
Tumor-Associated Macrophages Affect the Tumor Microenvironment and Radioresistance via the Upregulation of CXCL6/CXCR2 in Hepatocellular Carcinoma

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. HCC Cell Culture
2.2. The Cancer Genome Atlas and Gene Expression Omnibus Database Extraction
2.3. Cell Viability Assay
2.4. Western Blotting
2.5. Immunohistochemistry Staining
2.6. Immunofluorescence (IFC) Staining
2.7. RNA Extraction and Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction (RT-PCR)
2.8. Statistical Analysis
3. Results
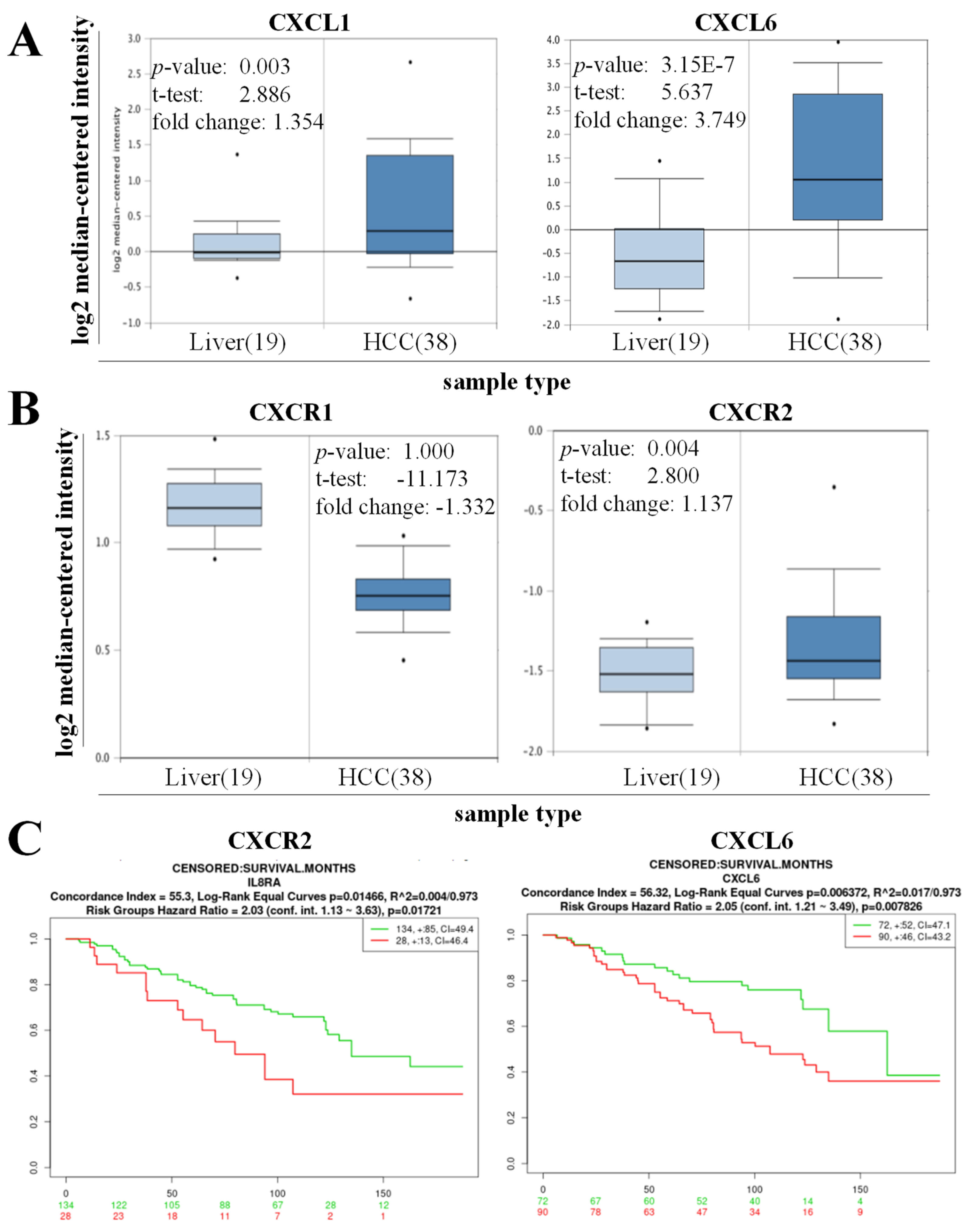
3.1. High Expression of CXCL6 and CXCR2 Was Associated with Hepatogenesis and Reduced the Overall Survival of Patients with HCC
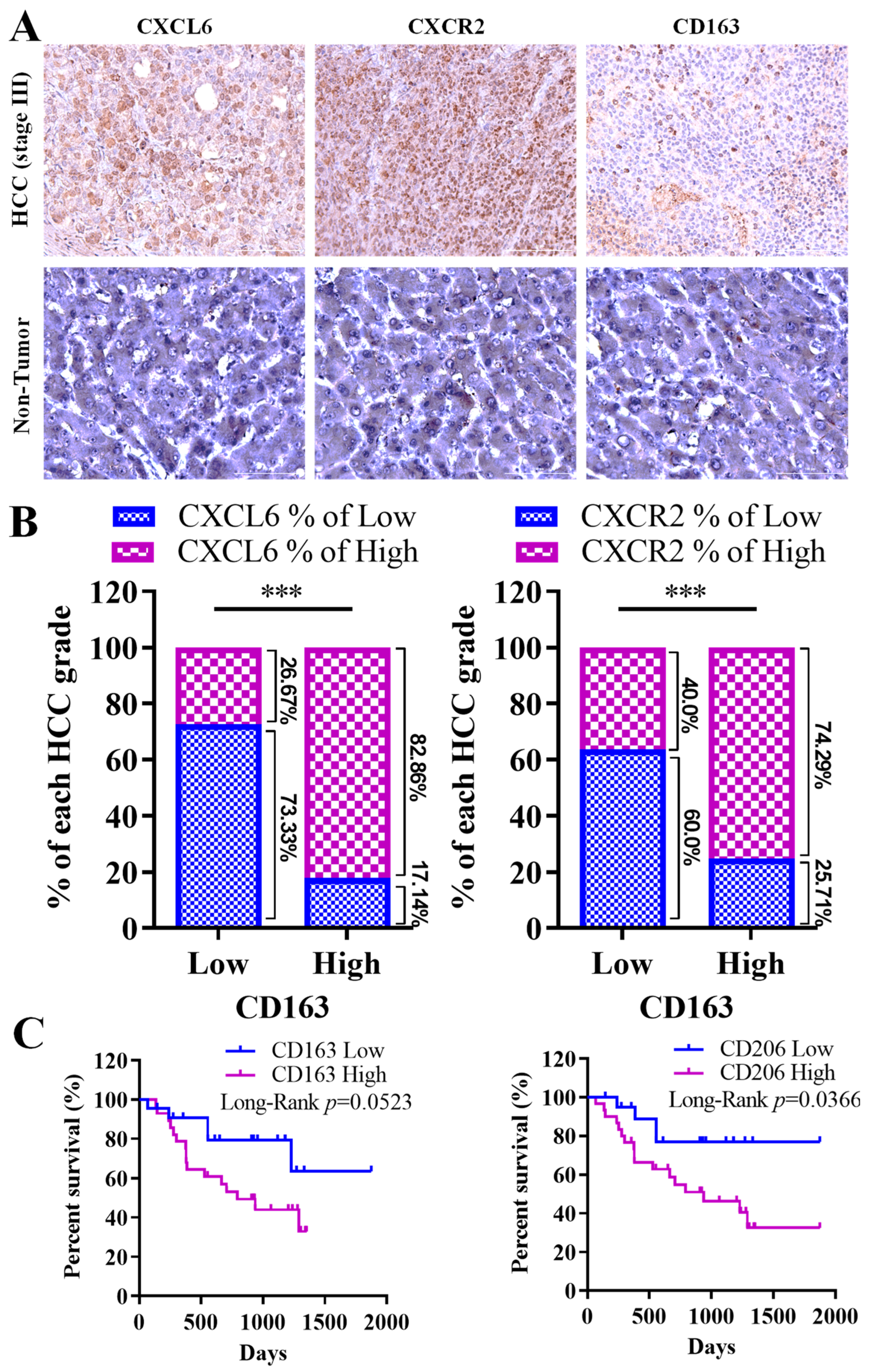
3.2. Strong Link between CXCL6 and CXCR2 Expression, M2 Macrophage Infiltration, and Unfavorable Prognosis in Hepatocellular Carcinoma Progression
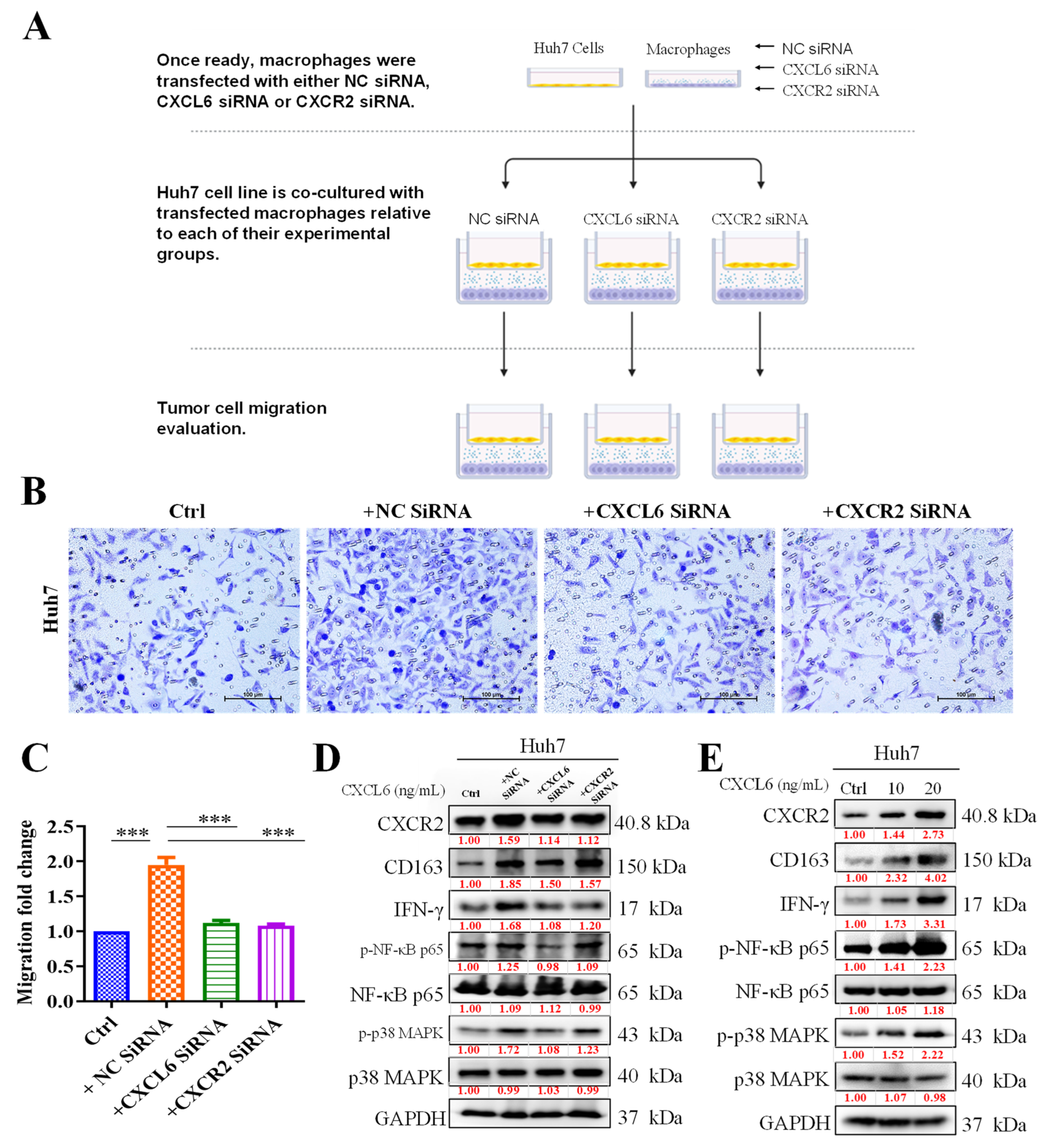
3.3. TAMs Promoted HCC Cell Proliferation and Migration by Activating CXCR2/IFN-g/p38 MAPK/NF-κB Signaling
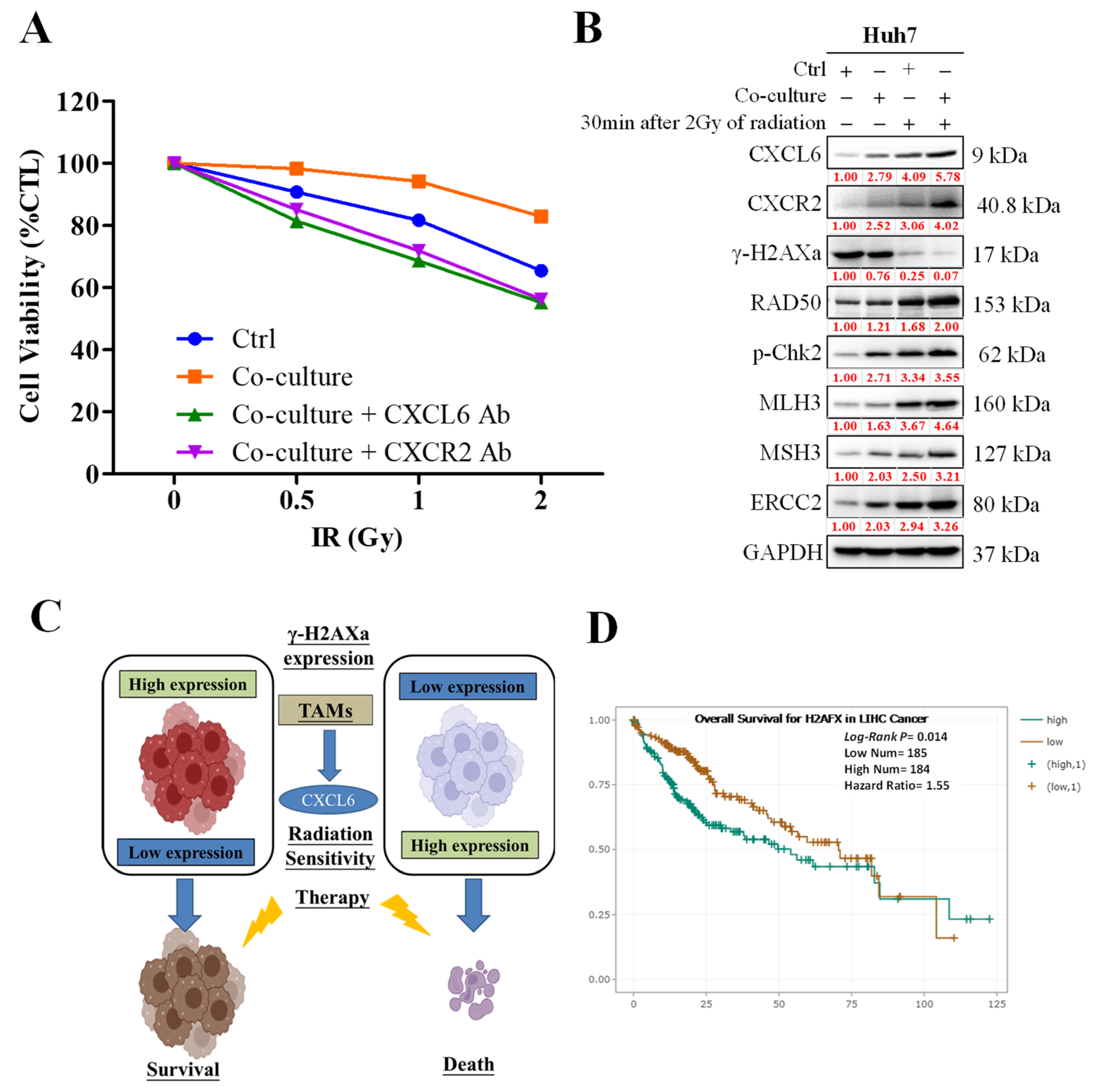
3.4. CXCL6 Conferred Radioresistance via the Enhancement of DNA Damage Repair
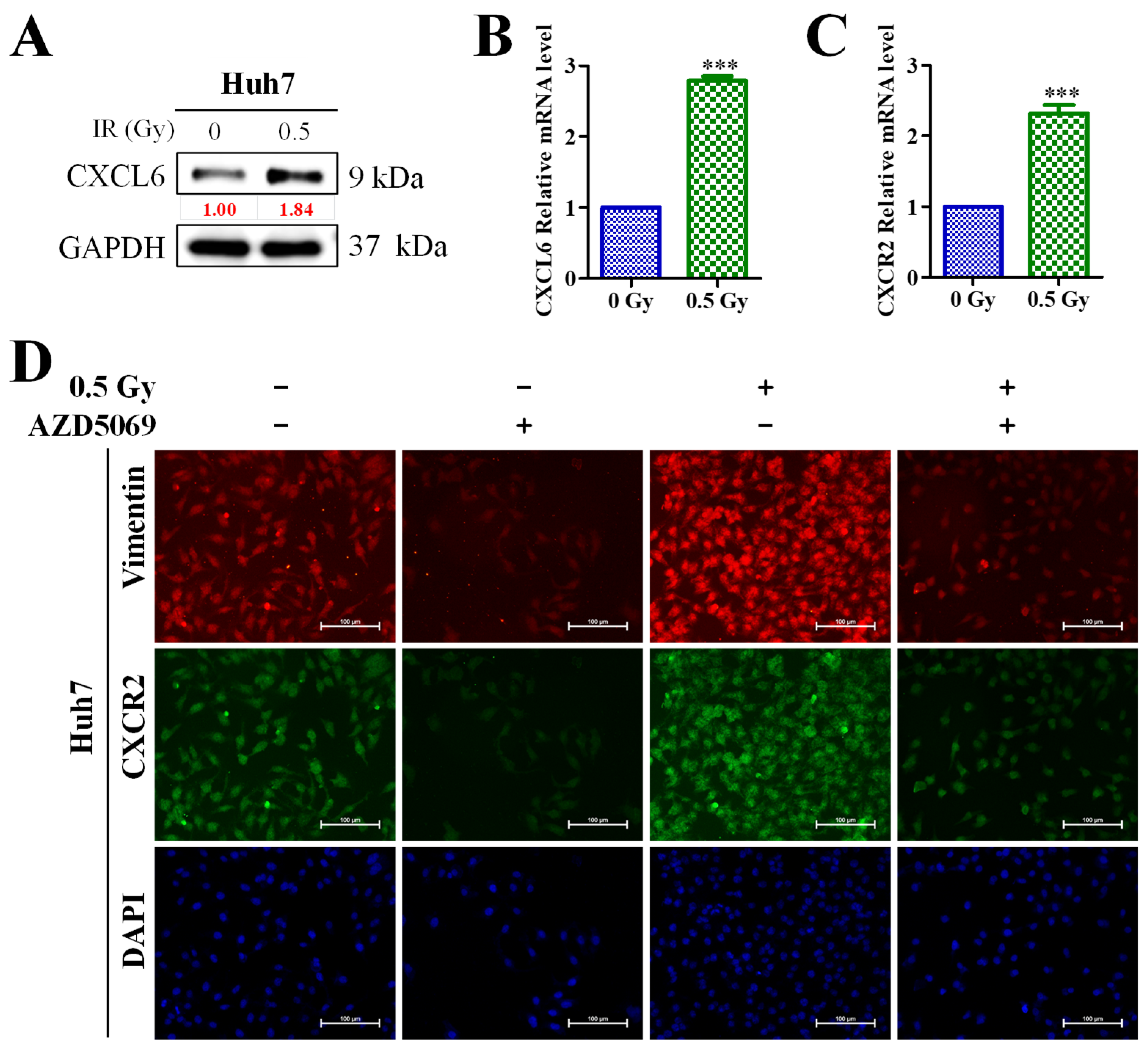
3.5. Enhanced CXCL6/CXCR2 Signaling Was Associated with Low-Dose (0.5-Gy) Radiation-Induced Epithelial–Mesenchymal Transition (EMT) of HCC Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baggiolini, M. Chemokines in pathology and medicine. J. Intern. Med. 2001, 250, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Bule, P.; Aguiar, S.I.; Aires-Da-Silva, F.; Dias, J.N.R. Chemokine-directed tumor microenvironment modulation in cancer immunotherapy. Int. J. Mol. Sci. 2021, 22, 9804. [Google Scholar] [CrossRef]
- Marcuzzi, E.; Angioni, R.; Molon, B.; Cali, B. Chemokines and chemokine receptors: Orchestrating tumor metastasization. Int. J. Mol. Sci. 2018, 20, 96. [Google Scholar] [CrossRef] [Green Version]
- Nagarsheth, N.; Wicha, M.S.; Zou, W. Chemokines in the cancer microenvironment and their relevance in cancer immunotherapy. Nat. Rev. Immunol. 2017, 17, 559–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cendrowicz, E.; Sas, Z.; Bremer, E.; Rygiel, T.P. The role of macrophages in cancer development and therapy. Cancers 2021, 13, 1946. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Xu, J.; Lan, H. Tumor-associated macrophages in tumor metastasis: Biological roles and clinical therapeutic applications. J. Hematol. Oncol. 2019, 12, 76. [Google Scholar] [CrossRef]
- Ruytinx, P.; Proost, P.; Van Damme, J.; Struyf, S. Chemokine-induced macrophage polarization in inflammatory conditions. Front. Immunol. 2018, 9, 1930. [Google Scholar] [CrossRef] [Green Version]
- Susek, K.H.; Karvouni, M.; Alici, E.; Lundqvist, A. The role of cxc chemokine receptors 1–4 on immune cells in the tumor microenvironment. Front. Immunol. 2018, 9, 2159. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Ma, X.-L.; Wei, Y.-Q.; Wei, X.-W. Potential roles and targeted therapy of the cxcls/cxcr2 axis in cancer and inflammatory diseases. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2019, 1871, 289–312. [Google Scholar] [CrossRef]
- Liu, G.; An, L.; Zhang, H.; Du, P.; Sheng, Y. Activation of cxcl6/cxcr1/2 axis promotes the growth and metastasis of osteosarcoma cells in vitro and in vivo. Front. Pharmacol. 2019, 10, 307. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Shen, J.; Man, K.; Chu, E.S.; Yau, T.O.; Sung, J.C.; Go, M.Y.; Deng, J.; Lu, L.; Wong, V.W.; et al. Cxcl10 plays a key role as an inflammatory mediator and a non-invasive biomarker of non-alcoholic steatohepatitis. J. Hepatol. 2014, 61, 1365–1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Naour, A.; Prat, M.; Thibault, B.; Mevel, R.; Lemaitre, L.; Leray, H.; Joubert, M.V.; Coulson, K.; Golzio, M.; Lefevre, L.; et al. Tumor cells educate mesenchymal stromal cells to release chemoprotective and immunomodulatory factors. J. Mol. Cell Biol. 2020, 12, 202–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mas, V.R.; Maluf, D.G.; Archer, K.J.; Yanek, K.; Kong, X.; Kulik, L.; Freise, C.E.; Olthoff, K.M.; Ghobrial, R.M.; Mclver, P. Genes involved in viral carcinogenesis and tumor initiation in hepatitis c virus-induced hepatocellular carcinoma. Mol. Med. 2009, 15, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, S.; Guyot, M.; Grépin, R.; Pagès, G. The elr(+)cxcl chemokines and their receptors cxcr1/cxcr2: A signaling axis and new target for the treatment of renal cell carcinoma. Oncoimmunology 2014, 3, e28399. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Ge, K.; Zhou, W.; Lu, J.; Jia, C.; Zhu, H. C-x-c chemokine receptor 2 (cxcr2) promotes hepatocellular carcinoma immune evasion via regulating programmed death-ligand 1 (pd-l1). Biol. Chem. 2021, 402, 729–737. [Google Scholar] [CrossRef]
- Linge, H.M.; Collin, M.; Nordenfelt, P.; Mörgelin, M.; Malmsten, M.; Egesten, A. The human cxc chemokine granulocyte chemotactic protein 2 (gcp-2)/cxcl6 possesses membrane-disrupting properties and is antibacterial. Antimicrob. Agents Chemother. 2008, 52, 2599–2607. [Google Scholar] [CrossRef] [Green Version]
- Wasmuth, H.E.; Lammert, F.; Zaldivar, M.M.; Weiskirchen, R.; Hellerbrand, C.; Scholten, D.; Berres, M.-L.; Zimmermann, H.; Streetz, K.L.; Tacke, F.J.G. Antifibrotic effects of cxcl9 and its receptor cxcr3 in livers of mice and humans. Gastroenterology 2009, 137, 309–319.E3. [Google Scholar] [CrossRef] [Green Version]
- Sadik, C.D.; Kim, N.D.; Luster, A.D. Neutrophils cascading their way to inflammation. Trends Immunol. 2011, 32, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Kebschull, M.; Demmer, R.; Behle, J.H.; Pollreisz, A.; Heidemann, J.; Belusko, P.B.; Celenti, R.; Pavlidis, P.; Papapanou, P.N. Granulocyte chemotactic protein 2 (gcp-2/cxcl6) complements interleukin-8 in periodontal disease. J. Periodontal Res. 2009, 44, 465–471. [Google Scholar] [CrossRef] [Green Version]
- Besnard, A.G.; Struyf, S.; Guabiraba, R.; Fauconnier, L.; Rouxel, N.; Proost, P.; Uyttenhove, C.; Van Snick, J.; Couillin, I.; Ryffel, B. Cxcl6 antibody neutralization prevents lung inflammation and fibrosis in mice in the bleomycin model. J. Leukoc. Biol. 2013, 94, 1317–1323. [Google Scholar] [CrossRef]
- Cai, X.; Li, Z.; Zhang, Q.; Qu, Y.; Xu, M.; Wan, X.; Lu, L. Cxcl6-egfr-induced kupffer cells secrete tgf-β1 promoting hepatic stellate cell activation via the smad2/brd4/c-myc/ezh2 pathway in liver fibrosis. J. Cell. Mol. Med. 2018, 22, 5050–5061. [Google Scholar] [CrossRef] [Green Version]
- Farha, M.; Jairath, N.K.; Lawrence, T.S.; El Naqa, I. Characterization of the tumor immune microenvironment identifies m0 macrophage-enriched cluster as a poor prognostic factor in hepatocellular carcinoma. JCO Clin. Cancer Inf. 2020, 4, 1002–1013. [Google Scholar] [CrossRef]
- Proost, P.; De Wolf-Peeters, C.; Conings, R.; Opdenakker, G.; Billiau, A.; Van Damme, J. Identification of a novel granulocyte chemotactic protein (gcp-2) from human tumor cells. In vitro and in vivo comparison with natural forms of gro, ip-10, and il-8. J. Immunol. 1993, 150, 1000–1010. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.; Delgado, M.B.; Jones, S.A.; Dewald, B.; Clark-Lewis, I.; Baggiolini, M. Granulocyte chemotactic protein 2 acts via both il-8 receptors, cxcr1 and cxcr2. Eur. J. Immunol. 1998, 28, 164–170. [Google Scholar] [CrossRef]
- Cullberg, M.; Arfvidsson, C.; Larsson, B.; Malmgren, A.; Mitchell, P.; Wahlby Hamren, U.; Wray, H. Pharmacokinetics of the oral selective cxcr2 antagonist azd5069: A summary of eight phase I studies in healthy volunteers. Drugs R&D 2018, 18, 149–159. [Google Scholar]
- Kirsten, A.M.; Forster, K.; Radeczky, E.; Linnhoff, A.; Balint, B.; Watz, H.; Wray, H.; Salkeld, L.; Cullberg, M.; Larsson, B. The safety and tolerability of oral azd5069, a selective cxcr2 antagonist, in patients with moderate-to-severe copd. Pulm. Pharmacol. Ther. 2015, 31, 36–41. [Google Scholar] [CrossRef]
- Nicholls, D.J.; Wiley, K.; Dainty, I.; MacIntosh, F.; Phillips, C.; Gaw, A.; Mardh, C.K. Pharmacological characterization of azd5069, a slowly reversible cxc chemokine receptor 2 antagonist. J. Pharmacol. Exp. Ther. 2015, 353, 340–350. [Google Scholar] [CrossRef] [Green Version]
- Bruni, D.; Angell, H.K.; Galon, J. The immune contexture and immunoscore in cancer prognosis and therapeutic efficacy. Nat. Rev. Cancer 2020, 20, 662–680. [Google Scholar] [CrossRef]
- Chavez-Galan, L.; Olleros, M.L.; Vesin, D.; Garcia, I. Much more than m1 and m2 macrophages, there are also cd169(+) and tcr(+) macrophages. Front. Immunol. 2015, 6, 263. [Google Scholar]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.C.; Baskaran, R.; Lee, N.H.; Hsu, H.H.; Ho, T.J.; Tu, C.C.; Lin, Y.M.; Viswanadha, V.P.; Kuo, W.W.; Huang, C.Y. Cxcl2/cxcr2 axis induces cancer stem cell characteristics in cpt-11-resistant lovo colon cancer cells via galphai-2 and galphaq/11. J. Cell Physiol. 2019, 234, 11822–11834. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-L.; Tsai, Y.-C.; Pikatan, N.W.; Yeh, C.-T.; Yadav, V.K.; Chen, M.-Y.; Tsai, J.-T. Tumor-Associated Macrophages Affect the Tumor Microenvironment and Radioresistance via the Upregulation of CXCL6/CXCR2 in Hepatocellular Carcinoma. Biomedicines 2023, 11, 2081. https://doi.org/10.3390/biomedicines11072081
Lee H-L, Tsai Y-C, Pikatan NW, Yeh C-T, Yadav VK, Chen M-Y, Tsai J-T. Tumor-Associated Macrophages Affect the Tumor Microenvironment and Radioresistance via the Upregulation of CXCL6/CXCR2 in Hepatocellular Carcinoma. Biomedicines. 2023; 11(7):2081. https://doi.org/10.3390/biomedicines11072081
Chicago/Turabian StyleLee, Hsin-Lun, Yi-Chieh Tsai, Narpati Wesa Pikatan, Chi-Tai Yeh, Vijesh Kumar Yadav, Ming-Yao Chen, and Jo-Ting Tsai. 2023. "Tumor-Associated Macrophages Affect the Tumor Microenvironment and Radioresistance via the Upregulation of CXCL6/CXCR2 in Hepatocellular Carcinoma" Biomedicines 11, no. 7: 2081. https://doi.org/10.3390/biomedicines11072081
APA StyleLee, H. -L., Tsai, Y. -C., Pikatan, N. W., Yeh, C. -T., Yadav, V. K., Chen, M. -Y., & Tsai, J. -T. (2023). Tumor-Associated Macrophages Affect the Tumor Microenvironment and Radioresistance via the Upregulation of CXCL6/CXCR2 in Hepatocellular Carcinoma. Biomedicines, 11(7), 2081. https://doi.org/10.3390/biomedicines11072081