
A Dual Role of Osteopontin in Modifying B Cell Responses



Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Tissue
2.2. Immunohistochemistry
2.3. B Cell Isolation and Cell Culture
2.4. Stimulation of B Cells with rOPN
2.5. Flow Cytometry
2.5.1. Purity Check of B Cells
2.5.2. B Cell Activation
2.6. Antibody Array and ELISA
2.7. Aggregation Assay
2.8. Statistical Analysis
3. Results
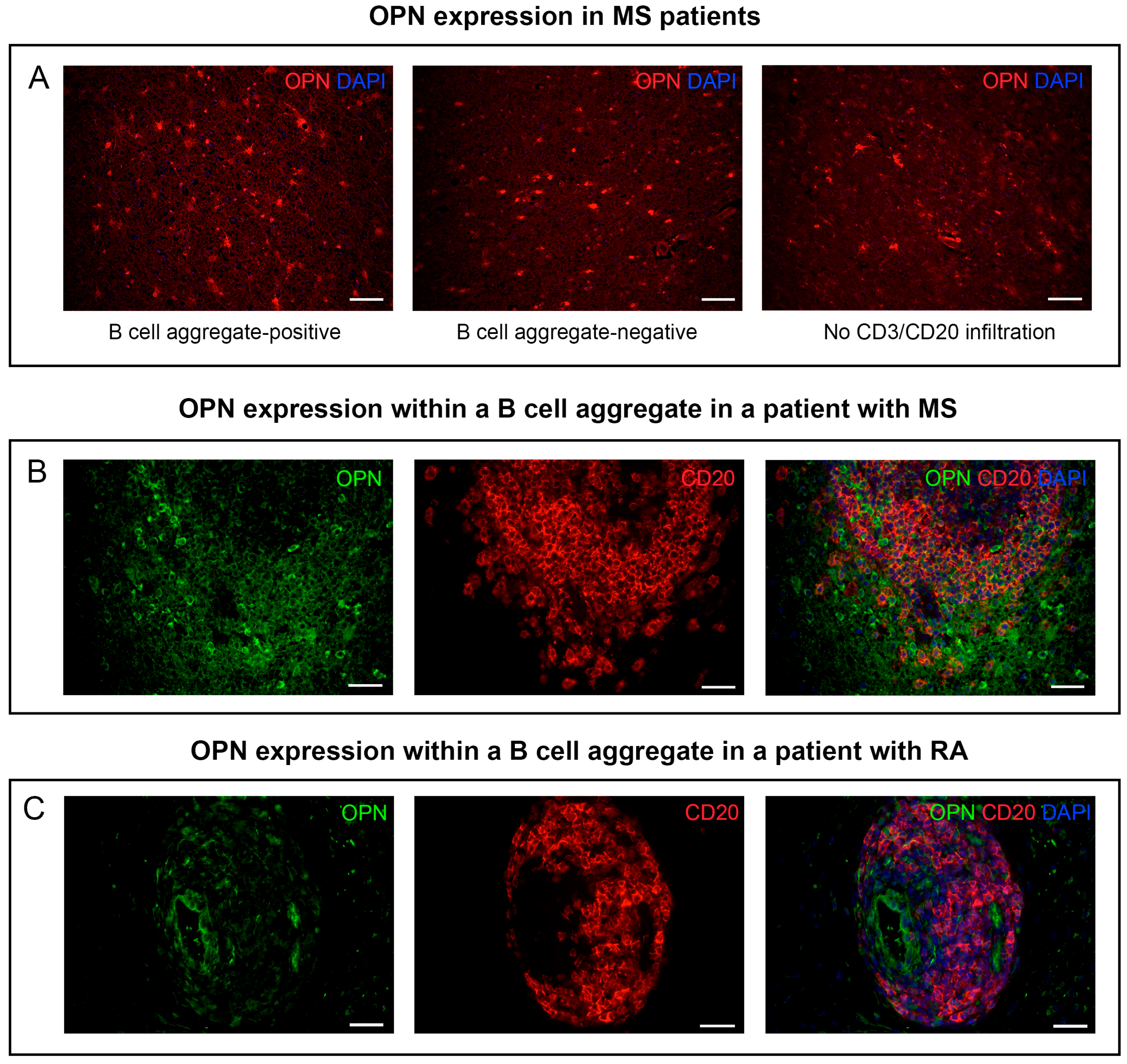
3.1. OPN Is Expressed within B Cell Aggregates in Human Tissues
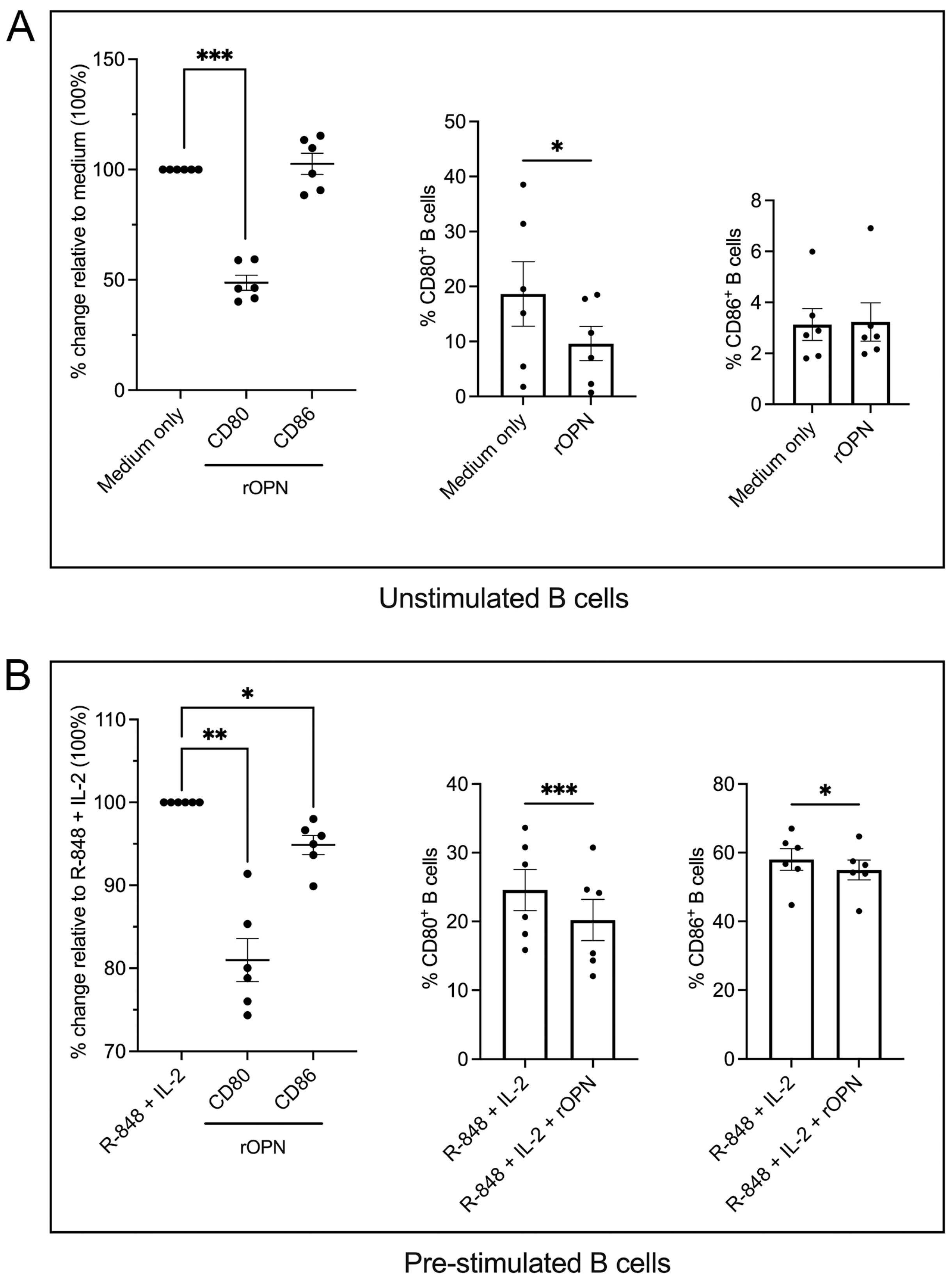
3.2. B Cells Treated with OPN Downregulate the Co-Stimulatory Molecules CD80 and CD86
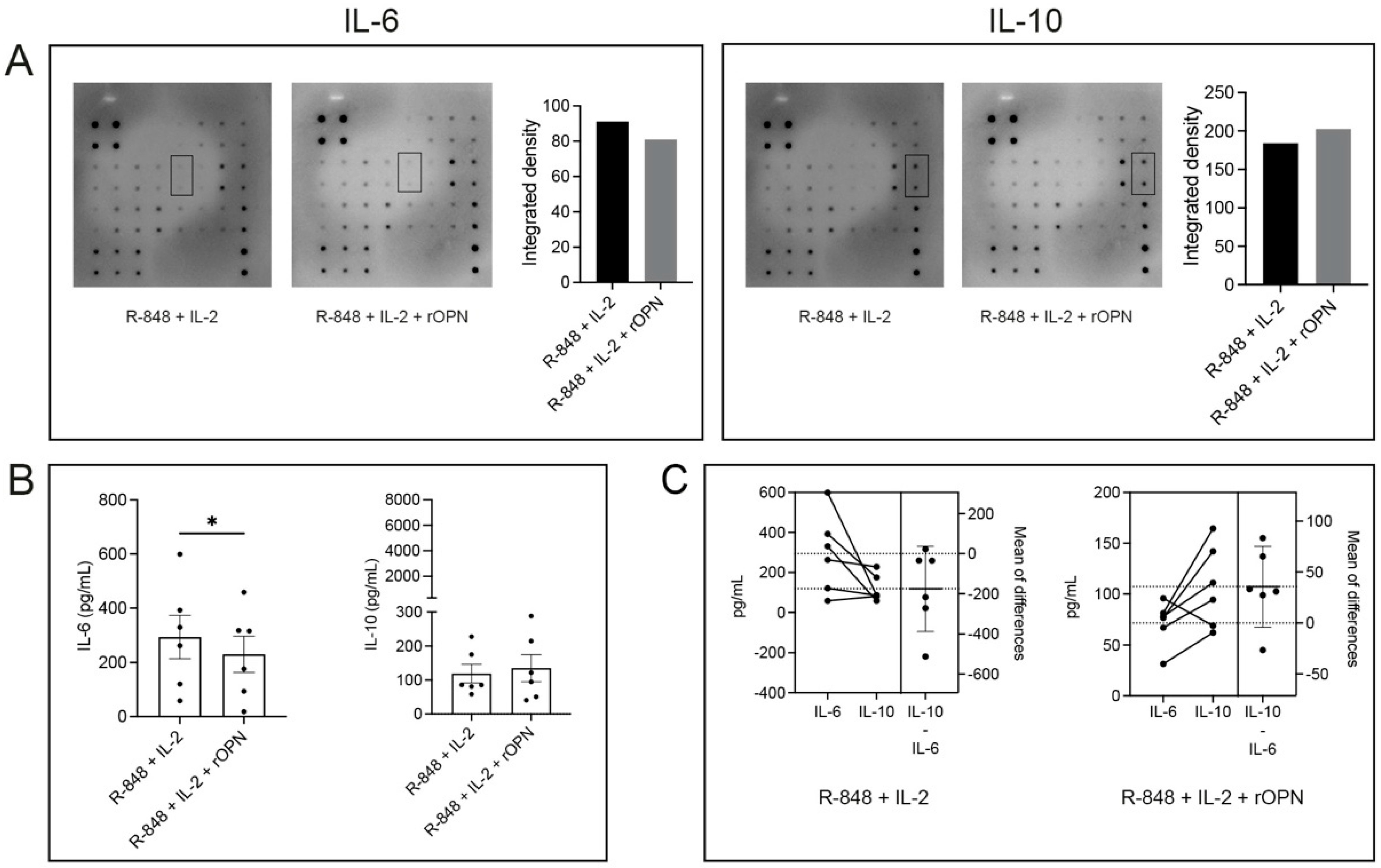
3.3. OPN Has Differential Effects on IL-6 and IL-10 Secretion by B Cells
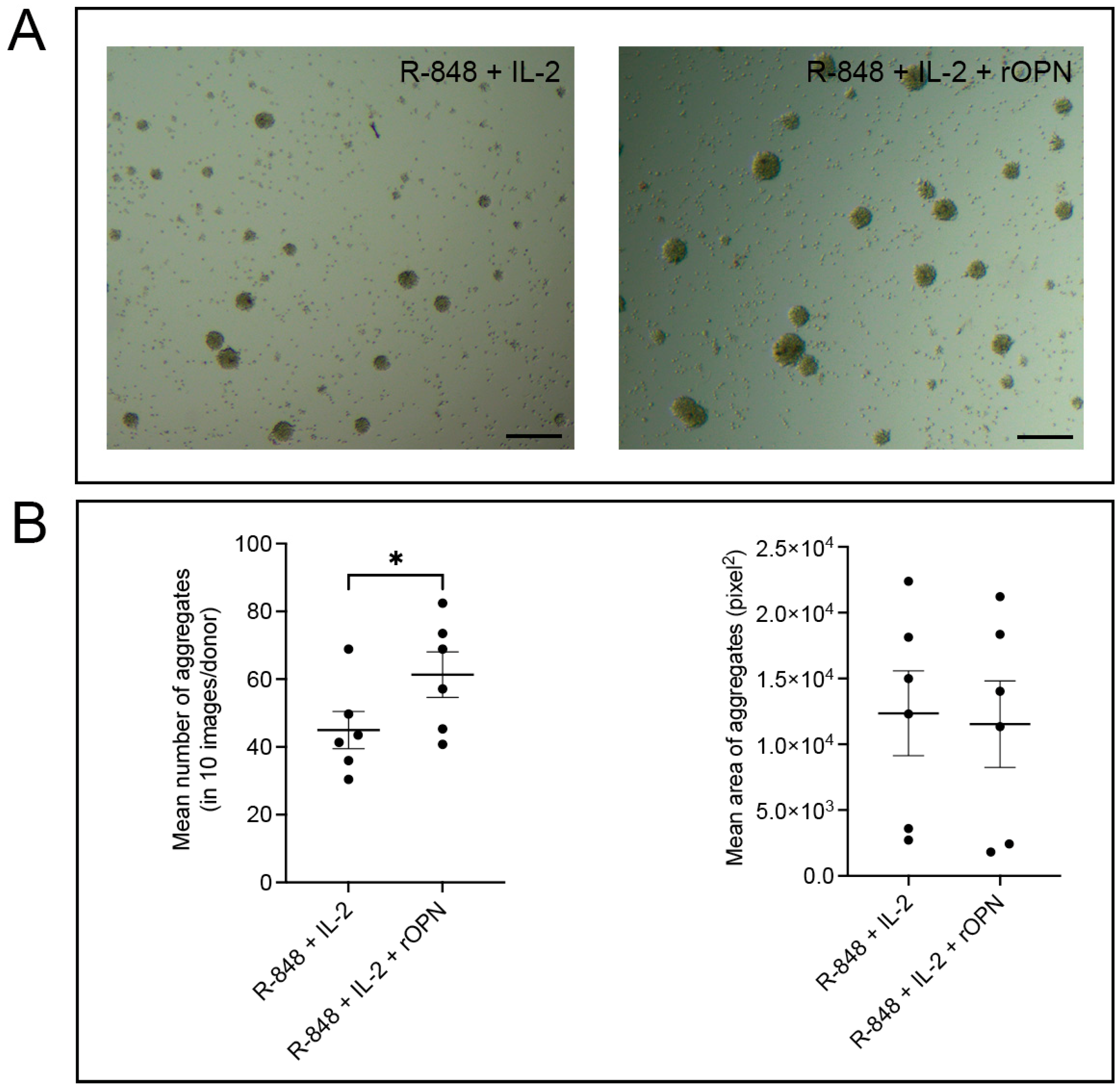
3.4. OPN Significantly Increases the Potential of B Cells to Form Aggregates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jain, R.W.; Yong, V.W. B cells in central nervous system disease: Diversity, locations and pathophysiology. Nat. Rev. Immunol. 2022, 22, 513–524. [Google Scholar] [CrossRef]
- Corsiero, E.; Nerviani, A.; Bombardieri, M.; Pitzalis, C. Ectopic lymphoid structures: Powerhouse of autoimmunity. Front. Immunol. 2016, 7, 430. [Google Scholar] [CrossRef] [Green Version]
- Serafini, B.; Rosicarelli, B.; Magliozzi, R.; Stigliano, E.; Aloisi, F. Detection of ectopic B-cell follicles with germinal centers in the meninges of patients with secondary progressive multiple sclerosis. Brain Pathol. 2004, 14, 164–174. [Google Scholar] [CrossRef]
- Magliozzi, R.; Howell, O.; Vora, A.; Serafini, B.; Nicholas, R.; Puopolo, M.; Reynolds, R.; Aloisi, F. Meningeal B-cell follicles in secondary progressive multiple sclerosis associate with early onset of disease and severe cortical pathology. Brain 2007, 130, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Howell, O.W.; Reeves, C.A.; Nicholas, R.; Carassiti, D.; Radotra, B.; Gentleman, S.M.; Serafini, B.; Aloisi, F.; Roncaroli, F.; Magliozzi, R.; et al. Meningeal inflammation is widespread and linked to cortical pathology in multiple sclerosis. Brain 2011, 134, 2755–2771. [Google Scholar] [CrossRef] [Green Version]
- Lucchinetti, C.F.; Popescu, B.F.G.; Bunyan, R.F.; Moll, N.M.; Roemer, S.F.; Lassmann, H.; Brück, W.; Parisi, J.E.; Scheithauer, B.W.; Giannini, C.; et al. Inflammatory cortical demyelination in early multiple sclerosis. N. Engl. J. Med. 2011, 365, 2188–2197. [Google Scholar] [CrossRef] [Green Version]
- Machado-Santos, J.; Saji, E.; Tröscher, A.; Paunovic, M.; Liblau, R.; Gabriely, G.; Bien, C.G.; Bauer, J.; Lassmann, H. The compartmentalized inflammatory response in the multiple sclerosis brain is composed of tissue-resident CD8+ T lymphocytes and B cells. Brain 2018, 141, 2066–2082. [Google Scholar] [CrossRef] [PubMed]
- Moccia, M.; Haider, L.; Eshaghi, A.; van de Pavert, S.H.P.; Morra, V.B.; Patel, A.; Wheeler-Kingshott, C.A.M.; Barkhof, F.; Ciccarelli, O. B cells in the CNS at postmortem are associated with worse outcome and cell types in multiple sclerosis. Neurol.—Neuroimmunol. Neuroinflamm. 2022, 9, e1108. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Howell, O.W.; Carassiti, D.; Magliozzi, R.; Gveric, D.; Muraro, P.A.; Nicholas, R.; Roncaroli, F.; Reynolds, R. Meningeal inflammation plays a role in the pathology of primary progressive multiple sclerosis. Brain 2012, 135, 2925–2937. [Google Scholar] [CrossRef] [Green Version]
- Reali, C.; Magliozzi, R.; Roncaroli, F.; Nicholas, R.; Howell, O.W.; Reynolds, R. B cell rich meningeal inflammation associates with increased spinal cord pathology in multiple sclerosis. Brain Pathol. 2020, 30, 779–793. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.M.; Fransen, N.L.; Touil, H.; Michailidou, I.; Huitinga, I.; Gommerman, J.L.; Bar-Or, A.; Ramaglia, V. Accumulation of meningeal lymphocytes correlates with white matter lesion activity in progressive multiple sclerosis. JCI Insight 2022, 7, e151683. [Google Scholar] [CrossRef] [PubMed]
- Chunder, R.; Schropp, V.; Kuerten, S. B cells in multiple sclerosis and virus-induced neuroinflammation. Front. Neurol. 2020, 11, 591894. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.W.; Jones, S.A. Ectopic lymphoid follicles: Inducible centers for generating antigen-specific immune responses within tissues. Immunology 2016, 147, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Negron, A.; Stüve, O.; Forsthuber, T.G. Ectopic lymphoid follicles in multiple sclerosis: Centers for disease control? Front. Neurol. 2020, 11, 607766. [Google Scholar] [CrossRef] [PubMed]
- Pikor, N.B.; Prat, A.; Bar-Or, A.; Gommerman, J.L. Meningeal tertiary lymphoid tissues and multiple sclerosis: A gathering place for diverse types of immune cells during CNS autoimmunity. Front. Immunol. 2016, 6, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uccelli, A.; Aloisi, F.; Pistoia, V. Unveiling the enigma of the CNS as a B-cell fostering environment. Trends Immunol. 2005, 26, 254–259. [Google Scholar] [CrossRef]
- Sodek, J.; Ganss, B.; McKee, M.D. Osteopontin. Crit. Rev. Oral Biol. Med. 2000, 11, 279–303. [Google Scholar] [CrossRef]
- Steinman, L. A molecular trio in relapse and remission in multiple sclerosis. Nat. Rev. Immunol. 2009, 9, 440–447. [Google Scholar] [CrossRef]
- Cantor, H.; Shinohara, M.L. Regulation of T-helper-cell lineage development by osteopontin: The inside story. Nat. Rev. Immunol. 2009, 9, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Jansson, M.; Panoutsakopoulou, V.; Baker, J.; Klein, L.; Cantor, H. Cutting Edge: Attenuated experimental autoimmune encephalomyelitis in Eta-1/osteopontin-deficient mice. J. Immunol. 2002, 168, 2096–2099. [Google Scholar] [CrossRef]
- Hur, E.M.; Youssef, S.; Haws, M.E.; Zhang, S.Y.; Sobel, R.A.; Steinman, L. Osteopontin-induced relapse and progression of autoimmune brain disease through enhanced survival of activated T cells. Nat. Immunol. 2007, 8, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, C.-C.; Engelenburg, H.J.; Jongejan, A.; Zhu, J.; Zhang, B.; Mingueneau, M.; Moerland, P.D.; Huitinga, I.; Smolders, J.; Hamann, J. Osteopontin associates with brain TRM-cell transcriptome and compartmentalization in donors with and without multiple sclerosis. iScience 2023, 26, 105785. [Google Scholar] [CrossRef] [PubMed]
- Guo, B.; Tumang, J.R.; Rothstein, T.L. B cell receptor crosstalk: B cells express osteopontin through the combined action of the alternate and classical BCR signaling pathways. Mol. Immunol. 2009, 46, 587–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rittling, S.R.; Singh, R. Osteopontin in immune-mediated diseases. J. Dent. Res. 2015, 94, 1638–1645. [Google Scholar] [CrossRef] [Green Version]
- Chunder, R.; Schropp, V.; Jabari, S.; Marzin, M.; Amor, S.; Kuerten, S. Identification of a novel role for matrix metalloproteinase-3 in the modulation of B cell responses in multiple sclerosis. Front. Immunol. 2022, 13, 1025377. [Google Scholar] [CrossRef]
- Wada, T.; McKee, M.D.; Steitz, S.; Giachelli, C.M. Calcification of vascular smooth muscle cell cultures: Inhibition by osteopontin. Circ. Res. 1999, 84, 166–178. [Google Scholar] [CrossRef]
- Rabenstein, M.; Hucklenbroich, J.; Willuweit, A.; Ladwig, A.; Fink, G.R.; Schroeter, M.; Langen, K.-J.; Rueger, M.A. Osteopontin mediates survival, proliferation and migration of neural stem cells through the chemokine receptor CXCR4. Stem Cell Res. Ther. 2015, 6, 99. [Google Scholar] [CrossRef] [Green Version]
- Rovituso, D.M.; Scheffler, L.; Wunsch, M.; Kleinschnitz, C.; Dörck, S.; Ulzheimer, J.; Bayas, A.; Steinman, L.; Ergün, S.; Kuerten, S. CEACAM1 mediates B cell aggregation in central nervous system autoimmunity. Sci. Rep. 2016, 20, 29847. [Google Scholar] [CrossRef] [Green Version]
- Take, Y.; Nakata, K.; Hashimoto, J.; Tsuboi, H.; Nishimoto, N.; Ochi, T.; Yoshikawa, H. Specifically modified osteopontin in rheumatoid arthritis fibroblast-like synoviocytes supports interaction with B cells and enhances production of interleukin-6. Arthritis Rheum. 2009, 60, 3591–3601. [Google Scholar] [CrossRef]
- Bar-Or, A.; Oliveira, E.M.; Anderson, D.E.; Krieger, J.I.; Duddy, M.; O’Connor, K.C.; Hafler, D.A. Immunological memory: Contribution of memory B cells expressing costimulatory molecules in the resting state. J. Immunol. 2001, 167, 5669–5677. [Google Scholar] [CrossRef]
- Hathcock, K.S.; Laszlo, G.; Pucillo, C.; Linsley, P.; Hodes, R.J. Comparative analysis of B7-1 and B7-2 costimulatory ligands: Expression and function. J. Exp. Med. 1994, 180, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Guerrier, T.; Labalette, M.; Launay, D.; Lee-Chang, C.; Outteryck, O.; Lefèvre, G.; Vermersch, P.; Dubucquoi, S.; Zéphir, H. Proinflammatory B-cell profile in the early phases of MS predicts an active disease. Neurol.—Neuroimmunol. Neuroinflamm. 2017, 5, e431. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.H.; Rigley, K.P.; Callard, R.E. Activation of human B cells through the CD19 surface antigen results in homotypic adhesion by LFA-1-dependent and -independent mechanisms. Immunology 1991, 73, 293–297. [Google Scholar] [PubMed]
- Stott, D.I.; Hiepe, F.; Hummel, M.; Steinhauser, G.; Berek, C. Antigen-driven clonal proliferation of B cells within the target tissue of an autoimmune disease. The salivary glands of patients with Sjögren’s syndrome. J. Clin. Investig. 1998, 102, 938–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlsen, H.S.; Baekkevold, E.S.; Johansen, F.E.; Haraldsen, G.; Brandtzaeg, P. B cell attracting chemokine 1 (CXCL13) and its receptor CXCR5 are expressed in normal and aberrant gut associated lymphoid tissue. Gut 2002, 51, 364–371. [Google Scholar] [CrossRef] [Green Version]
- Lund, S.A.; Giachelli, C.M.; Scatena, M. The role of osteopontin in inflammatory processes. J. Cell Commun. Signal. 2009, 3, 311–322. [Google Scholar] [CrossRef] [Green Version]
- DiSano, K.D.; Gilli, F.; Pachner, A.R. Memory B cells in multiple sclerosis: Emerging players in disease pathogenesis. Front. Immunol. 2021, 12, 676686. [Google Scholar] [CrossRef]
- Harp, C.T.; Ireland, S.; Davis, L.S.; Remington, G.; Cassidy, B.; Cravens, P.D.; Stuve, O.; Lovett-Racke, A.E.; Eagar, T.N.; Greenberg, B.M.; et al. Memory B cells from a subset of treatment-naïve relapsing-remitting multiple sclerosis patients elicit CD4+ T-cell proliferation and IFN-γ production in response to myelin basic protein and myelin oligodendrocyte glycoprotein. Eur. J. Immunol. 2010, 40, 2942–2956. [Google Scholar] [CrossRef] [Green Version]
- Duddy, M.; Niino, M.; Adatia, F.; Hebert, S.; Freedman, M.; Atkins, H.; Kim, H.J.; Bar-Or, A. Distinct effector cytokine profiles of memory and naive human B cell subsets and implication in multiple sclerosis. J. Immunol. 2007, 178, 6092–6099. [Google Scholar] [CrossRef]
- Jelcic, I.; Al Nimer, F.; Wang, J.; Lentsch, V.; Planas, R.; Jelcic, I.; Madjovski, A.; Ruhrmann, S.; Faigle, W.; Frauenknecht, K.; et al. Memory B cells activate brain-homing, autoreactive CD4+ T cells in multiple sclerosis. Cell 2018, 175, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Pinna, D.; Corti, D.; Jarrossay, D.; Sallusto, F.; Lanzavecchia, A. Clonal dissection of the human memory B-cell repertoire following infection and vaccination. Eur. J. Immunol. 2009, 39, 1260–1270. [Google Scholar] [CrossRef] [PubMed]
- Fraussen, J.; Claes, N.; Van Wijmeersch, B.; van Horssen, J.; Stinissen, P.; Hupperts, R.; Somers, V. B cells of multiple sclerosis patients induce autoreactive proinflammatory T cell responses. Clin. Immunol. 2016, 173, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Menezes, S.M.; Decanine, D.; Brassat, D.; Khouri, R.; Schnitman, S.V.; Kruschewsky, R.; López, G.; Alvarez, C.; Talledo, M.; Gotuzzo, E.; et al. CD80+ and CD86+ B cells as biomarkers and possible therapeutic targets in HTLV-1 associated myelopathy/tropical spastic paraparesis and multiple sclerosis. J. Neuroinflamm. 2014, 11, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arkatkar, T.; Du, S.W.; Jacobs, H.M.; Dam, E.M.; Hou, B.; Buckner, J.H.; Rawlings, D.J.; Jackson, S.W. B cell-derived IL-6 initiates spontaneous germinal center formation during systemic autoimmunity. J. Exp. Med. 2017, 214, 3207–3217. [Google Scholar] [CrossRef]
- Maeda, K.; Mehta, H.; Drevets, D.A.; Coggeshall, K.M. IL-6 increases B-cell IgG production in a feed-forward proinflammatory mechanism to skew hematopoiesis and elevate myeloid production. Blood 2010, 115, 4699–4706. [Google Scholar] [CrossRef] [Green Version]
- Ran, Z.; Yue-Bei, L.; Qiu-Ming, Z.; Huan, Y. Regulatory B cells and its role in central nervous system inflammatory demyelinating diseases. Front. Immunol. 2020, 11, 1884. [Google Scholar] [CrossRef]
- Radomir, L.; Kramer, M.P.; Perpinial, M.; Schottlender, N.; Rabani, S.; David, K.; Wiener, A.; Lewinsky, H.; Becker-Herman, S.; Aharoni, R.; et al. The survival and function of IL-10-producing regulatory B cells are negatively controlled by SLAMF5. Nat. Commun. 2021, 12, 1893. [Google Scholar] [CrossRef] [PubMed]
- Renkl, A.C.; Wussler, J.; Ahrens, T.; Thoma, K.; Kon, S.; Uede, T.; Martin, S.F.; Simon, J.C.; Weiss, J.M. Osteopontin functionally activates dendritic cells and induces their differentiation toward a Th1-polarizing phenotype. Blood 2005, 106, 946–955. [Google Scholar] [CrossRef] [Green Version]
- Chabas, D.; Baranzini, S.E.; Mitchell, D.; Bernard, C.C.A.; Rittling, S.R.; Denhardt, D.T.; Sobel, R.A.; Lock, C.; Karpuj, M.; Pedotti, R.; et al. The influence of the proinflammatory cytokine, osteopontin, on autoimmune demyelinating disease. Science 2001, 294, 1731–1735. [Google Scholar] [CrossRef]
- Weber, G.F.; Ashkar, S.; Glimcher, M.J.; Cantor, H. Receptor-ligand interaction between CD44 and osteopontin (Eta-1). Science 1996, 271, 509–512. [Google Scholar] [CrossRef]
- Kee, R.; Naughton, M.; McDonnell, G.V.; Howell, O.W.; Fitzgerald, D.C. A review of compartmentalised inflammation and tertiary lymphoid structures in the pathophysiology of multiple sclerosis. Biomedicines 2022, 10, 2604. [Google Scholar] [CrossRef] [PubMed]
- Kahles, F.; Findeisen, H.M.; Bruemmer, D. Osteopontin: A novel regulator at the cross roads of inflammation, obesity and diabetes. Mol. Metab. 2014, 3, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Clemente, N.; Raineri, D.; Cappellano, G.; Boggio, E.; Favero, F.; Soluri, M.F.; Dianzani, C.; Comi, C.; Dianzani, U.; Chiocchetti, A. Osteopontin bridging innate and adaptive immunity in autoimmune diseases. J. Immunol. Res. 2016, 2016, 7675437. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MS patients | |||||||
| Number | Age | Sex | MS Type | Disease Duration (In Years) | Cause of Death | Number of Blocks | Patient Type |
| MS325 | 51 | M | PPMS | 2 | Bronchopneumonia | 2 | NI |
| MS342 | 35 | F | SPMS | 5 | MS | 2 | B cell |
| MS402 | 46 | M | SPMS | 20 | MS, bronchopneumonia | 1 | B cell |
| MS407 | 44 | F | SPMS | 19 | Sepsis, pneumonia | 1 | B cell |
| MS408 | 39 | M | SPMS | 10 | Sepsis, pneumonia | 1 | NI |
| MS438 | 53 | F | Unknown | 18 | MS | 2 | NI |
| MS444 | 49 | M | SPMS | 20 | Kidney failure | 2 | T cell |
| MS473 | 39 | F | PPMS | 13 | MS, bronchopneumonia | 2 | T cell |
| MS485 | 57 | F | PPMS | 29 | MS, bronchopneumonia | 2 | T cell |
| MS510 | 38 | F | SPMS | 22 | MS, pneumonia | 1 | T cell |
| MS523 | 63 | F | SPMS | 32 | Bronchopneumonia | 1 | T cell |
| MS528 | 45 | F | SPMS | 25 | Pneumonia | 1 | T cell |
| 11-077 | 66 | F | PPMS | 32 | Euthanasia | 5 | B cell |
| 14-006 | 56 | F | Unknown | 31 | Suicide by pentobarbital intoxication | 2 | B cell |
| 15-047 | 50 | F | SPMS | 18 | Euthanasia | 2 | B cell |
| 16-019 | 48 | M | Unknown | 15 | Ileus, dehydration | 4 | B cell |
| RA patients | |||||||
| Number | Age | Sex | Tissue type | Number of blocks | |||
| 427-19 | 57 | F | Surgical biopsy | 1 | |||
| 470-19a | 74 | F | Surgical biopsy | 1 | |||
| 471-19b | 82 | F | Surgical biopsy | 1 | |||
| Antibodies Used for Immunofluorescence | ||||
|---|---|---|---|---|
| Antibody | Source | Host Species | Clone | Dilution |
| Anti-CD20 | Invitrogen | Mouse | L26 | 1:400 |
| Anti-CD3 | Abcam | Rabbit | SP162 | 1:150 |
| Anti-OPN | Polyclonal | 1:100 | ||
| Anti-human IgG | Abcam | Rabbit | Polyclonal | 1:200 |
| Antibodies used for flow cytometry | ||||
| BB700 Anti-CD3 | BD Biosciences | Mouse | SK7 | 1:20 |
| APC Anti-CD19 | HIB19 | 1:5 | ||
| APC Anti-CD20 | 2H7 | 1:5 | ||
| BB515 Anti-CD45 | HI30 | 1:20 | ||
| BV605 Anti-CD80 | 2D10.4 | 1:40 | ||
| BB515 Anti-CD86 | FUN-1 | 1:20 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chunder, R.; Schropp, V.; Marzin, M.; Amor, S.; Kuerten, S. A Dual Role of Osteopontin in Modifying B Cell Responses. Biomedicines 2023, 11, 1969. https://doi.org/10.3390/biomedicines11071969
Chunder R, Schropp V, Marzin M, Amor S, Kuerten S. A Dual Role of Osteopontin in Modifying B Cell Responses. Biomedicines. 2023; 11(7):1969. https://doi.org/10.3390/biomedicines11071969
Chicago/Turabian StyleChunder, Rittika, Verena Schropp, Manuel Marzin, Sandra Amor, and Stefanie Kuerten. 2023. "A Dual Role of Osteopontin in Modifying B Cell Responses" Biomedicines 11, no. 7: 1969. https://doi.org/10.3390/biomedicines11071969
APA StyleChunder, R., Schropp, V., Marzin, M., Amor, S., & Kuerten, S. (2023). A Dual Role of Osteopontin in Modifying B Cell Responses. Biomedicines, 11(7), 1969. https://doi.org/10.3390/biomedicines11071969