106, 1232–1239. [Google Scholar] [CrossRef]
- Nojima, N.; Kobayashi, M.; Shionome, M.; Takahashi, N.; Suda, T.; Hasegawa, K. Fibroblastic Cells Derived from Bovine Periodontal Ligaments Have the Phenotypes of Osteoblasts. J. Periodontal Res. 1990, 25, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Basdra, E.K.; Komposch, G. Osteoblast-like Properties of Human Periodontal Ligament Cells: An in Vitro Analysis. Eur. J. Orthod. 1997, 19, 615–621. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Pei, X.; Zhao, Y.; Tulu, U.S.; Liu, B.; Helms, J.A. A Wnt-Responsive PDL Population Effectuates Extraction Socket Healing. J. Dent. Res. 2018, 97, 803–809. [Google Scholar] [CrossRef]
- Yu, Z.; Philippe, G. Differential Properties of Human ALP+ Periodontal Ligament Stem Cells vs Their ALP− Counterparts. J. Stem Cell Res. Ther. 2015, 5, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.T.; Penny, C.B.; Paiker, J.E.; Psaras, G.; Ikram, F.; Crowther, N.J. The Relationship between Alkaline Phosphatase Activity and Intracellular Lipid Accumulation in Murine 3T3-L1 Cells and Human Preadipocytes. Anal. Biochem. 2006, 354, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ferris, W.; Penny, C.; Van der Merwe, M.-T.; Jacobson, B.; Paiker, J.; Crowther, N. Lipid Accumulation and Alkaline Phosphatase Activity in Human Preadipocytes Isolated from Different Body Fat Depots. J. Endocrinol. Metab. Diabetes S. Afr. 2013, 18, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Birmingham, E.; Niebur, G.; McHugh, P.; Shaw, G.; Barry, F.; McNamara, L. Osteogenic Differentiation of Mesenchymal Stem Cells Is Regulated by Osteocyte and Osteoblast Cells in a Simplified Bone Niche. Eur. Cells Mater. 2012, 23, 13–27. [Google Scholar] [CrossRef]
- Hayrapetyan, A.; Jansen, J.A.; van den Beucken, J.J.J.P. Signaling Pathways Involved in Osteogenesis and Their Application for Bone Regenerative Medicine. Tissue Eng. Part B Rev. 2015, 21, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.S.; Poundarik, A.A.; Cabral, J.M.S.; da Silva, C.L.; Vashishth, D. Biomimetic Matrices for Rapidly Forming Mineralized Bone Tissue Based on Stem Cell-Mediated Osteogenesis. Sci. Rep. 2018, 8, 14388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, M.S.; Cabral, J.M.; da Silva, C.L.; Vashishth, D. Synergistic Effect of Extracellularly Supplemented Osteopontin and Osteocalcin on Stem Cell Proliferation, Osteogenic Differentiation, and Angiogenic Properties. J. Cell. Biochem. 2019, 120, 6555–6569. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, F.; Zhang, N.; Geng, X.; Meng, C.; Wang, X.; Yang, Y. Osteogenic Capacity and Cytotherapeutic Potential of Periodontal Ligament Cells for Periodontal Regeneration in Vitro and in Vivo. PeerJ 2019, 7, e6589. [Google Scholar] [CrossRef] [Green Version]
- Kadokura, H.; Yamazaki, T.; Masuda, Y.; Kato, Y.; Hasegawa, A.; Sakagami, H.; Yokose, S. Establishment of a Primary Culture System of Human Periodontal Ligament Cells That Differentiate into Cementum Protein 1-Expressing Cementoblast-like Cells. In Vivo 2019, 33, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Arzate, H.; Olson, S.W.; Page, R.C.; Gown, A.M.; Narayanan, A.S. Production of a Monoclonal Antibody to an Attachment Protein Derived from Human Cementum. FASEB J. 1992, 6, 2990–2995. [Google Scholar] [CrossRef]
- McCulloch, C.A.G.; Melcher, A.H. Cell Density and Cell Generation in the Periodontal Ligament of Mice. Am. J. Anat. 1983, 167, 43–58. [Google Scholar] [CrossRef]
- Shinagawa-Ohama, R.; Mochizuki, M.; Tamaki, Y.; Suda, N.; Nakahara, T. Heterogeneous Human Periodontal Ligament-Committed Progenitor and Stem Cell Populations Exhibit a Unique Cementogenic Property Under In Vitro and In Vivo Conditions. Stem Cells Dev. 2017, 26, 632–645. [Google Scholar] [CrossRef]
- Kii, I.; Amizuka, N.; Minqi, L.; Kitajima, S.; Saga, Y.; Kudo, A. Periostin Is an Extracellular Matrix Protein Required for Eruption of Incisors in Mice. Biochem. Biophys. Res. Commun. 2006, 342, 766–772. [Google Scholar] [CrossRef]
- Rios, H.; Koushik, S.V.; Wang, H.; Wang, J.; Zhou, H.-M.; Lindsley, A.; Rogers, R.; Chen, Z.; Maeda, M.; Kruzynska-Frejtag, A.; et al. Periostin Null Mice Exhibit Dwarfism, Incisor Enamel Defects, and an Early-Onset Periodontal Disease-Like Phenotype. Mol. Cell. Biol. 2005, 25, 11131–11144. [Google Scholar] [CrossRef] [Green Version]
- Iwata, T.; Yamato, M.; Washio, K.; Yoshida, T.; Tsumanuma, Y.; Yamada, A.; Onizuka, S.; Izumi, Y.; Ando, T.; Okano, T.; et al. Periodontal Regeneration with Autologous Periodontal Ligament-Derived Cell Sheets—A Safety and Efficacy Study in Ten Patients. Regen. Ther. 2018, 9, 38–44. [Google Scholar] [CrossRef]
- Zhao, Y.; Gong, Y.; Liu, X.; He, J.; Zheng, B.; Liu, Y. The Experimental Study of Periodontal Ligament Stem Cells Derived Exosomes with Hydrogel Accelerating Bone Regeneration on Alveolar Bone Defect. Pharmaceutics 2022, 14, 2189. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, P.; Wang, Y.; Li, J.; Qiao, D.; Chen, R.; Yang, W.; Yan, F. Gold Nanoparticles Promote the Bone Regeneration of Periodontal Ligament Stem Cell Sheets Through Activation of Autophagy. Int. J. Nanomed. 2021, 16, 61–73. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | GGTCACCAGGCTTTTA | CCTGGAAGATATGGGA |
| Col1A1 | CATCTCCCCTTCGTTTTTGA | CCAAATCCGATGTTTCTGCT |
| Runx2 | AGATGATGACACTGCCACCTCTG | GGGATGAAATGCTTGGGAACT |
| ALP | ACCATTCCCACGTCTTCACATTT | AGACATTCTCTCGTTCACCGCC |
| OPN | TGTGAGGTGATGTCCTCGTCTGTAG | ACACATATGATGGCCGAGGTGA |
| OC | TGCCTCAGAAGAGCTGAAAAC | CACAGACTCCCTGCTTTTGCT |
| CEMP-1 | ACATCAAGCACTGACAGCCA | GTTGATCTCCGCCCATAAGC |
| POSTN | ACATCAAGCACTGACAGCCA | GCCTCCAATATGTCCGATGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alves, L.; Machado, V.; Botelho, J.; Mendes, J.J.; Cabral, J.M.S.; da Silva, C.L.; Carvalho, M.S. Enhanced Proliferative and Osteogenic Potential of Periodontal Ligament Stromal Cells. Biomedicines 2023, 11, 1352. https://doi.org/10.3390/biomedicines11051352
Alves L, Machado V, Botelho J, Mendes JJ, Cabral JMS, da Silva CL, Carvalho MS. Enhanced Proliferative and Osteogenic Potential of Periodontal Ligament Stromal Cells. Biomedicines. 2023; 11(5):1352. https://doi.org/10.3390/biomedicines11051352
Chicago/Turabian StyleAlves, Laura, Vanessa Machado, João Botelho, José João Mendes, Joaquim M. S. Cabral, Cláudia L. da Silva, and Marta S. Carvalho. 2023. "Enhanced Proliferative and Osteogenic Potential of Periodontal Ligament Stromal Cells" Biomedicines 11, no. 5: 1352. https://doi.org/10.3390/biomedicines11051352
APA StyleAlves, L., Machado, V., Botelho, J., Mendes, J. J., Cabral, J. M. S., da Silva, C. L., & Carvalho, M. S. (2023). Enhanced Proliferative and Osteogenic Potential of Periodontal Ligament Stromal Cells. Biomedicines, 11(5), 1352. https://doi.org/10.3390/biomedicines11051352