
Biosynthesized Selenium Nanoparticles Using Epigallocatechin Gallate Protect against Pentylenetetrazole-Induced Acute Epileptic Seizures in Mice via Antioxidative, Anti-Inflammatory, and Anti-Apoptotic Activities

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs
Preparation and Characterization of EGCG-SeNPs
2.2. Experimental Design
2.3. Seizures Induction by PTZ
2.4. Tissue Sampling
2.5. Estimation of Oxidant/Antioxidant Condition
2.6. Estimation of Inflammatory Biomarkers
2.7. Estimation of Apoptosis Biomarkers
2.8. Gene Expression Analysis
2.9. Estimation of Neurotransmitters Levels
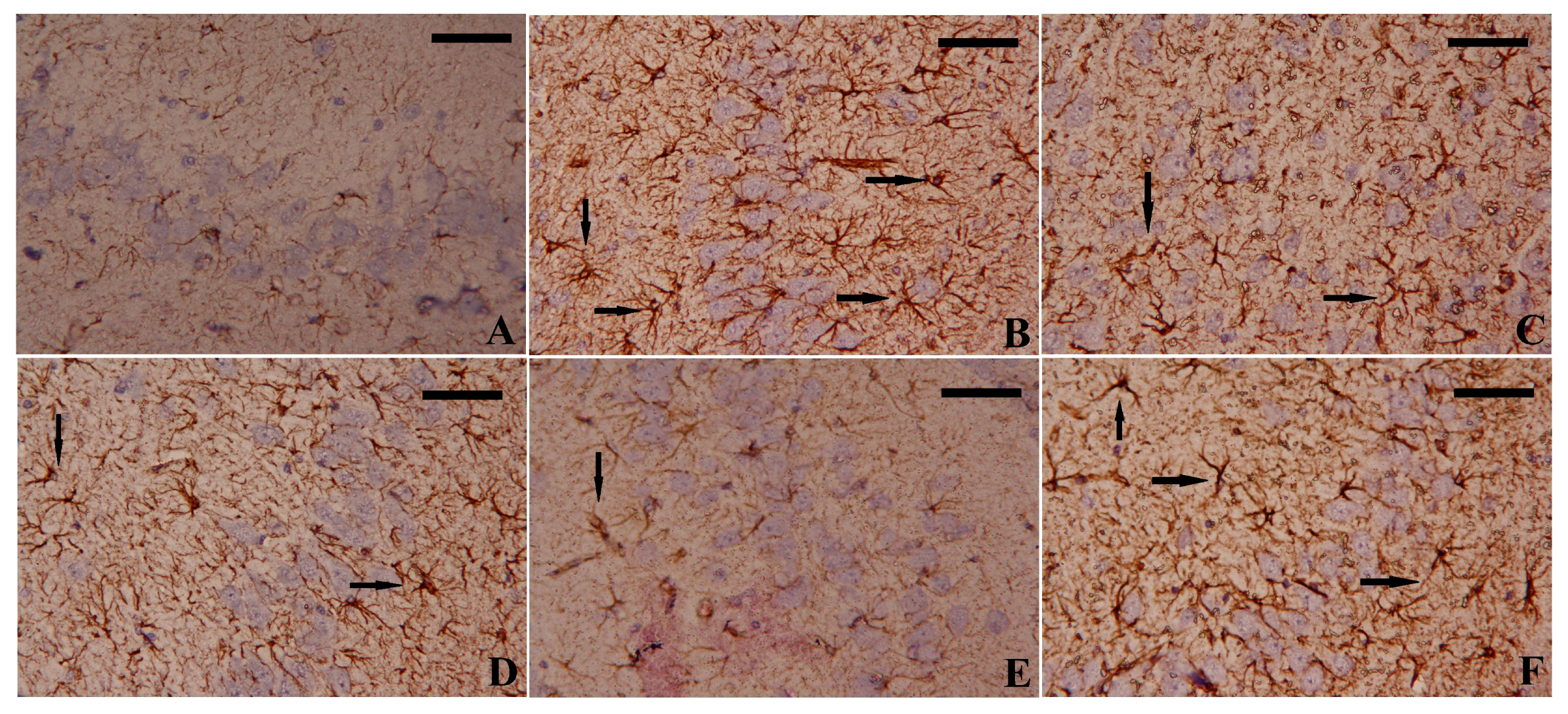
2.10. Estimation of Brain Astrocytes and Microglia Biomarkers
2.11. Immunohistochemistry Analysis
2.12. Histopathological Examination
2.13. Statistical Analysis
3. Results
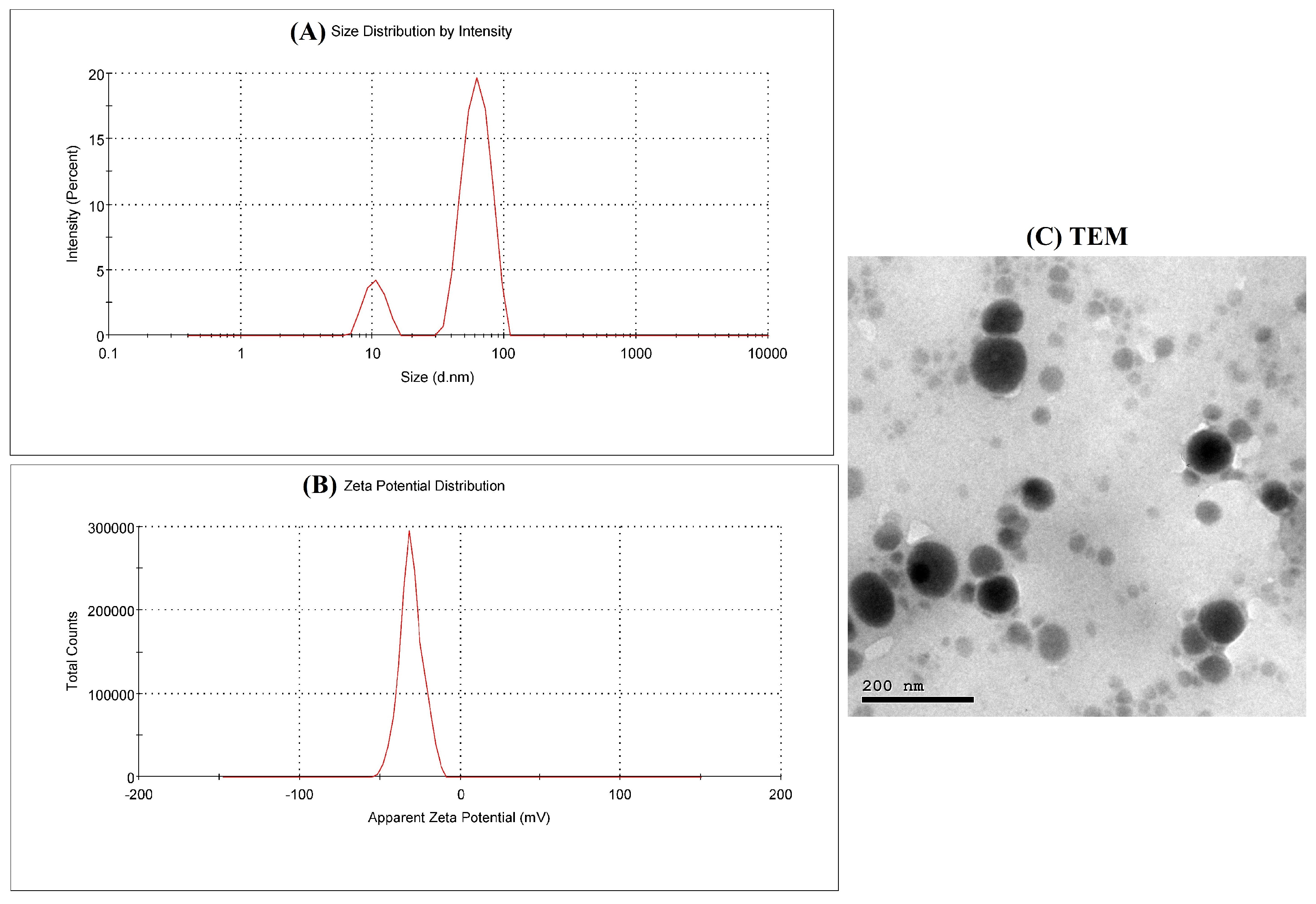
3.1. EGCG-SeNPs Characterization
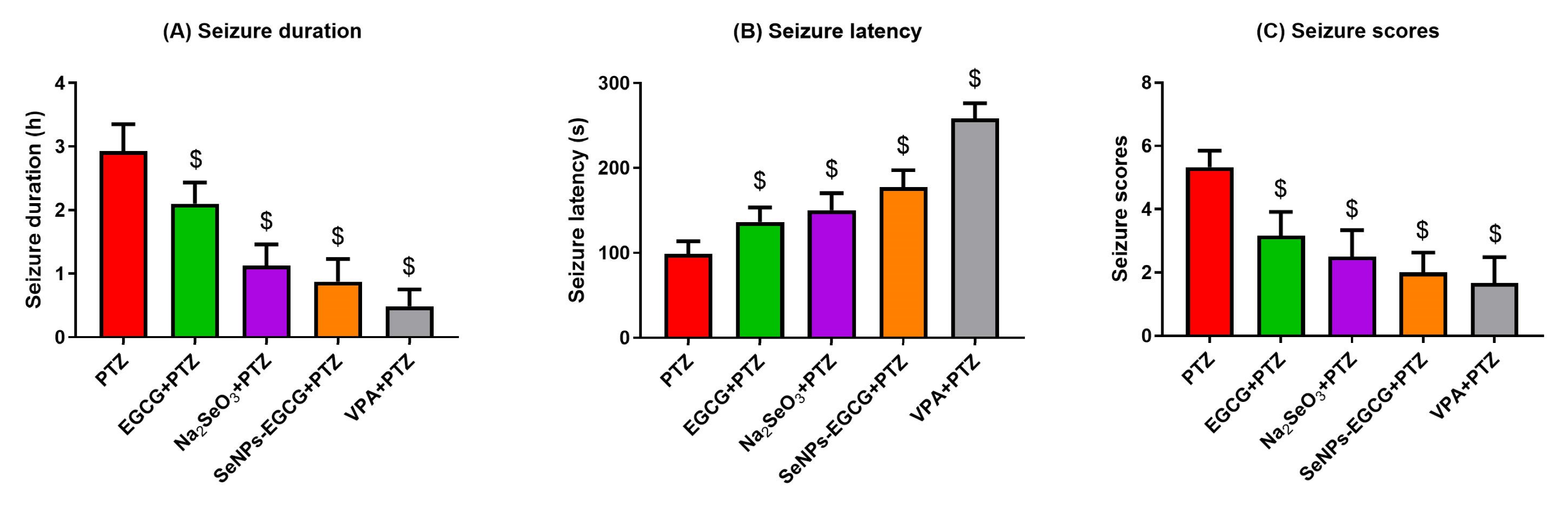
3.2. Epileptic Seizures Analysis
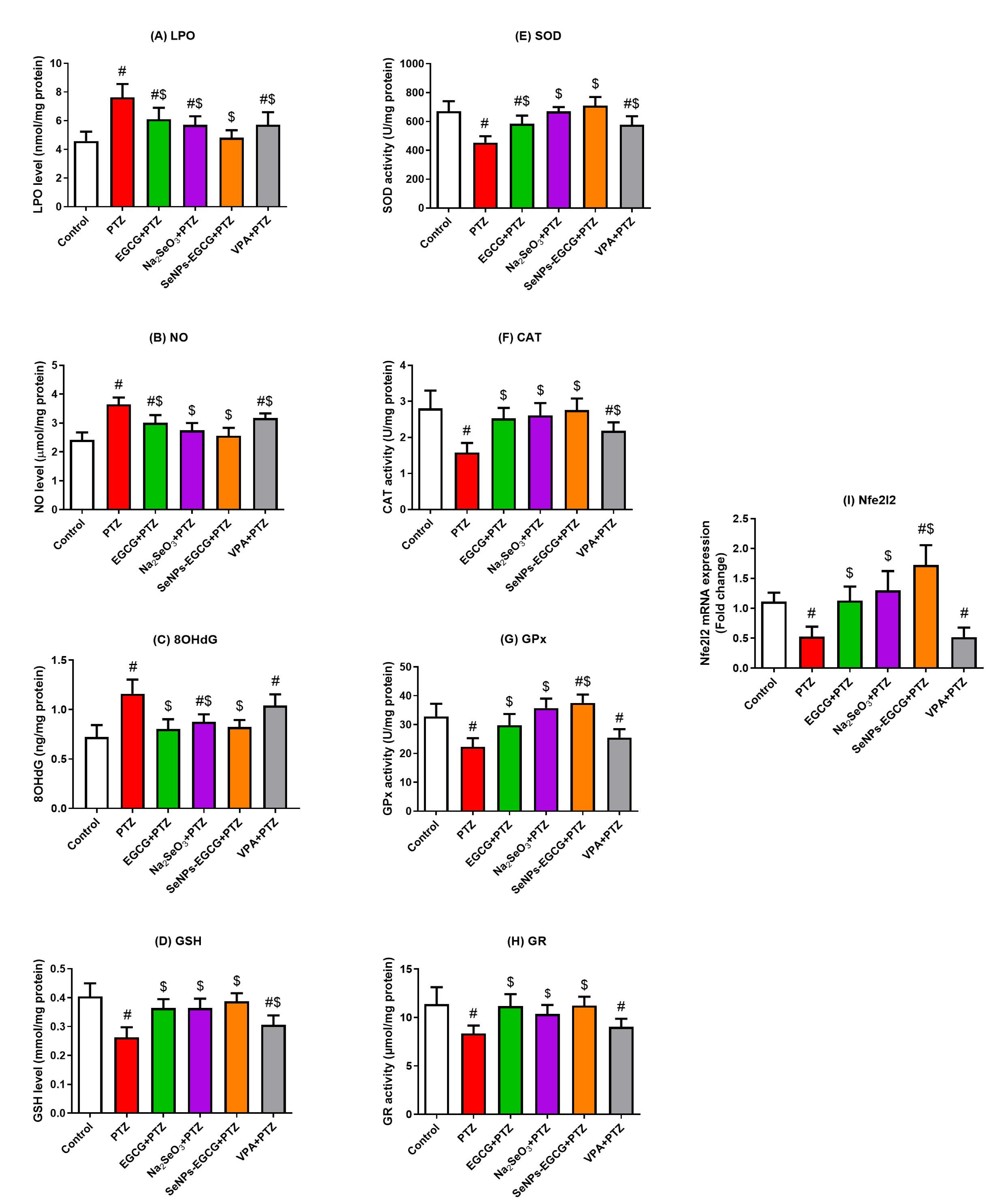
3.3. Antioxidant Activity of EGCG-SeNPs against PTZ-Induced Oxidative Damage
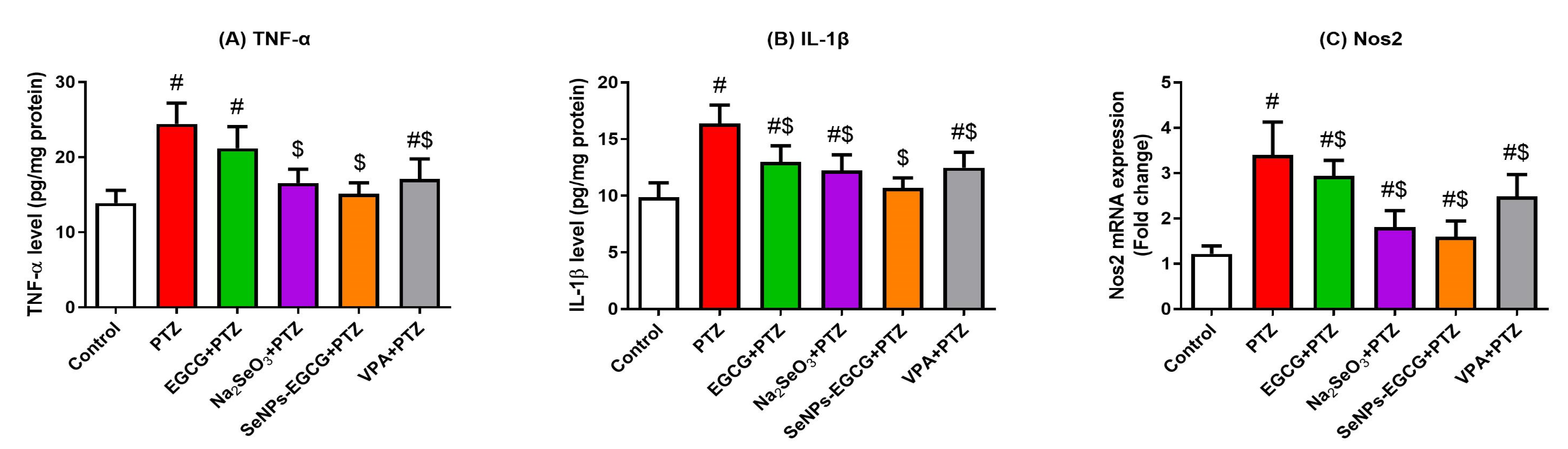
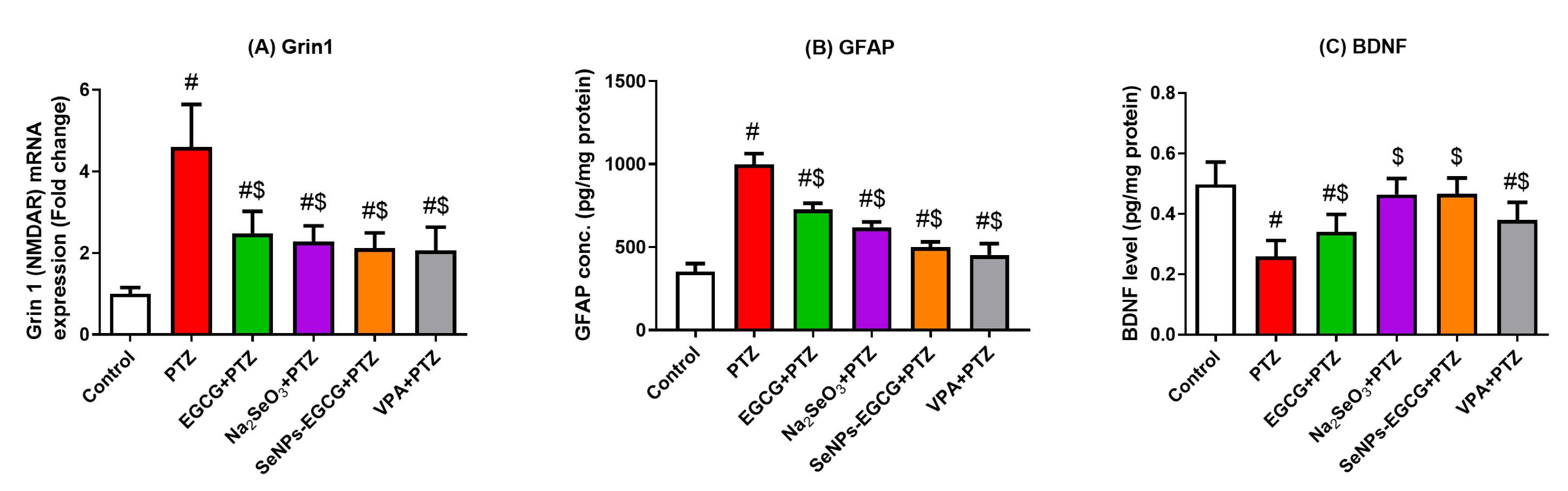
3.4. Anti-Inflammation Effect of EGCG-SeNPs against the PTZ-Induced Neuro-Inflammation
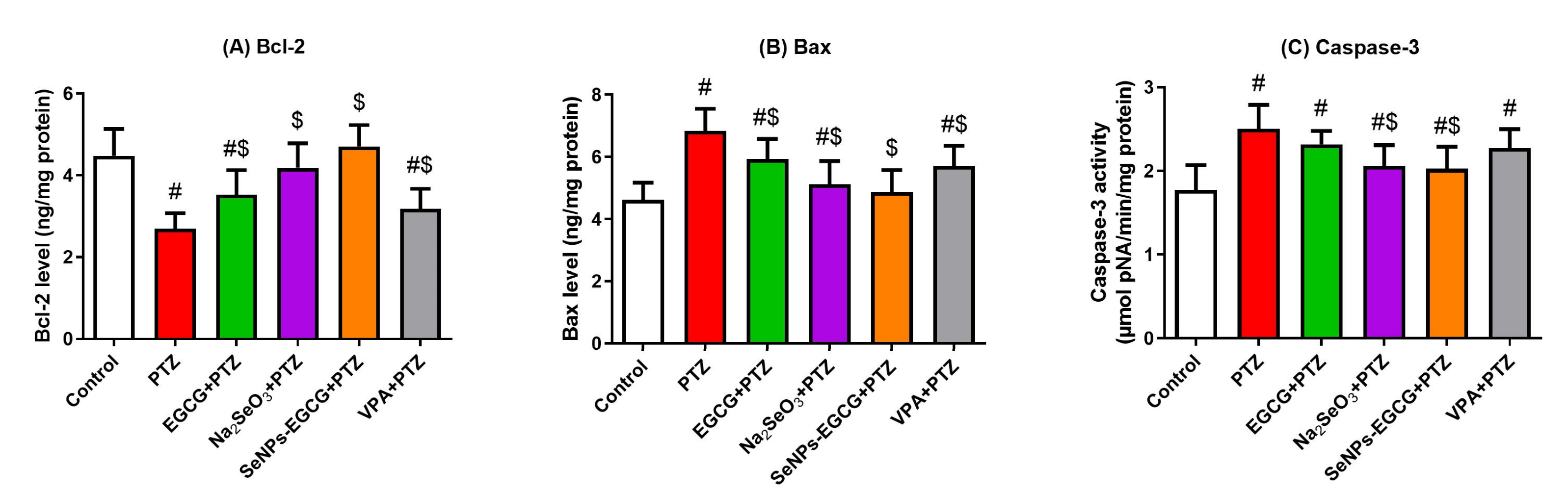
3.5. Anti-Apoptotic Activity of EGCG-SeNPs against the PTZ-Provoked Neuronal Apoptosis
3.6. Effect of EGCG-SeNPs Treatment on the Neurotransmission
3.7. The Effect of EGCG-SeNPs on the Neurological Function during the Epileptic Seizures
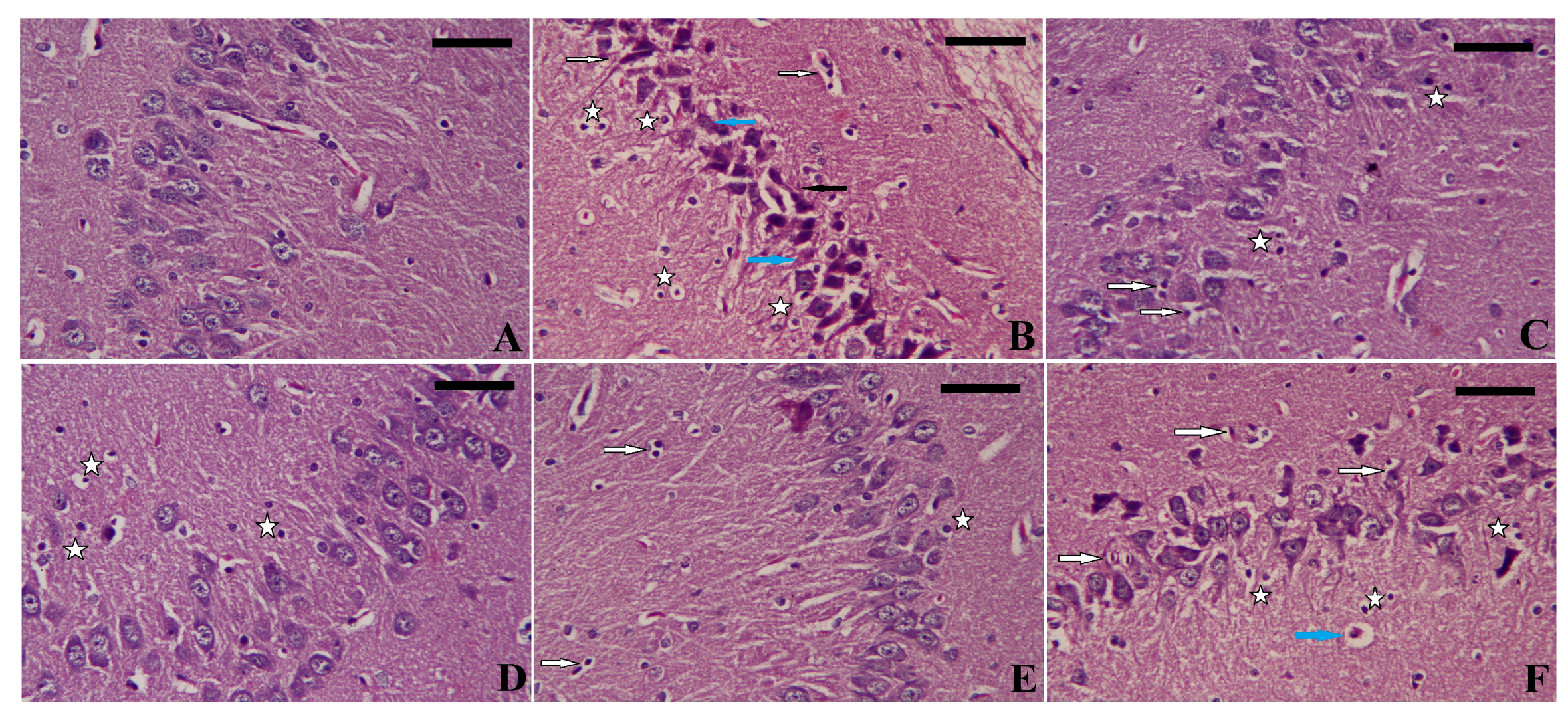
3.8. Effect of EGCG-SeNPs on the PTZ-Induced Histopathological Alterations during the Epileptic Seizures
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fisher, R.S.; Acevedo, C.; Arzimanoglou, A.; Bogacz, A.; Cross, J.H.; Elger, C.E.; Engel, J., Jr.; Forsgren, L.; French, J.A.; Glynn, M.; et al. ILAE Official Report: A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef]
- Riva, A.; Golda, A.; Balagura, G.; Amadori, E.; Vari, M.S.; Piccolo, G.; Iacomino, M.; Lattanzi, S.; Salpietro, V.; Minetti, C.; et al. New Trends and Most Promising Therapeutic Strategies for Epilepsy Treatment. Front. Neurol. 2021, 12, 753753. [Google Scholar] [CrossRef] [PubMed]
- Avanzini, G.; Franceschetti, S.; Mantegazza, M. Epileptogenic channelopathies: Experimental models of human pathologies. Epilepsia 2007, 48 (Suppl. S2), 51–64. [Google Scholar] [CrossRef]
- Mendez-Armenta, M.; Nava-Ruiz, C.; Juarez-Rebollar, D.; Rodriguez-Martinez, E.; Gomez, P.Y. Oxidative stress associated with neuronal apoptosis in experimental models of epilepsy. Oxid. Med. Cell Longev 2014, 2014, 293689. [Google Scholar] [CrossRef] [PubMed]
- Cavazos, J.E.; Jones, S.M.; Cross, D.J. Sprouting and synaptic reorganization in the subiculum and CA1 region of the hippocampus in acute and chronic models of partial-onset epilepsy. Neuroscience 2004, 126, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Mangan, P.S.; Scott, C.A.; Williamson, J.M.; Bertram, E.H. Aberrant neuronal physiology in the basal nucleus of the amygdala in a model of chronic limbic epilepsy. Neuroscience 2000, 101, 377–391. [Google Scholar] [CrossRef]
- Mao, X.-Y.; Zhou, H.-H.; Jin, W.-L. Redox-related neuronal death and crosstalk as drug targets: Focus on epilepsy. Front. Neurosci. 2019, 13, 512. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- da Fonsêca, D.V.; da Silva Maia Bezerra Filho, C.; Lima, T.C.; de Almeida, R.N.; de Sousa, D.P. Anticonvulsant Essential Oils and Their Relationship with Oxidative Stress in Epilepsy. Biomolecules 2019, 9, 835. [Google Scholar] [CrossRef] [PubMed]
- Hyttinen, J.M.; Kannan, R.; Felszeghy, S.; Niittykoski, M.; Salminen, A.; Kaarniranta, K. The regulation of NFE2L2 (NRF2) signalling and epithelial-to-mesenchymal transition in age-related macular degeneration pathology. Int. J. Mol. Sci. 2019, 20, 5800. [Google Scholar] [CrossRef] [PubMed]
- Rana, A.; Musto, A.E. The role of inflammation in the development of epilepsy. J. Neuroinflammation 2018, 15, 144. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, D.W.; Lee, S.-T.; Byun, J.-I.; Seo, J.-G.; No, Y.J.; Kang, K.W.; Kim, D.; Kim, K.T.; Cho, Y.W. Antiepileptic drug selection according to seizure type in adult patients with epilepsy. J. Clin. Neurol. 2020, 16, 547. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.S.; Cross, J.H.; D’souza, C.; French, J.A.; Haut, S.R.; Higurashi, N.; Hirsch, E.; Jansen, F.E.; Lagae, L.; Moshé, S.L. Instruction manual for the ILAE 2017 operational classification of seizure types. Epilepsia 2017, 58, 531–542. [Google Scholar] [CrossRef]
- Perucca, E. The treatment of the first seizure: The risks. Epilepsia 2008, 49, 29–34. [Google Scholar] [CrossRef]
- Perucca, P.; Gilliam, F.G. Adverse effects of antiepileptic drugs. Lancet Neurol. 2012, 11, 792–802. [Google Scholar] [CrossRef]
- Tsvere, M.; Chiweshe, M.K.; Mutanana, N. General side effects and challenges associated with anti-epilepsy medication: A review of related literature. Afr. J. Prim. Health Care Fam. Med. 2020, 12, 1–5. [Google Scholar]
- Yetisgin, A.A.; Cetinel, S.; Zuvin, M.; Kosar, A.; Kutlu, O. Therapeutic nanoparticles and their targeted delivery applications. Molecules 2020, 25, 2193. [Google Scholar] [CrossRef]
- FEHAID, A.; TANIGUCHI, A. The Double-edged Effect of Silver Nanoparticles is Determined by Their Physical Characteristics. Nano Biomed. 2019, 11, 49–56. [Google Scholar]
- Gupta, J.; Fatima, M.T.; Islam, Z.; Khan, R.H.; Uversky, V.N.; Salahuddin, P. Nanoparticle formulations in the diagnosis and therapy of Alzheimer’s disease. Int. J. Biol. Macromol. 2019, 130, 515–526. [Google Scholar] [CrossRef] [PubMed]
- Khanna, P.K.; Bisht, N.; Phalswal, P. Selenium nanoparticles: A review on synthesis and biomedical applications. Mater. Adv. 2022, 3, 1415–1431. [Google Scholar]
- Yuan, X.; Fu, Z.; Ji, P.; Guo, L.; Al-Ghamdy, A.O.; Alkandiri, A.; Habotta, O.A.; Abdel Moneim, A.E.; Kassab, R.B. Selenium Nanoparticles Pre-Treatment Reverse Behavioral, Oxidative Damage, Neuronal Loss and Neurochemical Alterations in Pentylenetetrazole-Induced Epileptic Seizures in Mice. Int. J. Nanomed. 2020, 15, 6339–6353. [Google Scholar] [CrossRef]
- Ibrahim, H.M.; Zommara, M.A.; Elnaggar, M.E. Ameliorating effect of selenium nanoparticles on cyclophosphamide-induced hippocampal neurotoxicity in male rats: Light, electron microscopic and immunohistochemical study. Folia Morphol. 2021, 80, 806–819. [Google Scholar] [CrossRef]
- Ikram, M.; Javed, B.; Raja, N.I. Biomedical potential of plant-based selenium nanoparticles: A comprehensive review on therapeutic and mechanistic aspects. Int. J. Nanomed. 2021, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Othman, M.S.; Obeidat, S.T.; Al-Bagawi, A.H.; Fareid, M.A.; Fehaid, A.; Abdel Moneim, A.E. Green-synthetized selenium nanoparticles using berberine as a promising anticancer agent. J. Integr. Med. 2022, 20, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Liang, T.; Sun, L.; Meng, L.; Yang, C.; Wang, L.; Li, Q. Green synthesis of selenium nanoparticles with extract of hawthorn fruit induced HepG2 cells apoptosis. Pharm. Biol. 2018, 56, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Anu, K.; Singaravelu, G.; Murugan, K.; Benelli, G. Green-Synthesis of Selenium Nanoparticles Using Garlic Cloves (Allium sativum): Biophysical Characterization and Cytotoxicity on Vero Cells. J. Clust. Sci. 2017, 28, 551–563. [Google Scholar] [CrossRef]
- Rajasekar, S.; Kuppusamy, S. Eco-friendly formulation of selenium nanoparticles and its functional characterization against breast cancer and normal cells. J. Clust. Sci. 2021, 32, 907–915. [Google Scholar] [CrossRef]
- AlBasher, G.; Alfarraj, S.; Alarifi, S.; Alkhtani, S.; Almeer, R.; Alsultan, N.; Alharthi, M.; Alotibi, N.; Al-Dbass, A.; Abdel Moneim, A.E. Nephroprotective Role of Selenium Nanoparticles Against Glycerol-Induced Acute Kidney Injury in Rats. Biol. Trace Elem. Res. 2020, 194, 444–454. [Google Scholar] [CrossRef]
- Bhagwat, S.; Haytowitz, D.B.; Holden, J.M. USDA Database for the Flavonoid Content of Selected Foods, Release 3.1; US Department of Agriculture: Beltsville, MD, USA, 2014. [Google Scholar]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Pervin, M.; Unno, K.; Ohishi, T.; Tanabe, H.; Miyoshi, N.; Nakamura, Y. Beneficial Effects of Green Tea Catechins on Neurodegenerative Diseases. Molecules 2018, 23, 1297. [Google Scholar] [CrossRef] [PubMed]
- Al Omairi, N.E.; Albrakati, A.; Alsharif, K.F.; Almalki, A.S.; Alsanie, W.; Abd Elmageed, Z.Y.; Zaafar, D.; Lokman, M.S.; Bauomy, A.A.; Belal, S.K.; et al. Selenium Nanoparticles with Prodigiosin Rescue Hippocampal Damage Associated with Epileptic Seizures Induced by Pentylenetetrazole in Rats. Biology 2022, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Wang, W.-P.; Mao, Z.-F.; Qu, Z.-Z.; Luan, S.-Q.; Jia, L.-J.; Kan, M.-C. Effects of epigallocatechin-3-gallate on pentylenetetrazole-induced kindling, cognitive impairment and oxidative stress in rats. Neurosci. Lett. 2012, 516, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Kędzierska, E.; Dąbkowska, L.; Obierzyński, P.; Polakowska, M.; Poleszak, E.; Wlaź, P.; Szewczyk, K.; Kotlińska, J. Synergistic Action of Sodium Selenite with some Antidepressants and Diazepam in Mice. Pharmaceutics 2018, 10, 270. [Google Scholar] [CrossRef]
- Dkhil, M.A.; Zrieq, R.; Al-Quraishy, S.; Abdel Moneim, A.E.J.M. Selenium nanoparticles attenuate oxidative stress and testicular damage in streptozotocin-induced diabetic rats. Molecules 2016, 21, 1517. [Google Scholar] [CrossRef]
- Bass, J.S.; Tuo, A.H.; Ton, L.T.; Jankovic, M.J.; Kapadia, P.K.; Schirmer, C.; Krishnan, V. On the digital psychopharmacology of valproic acid in mice. Front. Neurosci. 2020, 14, 594612. [Google Scholar] [CrossRef]
- Van Erum, J.; Van Dam, D.; De Deyn, P.P. PTZ-induced seizures in mice require a revised Racine scale. Epilepsy Behav. 2019, 95, 51–55. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.-Y.; Li, J.-R.; Wang, Y.-Y.; Lin, S.-Y.; Ou, Y.-C.; Lin, C.-J.; Wang, J.-D.; Liao, S.-L.; Chen, C.-J. Indoxyl sulfate caused behavioral abnormality and neurodegeneration in mice with unilateral nephrectomy. Aging 2021, 13, 6681. [Google Scholar] [CrossRef] [PubMed]
- Misra, H.P.; Fridovich, I. The role of superoxide anion in the autoxidation of epinephrine and a simple assay for superoxide dismutase. J. Biol. Chem. 1972, 247, 3170–3175. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [PubMed]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Factor, V.M.; Kiss, A.; Woitach, J.T.; Wirth, P.J.; Thorgeirsson, S.S. Disruption of redox homeostasis in the transforming growth factor-alpha/c-myc transgenic mouse model of accelerated hepatocarcinogenesis. J. Biol. Chem. 1998, 273, 15846–15853. [Google Scholar] [CrossRef]
- Pagel, P.; Blome, J.; Wolf, H.U. High-performance liquid chromatographic separation and measurement of various biogenic compounds possibly involved in the pathomechanism of Parkinson’s disease. J. Chromatogr. B Biomed. Sci. Appl. 2000, 746, 297–304. [Google Scholar] [CrossRef]
- Heinrikson, R.L.; Meredith, S.C. Amino acid analysis by reverse-phase high-performance liquid chromatography: Precolumn derivatization with phenylisothiocyanate. Anal. Biochem. 1984, 136, 65–74. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Khurana, A.; Tekula, S.; Saifi, M.A.; Venkatesh, P.; Godugu, C. Therapeutic applications of selenium nanoparticles. Biomed. Pharmacother. 2019, 111, 802–812. [Google Scholar] [CrossRef]
- Hosnedlova, B.; Kepinska, M.; Skalickova, S.; Fernandez, C.; Ruttkay-Nedecky, B.; Peng, Q.; Baron, M.; Melcova, M.; Opatrilova, R.; Zidkova, J. Nano-selenium and its nanomedicine applications: A critical review. Int. J. Nanomed. 2018, 13, 2107. [Google Scholar] [CrossRef] [PubMed]
- Borowska, M.; Jankowski, K. Green synthesis of selenium nanoparticles: Characterization and application. In Handbook of Greener Synthesis of Nanomaterials and Compounds; Elsevier: Amsterdam, The Netherlands, 2021; pp. 171–190. [Google Scholar]
- Shekh-Ahmad, T.; Kovac, S.; Abramov, A.; Walker, M. Reactive oxygen species in status epilepticus. Epilepsy Behav. 2019, 101, 106410. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Shiraishi, M.; Naito, Y.; Torii, Y.; Nakamura, Y.; Osawa, T. Activation of stress signaling pathways by the end product of lipid peroxidation. 4-hydroxy-2-nonenal is a potential inducer of intracellular peroxide production. J. Biol. Chem. 1999, 274, 2234–2242. [Google Scholar] [CrossRef] [PubMed]
- Banach, M.; Piskorska, B.; Czuczwar, S.J.; Borowicz, K.K. Nitric oxide, epileptic seizures, and action of antiepileptic drugs. CNS Neurol. Disord. Drug Targets 2011, 10, 808–819. [Google Scholar] [CrossRef]
- Abdelfattah, M.S.; Badr, S.E.A.; Lotfy, S.A.; Attia, G.H.; Aref, A.M.; Abdel Moneim, A.E.; Kassab, R.B. Rutin and Selenium Co-administration Reverse 3-Nitropropionic Acid-Induced Neurochemical and Molecular Impairments in a Mouse Model of Huntington’s Disease. Neurotox. Res. 2020, 37, 77–92. [Google Scholar] [CrossRef]
- Mueller, S.G.; Trabesinger, A.H.; Boesiger, P.; Wieser, H.G. Brain glutathione levels in patients with epilepsy measured by in vivo (1)H-MRS. Neurology 2001, 57, 1422–1427. [Google Scholar] [CrossRef]
- Lee, S.H.; Choi, B.Y.; Kho, A.R.; Jeong, J.H.; Hong, D.K.; Lee, S.Y.; Lee, M.W.; Song, H.K.; Choi, H.C.; Suh, S.W. Protective Effects of Protocatechuic Acid on Seizure-Induced Neuronal Death. Int. J. Mol. Sci. 2018, 19, 187. [Google Scholar] [CrossRef]
- Carmona-Aparicio, L.; Pérez-Cruz, C.; Zavala-Tecuapetla, C.; Granados-Rojas, L.; Rivera-Espinosa, L.; Montesinos-Correa, H.; Hernández-Damián, J.; Pedraza-Chaverri, J.; Sampieri, A.I.; Coballase-Urrutia, E. Overview of Nrf2 as therapeutic target in epilepsy. Int. J. Mol. Sci. 2015, 16, 18348–18367. [Google Scholar] [CrossRef]
- Martinc, B.; Grabnar, I.; Vovk, T. The role of reactive species in epileptogenesis and influence of antiepileptic drug therapy on oxidative stress. Curr. Neuropharmacol. 2012, 10, 328–343. [Google Scholar] [CrossRef]
- Font-Nieves, M.; Sans-Fons, M.G.; Gorina, R.; Bonfill-Teixidor, E.; Salas-Pérdomo, A.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. Induction of COX-2 enzyme and down-regulation of COX-1 expression by lipopolysaccharide (LPS) control prostaglandin E2 production in astrocytes. J. Biol. Chem. 2012, 287, 6454–6468. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, J.; Hou, J.; Chen, C. Free radical scavenging efficiency of Nano-Se in vitro. Free. Radic. Biol. Med. 2003, 35, 805–813. [Google Scholar] [CrossRef]
- Ebokaiwe, A.P.; Okori, S.; Nwankwo, J.O.; Ejike, C.E.; Osawe, S.O. Selenium nanoparticles and metformin ameliorate streptozotocin-instigated brain oxidative-inflammatory stress and neurobehavioral alterations in rats. Naunyn Schmiedeberg’s Arch. Pharmacol. 2021, 394, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Hagiu, A.; Attin, T.; Schmidlin, P.R.; Ramenzoni, L.L. Dose-dependent green tea effect on decrease of inflammation in human oral gingival epithelial keratinocytes: In vitro study. Clin. Oral Investig. 2020, 24, 2375–2383. [Google Scholar] [CrossRef] [PubMed]
- Man, G.C.W.; Wang, J.; Song, Y.; Wong, J.H.; Zhao, Y.; Lau, T.S.; Leung, K.T.; Chan, T.H.; Wang, H.; Kwong, J.; et al. Therapeutic potential of a novel prodrug of green tea extract in induction of apoptosis via ERK/JNK and Akt signaling pathway in human endometrial cancer. BMC Cancer 2020, 20, 964. [Google Scholar] [CrossRef] [PubMed]
- Lund, I.V.; Hu, Y.; Raol, Y.H.; Benham, R.S.; Faris, R.; Russek, S.J.; Brooks-Kayal, A.R. BDNF selectively regulates GABAA receptor transcription by activation of the JAK/STAT pathway. Sci. Signal. 2008, 1, ra9. [Google Scholar] [CrossRef] [PubMed]
- Aronica, E.; Ravizza, T.; Zurolo, E.; Vezzani, A. Astrocyte immune responses in epilepsy. Glia 2012, 60, 1258–1268. [Google Scholar] [CrossRef]
- Wang, S.W.; Lee, C.H.; Lin, M.S.; Chi, C.W.; Chen, Y.J.; Wang, G.S.; Liao, K.W.; Chiu, L.P.; Wu, S.H.; Huang, D.M.; et al. ZnO Nanoparticles Induced Caspase-Dependent Apoptosis in Gingival Squamous Cell Carcinoma through Mitochondrial Dysfunction and p70S6K Signaling Pathway. Int. J. Mol. Sci. 2020, 21, 1612. [Google Scholar] [CrossRef]
- Malchow, R.P.; Tchernookova, B.K.; Choi, J.-i.V.; Smith, P.J.; Kramer, R.H.; Kreitzer, M.A. Review and Hypothesis: A Potential Common Link Between Glial Cells, Calcium Changes, Modulation of Synaptic Transmission, Spreading Depression, Migraine, and Epilepsy—H+. Front. Cell. Neurosci. 2021, 15, 693095. [Google Scholar] [CrossRef]
- Rowley, N.M.; Madsen, K.K.; Schousboe, A.; White, H.S. Glutamate and GABA synthesis, release, transport and metabolism as targets for seizure control. Neurochem. Int. 2012, 61, 546–558. [Google Scholar] [CrossRef]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef]
- de Sales Santos, I.M.; Feitosa, C.M.; de Freitas, R.M. Pilocarpine-induced seizures produce alterations on choline acetyltransferase and acetylcholinesterase activities and deficit memory in rats. Cell. Mol. Neurobiol. 2010, 30, 569–575. [Google Scholar] [CrossRef] [PubMed]
- Schimidt, H.L.; Garcia, A.; Martins, A.; Mello-Carpes, P.B.; Carpes, F.P. Green tea supplementation produces better neuroprotective effects than red and black tea in Alzheimer-like rat model. Food Res. Int. 2017, 100, 442–448. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Wu, X.; Li, D.; Liu, P.; Zhang, S.; Gao, D.; Gao, F.; Zhang, M.; Xiao, Y. Protective effects of chondroitin sulphate nano-selenium on a mouse model of Alzheimer’s disease. Int. J. Biol. Macromol. 2020, 154, 233–245. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Forward (5′–-3′) | Reverse (3′–-5′) |
|---|---|---|---|
| Nrf2 | NM_001399173.1 | CCTCAGCATGATGGACTTGGA | GCGACTGAAATGTAGGTGAAGA |
| iNOS | NM_012611.3 | GGTGAGGGGACTGGACTTTTAG | TTGTTGGGCTGGGAATAGCA |
| NMDAR | NM_012573.4 | GGCAACTTGTATGGGAGCCT | ATTTACCGCCTGTGATGGCA |
| β-actin | NM_031144.3 | GTCCACCCGCGAGTACAAC | GGATGCCTCTCTTGCTCTGG |
| Parameters | Control | PTZ | EGCG + PTZ | Na2SeO3 + PTZ | SeNPs + PTZ | VPA + PTZ |
|---|---|---|---|---|---|---|
| 5-HT (μg/g tissue) | 12.63 ± 1.27 | 6.39 ± 0.94 # | 8.61 ± 0.2.27 #$ | 10.79 ± 1.56 #$ | 11.93 ± 1.19 $ | 13.25 ± 1.98 $ |
| DA (μg/g tissue) | 0.42 ± 0.08 | 0.15 ± 0.02 # | 0.34 ± 0.09 #$ | 0.38 ± 0.09 $ | 0.43 ± 0.07 $ | 0.49 ± 0.09 $ |
| NE (μg/g tissue) | 0.55 ± 0.09 | 0.27 ± 0.04 # | 0.35 ± 0.08 #$ | 0.46 ± 0.07 #$ | 0.54 ± 0.08 $ | 0.53 ± 0.09 $ |
| GABA (μg/g tissue) | 884.0 ± 71.70 | 624.4 ± 109.5 # | 732.3 ± 210.8 $ | 813.5 ± 143.9 $ | 875.2 ± 72.4 $ | 821.5 ± 139.3 $ |
| AChE (µmol/min/mg protein) | 9.79 ± 1.83 | 5.08 ± 1.52 # | 6.16 ± 1.95 #$ | 9.38 ± 1.84 $ | 8.64 ± 1.27 #$ | 8.97 ± 1.30 $ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alrashdi, B.M.; Fehaid, A.; Kassab, R.B.; Rizk, S.; Habotta, O.A.; Abdel Moneim, A.E. Biosynthesized Selenium Nanoparticles Using Epigallocatechin Gallate Protect against Pentylenetetrazole-Induced Acute Epileptic Seizures in Mice via Antioxidative, Anti-Inflammatory, and Anti-Apoptotic Activities. Biomedicines 2023, 11, 1955. https://doi.org/10.3390/biomedicines11071955
Alrashdi BM, Fehaid A, Kassab RB, Rizk S, Habotta OA, Abdel Moneim AE. Biosynthesized Selenium Nanoparticles Using Epigallocatechin Gallate Protect against Pentylenetetrazole-Induced Acute Epileptic Seizures in Mice via Antioxidative, Anti-Inflammatory, and Anti-Apoptotic Activities. Biomedicines. 2023; 11(7):1955. https://doi.org/10.3390/biomedicines11071955
Chicago/Turabian StyleAlrashdi, Barakat M., Alaa Fehaid, Rami B. Kassab, Sara Rizk, Ola A. Habotta, and Ahmed E. Abdel Moneim. 2023. "Biosynthesized Selenium Nanoparticles Using Epigallocatechin Gallate Protect against Pentylenetetrazole-Induced Acute Epileptic Seizures in Mice via Antioxidative, Anti-Inflammatory, and Anti-Apoptotic Activities" Biomedicines 11, no. 7: 1955. https://doi.org/10.3390/biomedicines11071955
APA StyleAlrashdi, B. M., Fehaid, A., Kassab, R. B., Rizk, S., Habotta, O. A., & Abdel Moneim, A. E. (2023). Biosynthesized Selenium Nanoparticles Using Epigallocatechin Gallate Protect against Pentylenetetrazole-Induced Acute Epileptic Seizures in Mice via Antioxidative, Anti-Inflammatory, and Anti-Apoptotic Activities. Biomedicines, 11(7), 1955. https://doi.org/10.3390/biomedicines11071955