


Development of New Models of Oral Mucosa to Investigate the Impact of the Structure of Transmembrane Mucin-1 on the Mucosal Pellicle Formation and Its Physicochemical Properties
 , , , ,
, , , ,  , , , , and
, , , , and 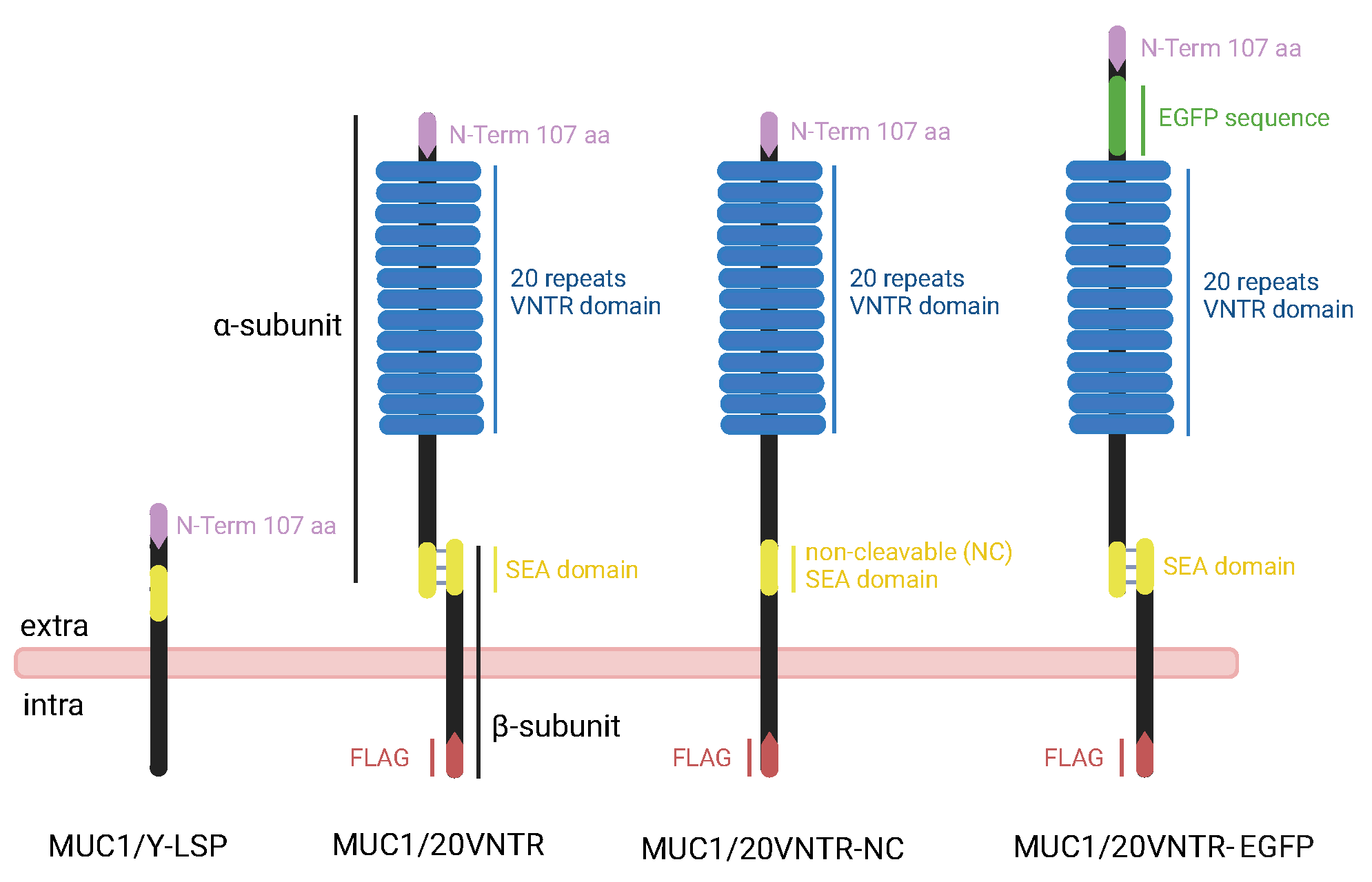{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. MUC1 Gene Design and Construction
2.2. Preparation of the Stable MUC1 TR146 Cell Lines
2.3. Saliva Collection and Application
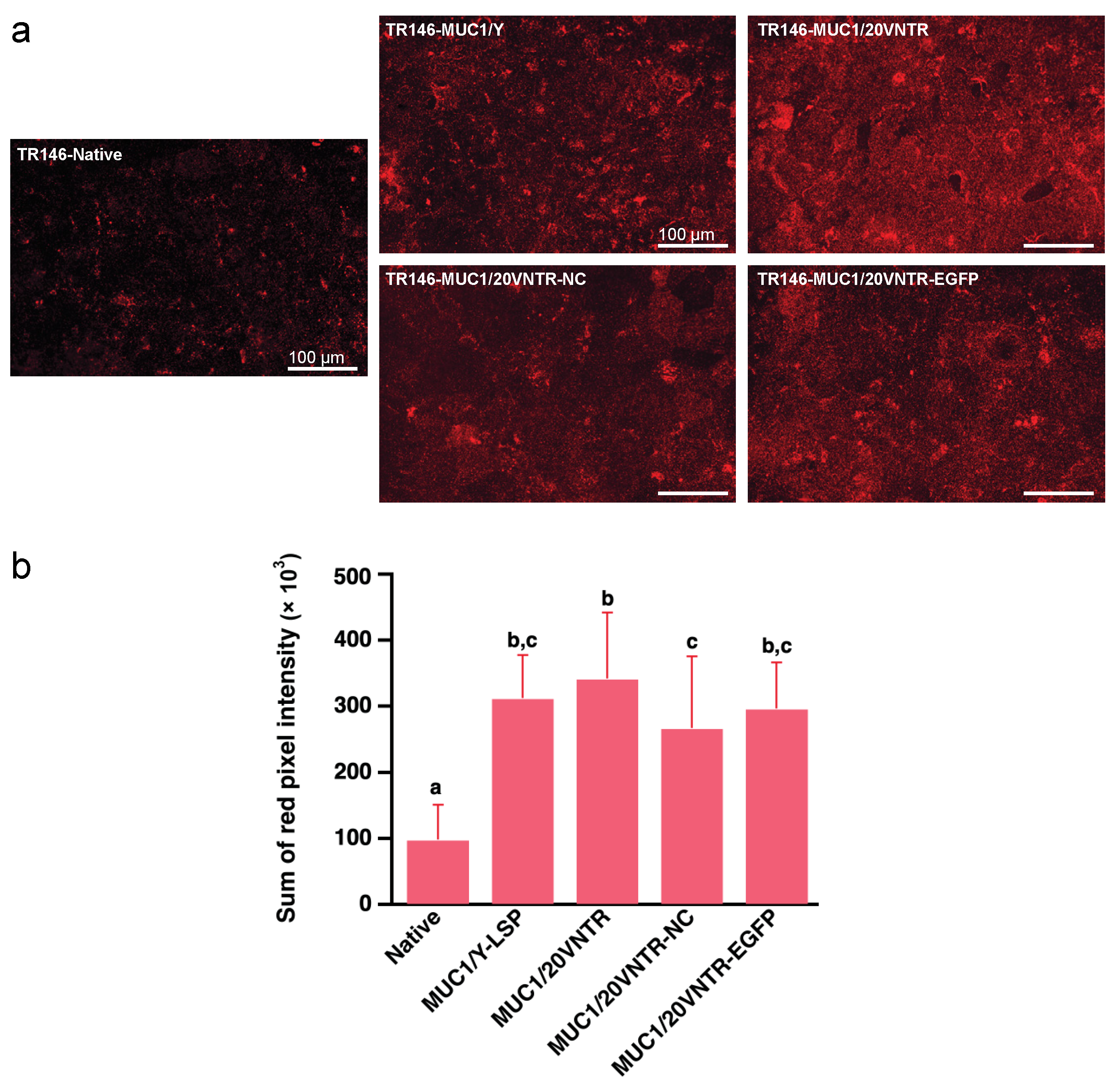
2.4. Fluorescent Immunostaining of MUC1, Plasma Membrane, and MUC5B
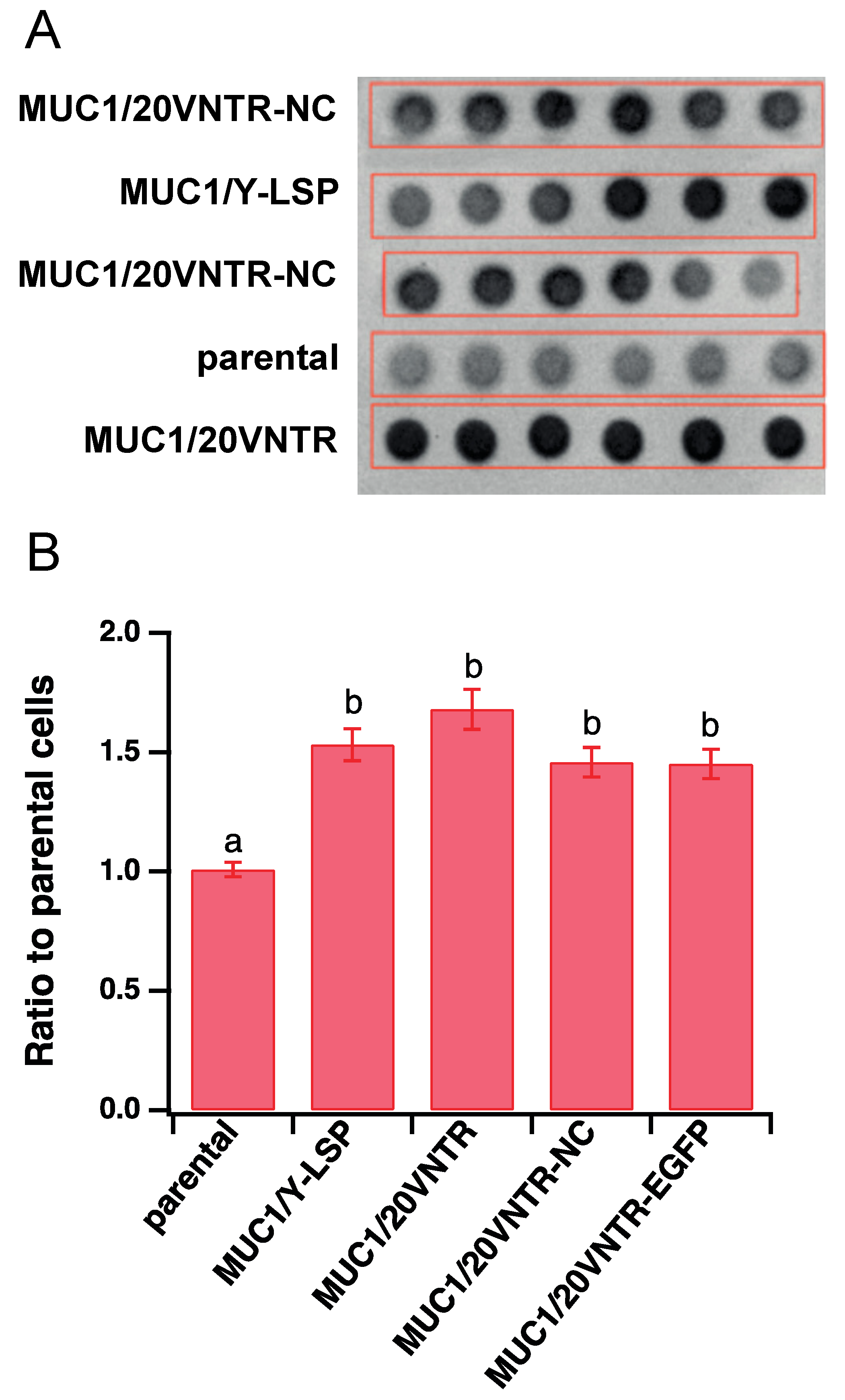
2.5. Semi-Quantification of the MUC1 via Dot Blot Analyses
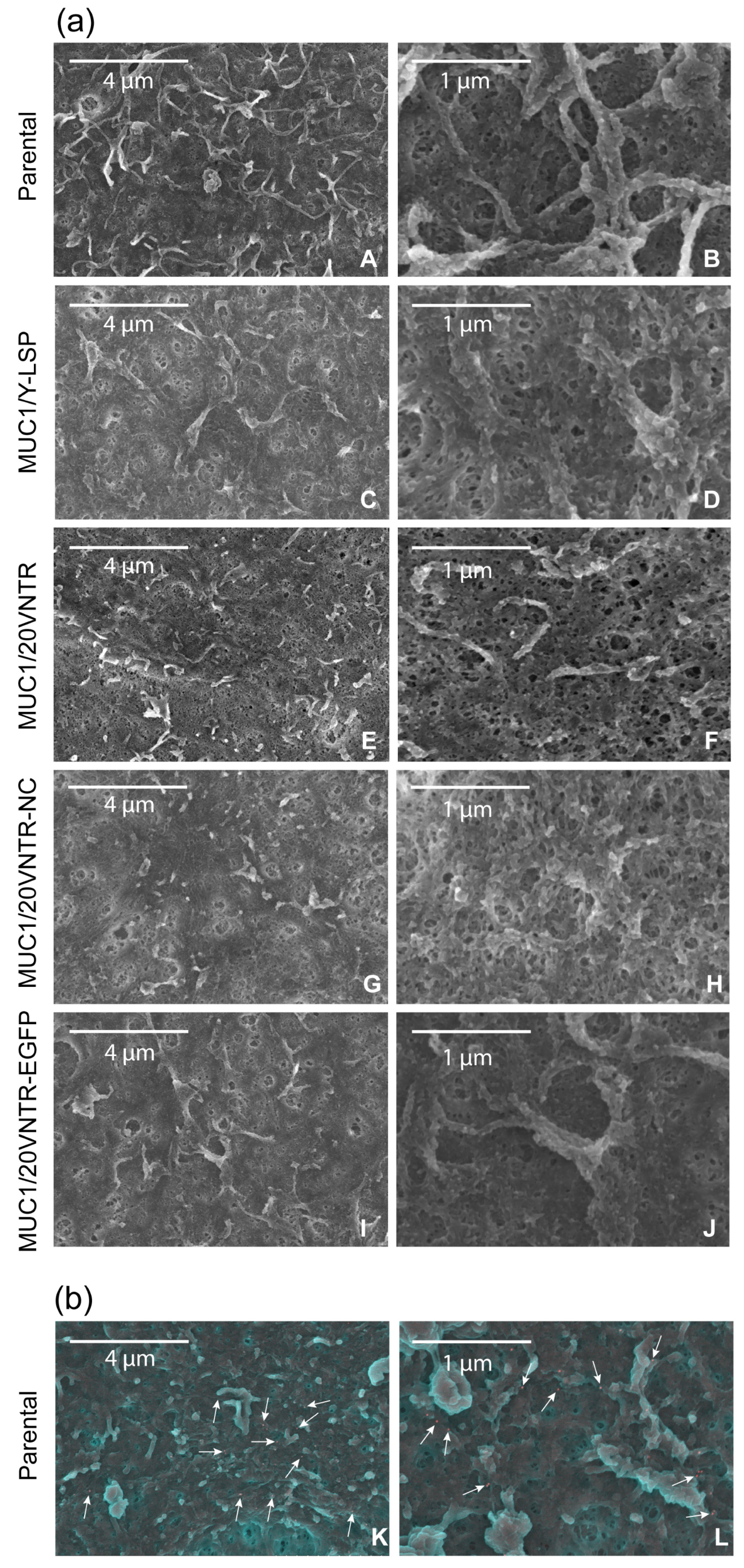
2.6. Immuno-Scanning Electron Microscopy (SEM) and Atomic Force Microscopy-Based Infrared Spectroscopy (AFM-IR)
3. Results
3.1. Expression and Localization of MUC1
3.1.1. Expression of MUC1 in Stably Transfected TR146 Cells
3.1.2. Cellular Localization of MUC1
3.2. Mucosal Pellicle (MP) Formation: Binding of the Salivary Protein MUC5B
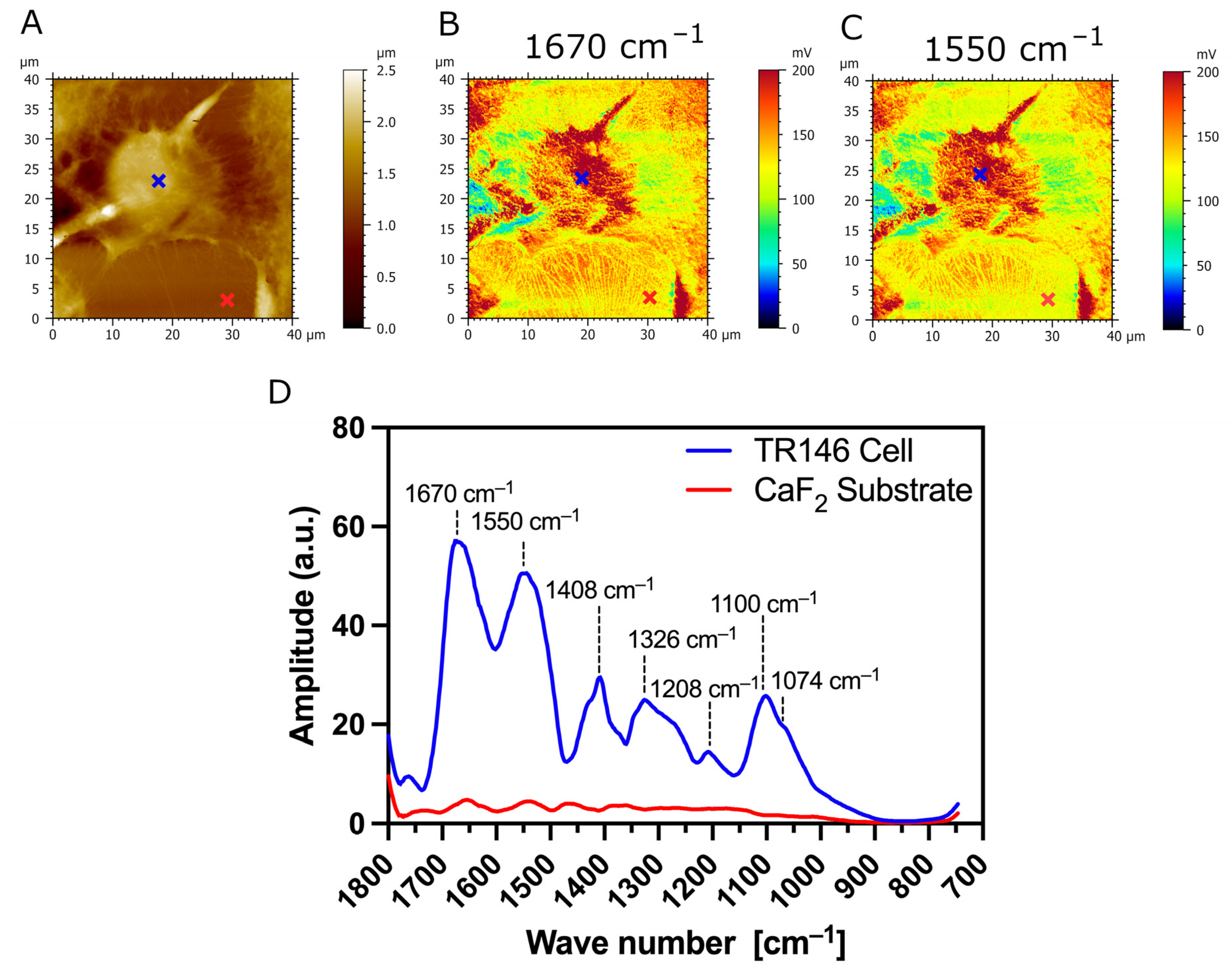
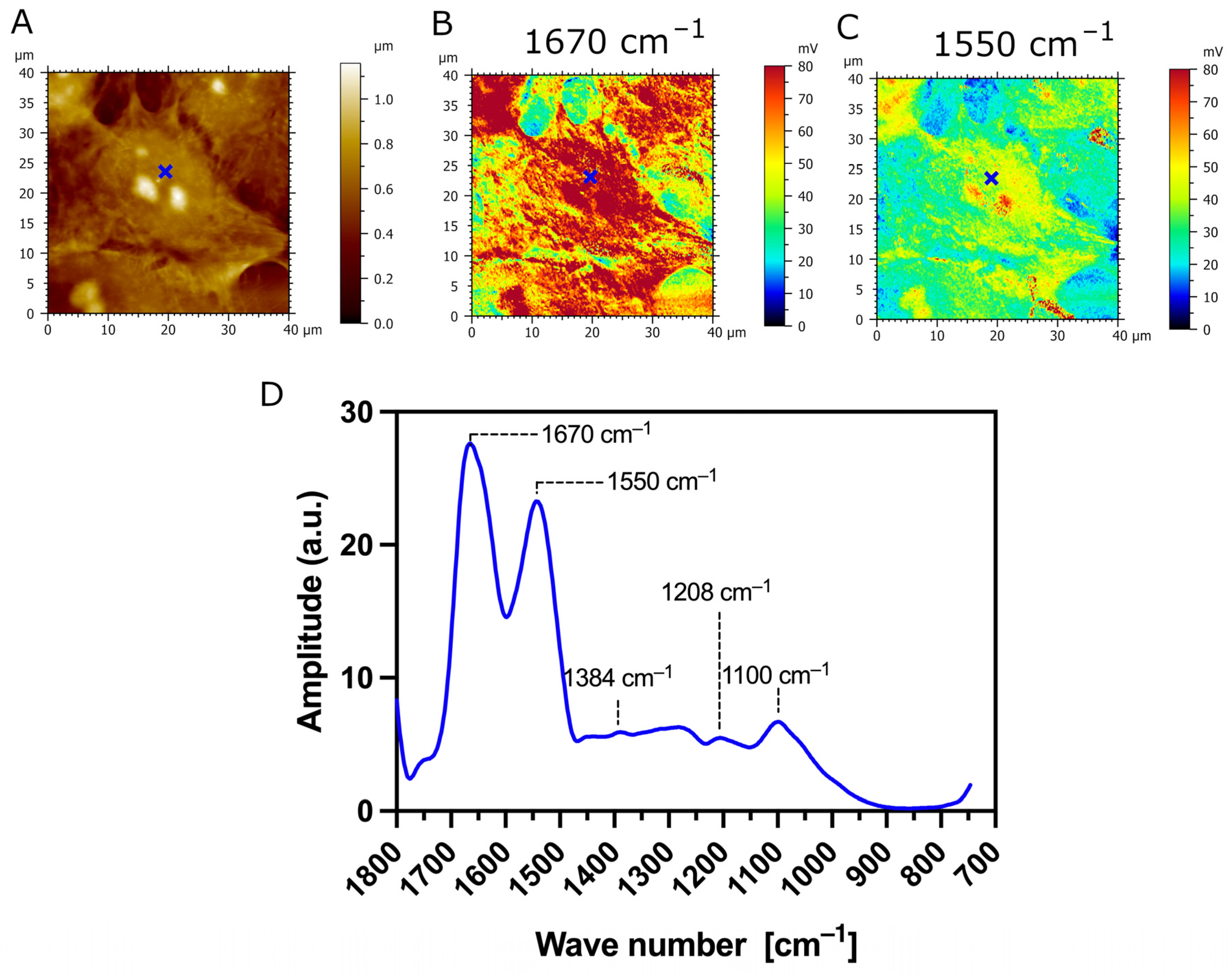
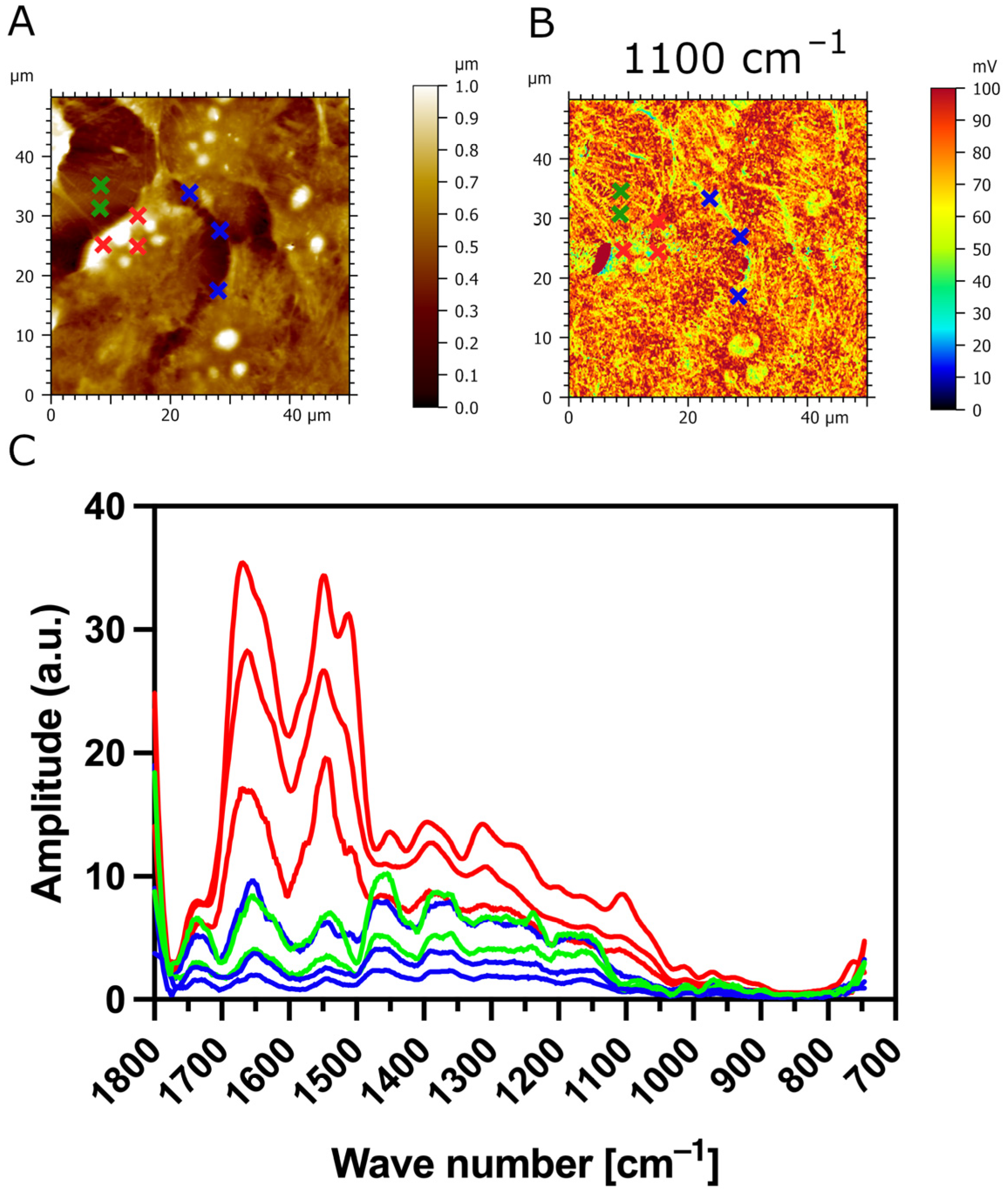
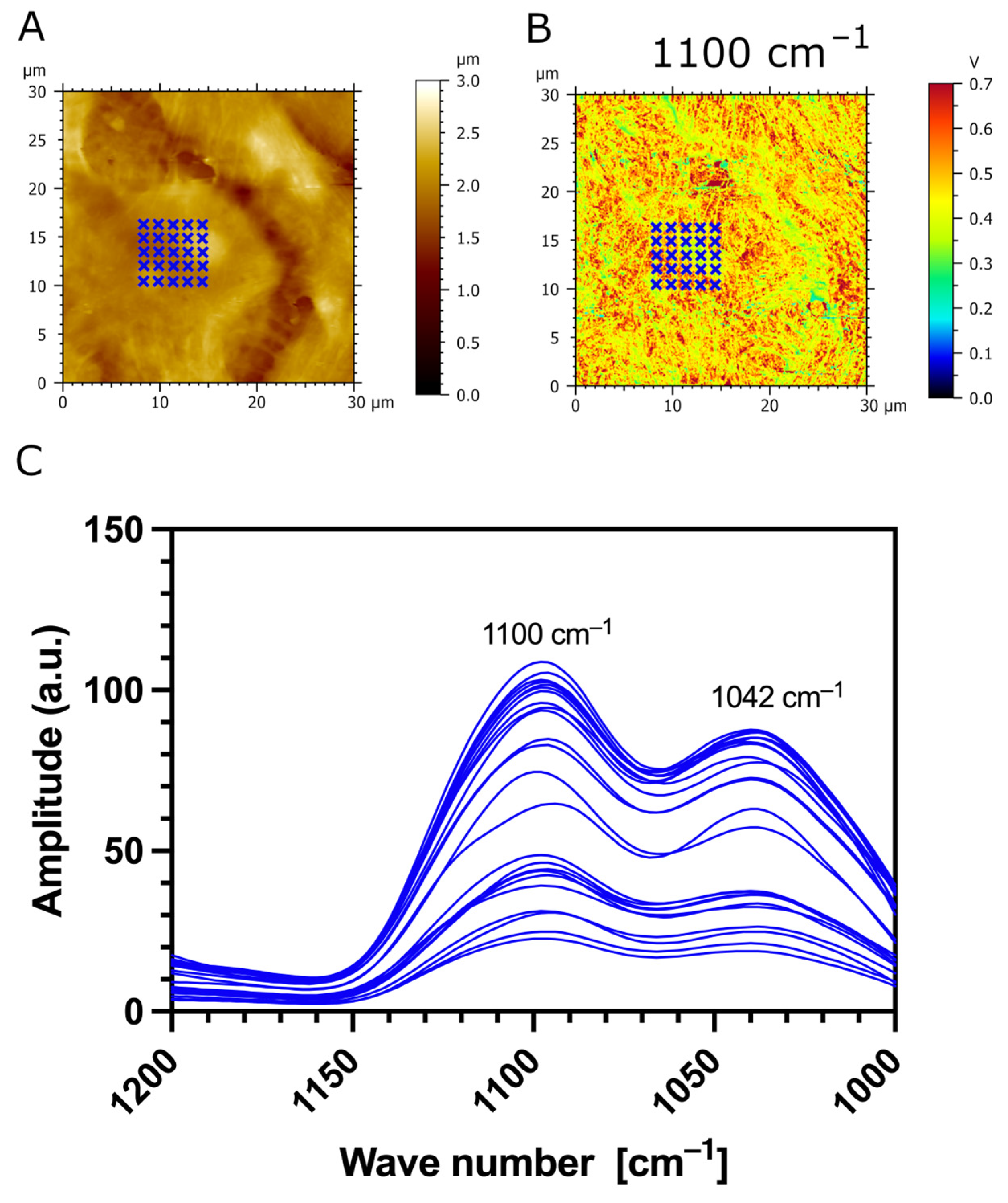
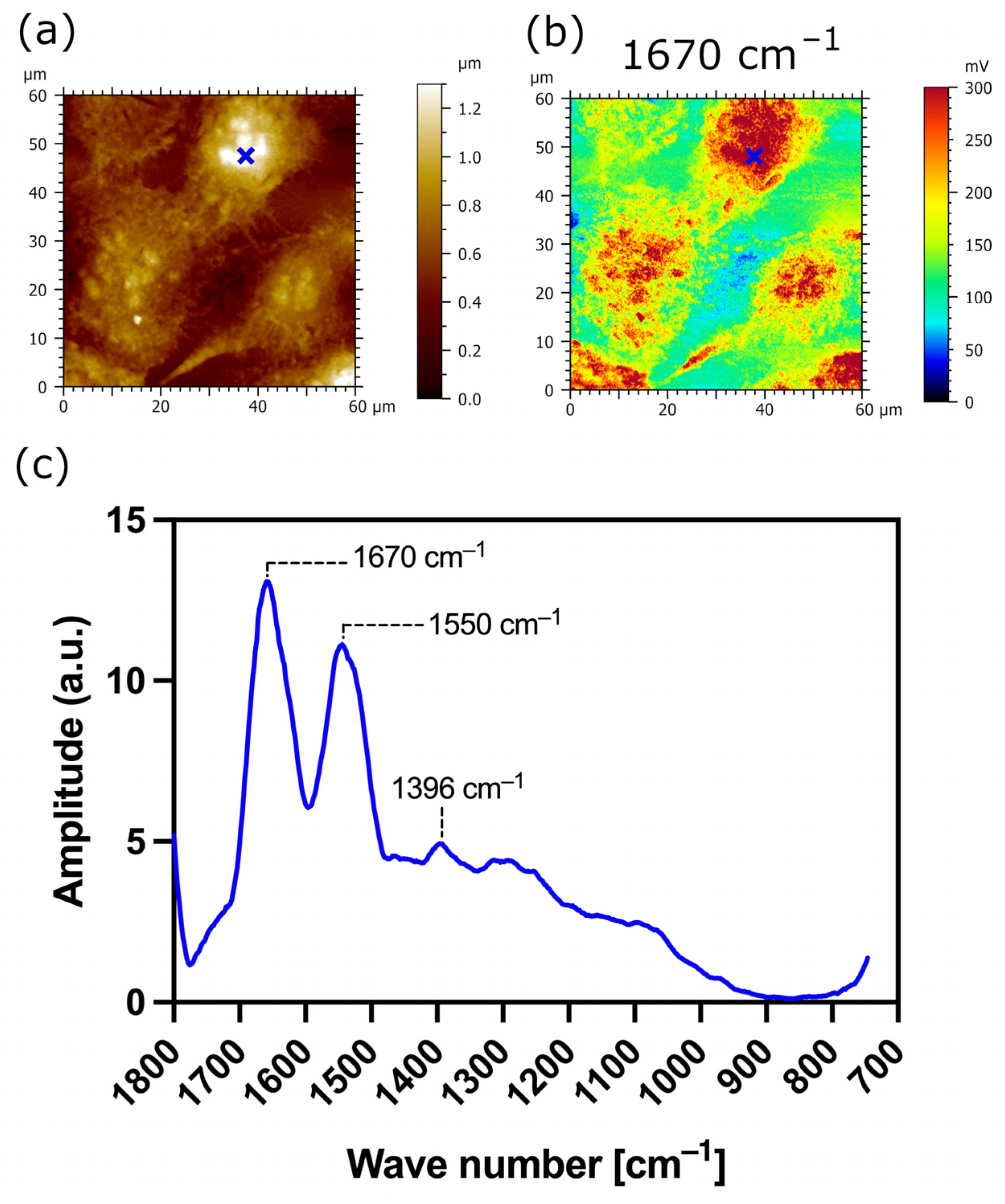
3.3. AFM-IR Nanospectroscopy Analysis of TR146 Buccal Cells: Impact of MUC1 Isoform on Chemical Properties of Cell Surfaces
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vielmuth, F. Anatomy of the Oral Mucosa. In Diseases of the Oral Mucosa: Study Guide and Review; Schmidt, E., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 5–19. [Google Scholar]
- Roed-Petersen, B.; Renstrup, G. A Topographical Classification of the Oral Mucosa Suitable for Electronic Data Processing Its Application to 560 Leukoplakias. Acta Odontol. Scand. 1969, 27, 681–695. [Google Scholar] [CrossRef]
- Canon, F.; Belloir, C.; Bourillot, E.; Brignot, H.; Briand, L.; Feron, G.; Lesniewska, E.; Nivet, C.; Septier, C.; Schwartz, M.; et al. Perspectives on Astringency Sensation: An Alternative Hypothesis on the Molecular Origin of Astringency. J. Agric. Food Chem. 2021, 69, 3822–3826. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.; Carpenter, G.H. A physiological model of tea-induced astringency. Physiol. Behav. 2008, 95, 290–294. [Google Scholar] [CrossRef] [PubMed]
- Ployon, S.; Morzel, M.; Belloir, C.; Bonnotte, A.; Bourillot, E.; Briand, L.; Lesniewska, E.; Lherminier, J.; Aybeke, E.; Canon, F. Mechanisms of astringency: Structural alteration of the oral mucosal pellicle by dietary tannins and protective effect of bPRPs. Food Chem. 2018, 253, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Ployon, S.; Brulé, M.; Andriot, I.; Morzel, M.; Canon, F. Understanding retention and metabolization of aroma compounds using an in vitro model of oral mucosa. Food Chem. 2020, 318, 126468. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-González, C.; Brule, M.; Martin, C.; Feron, G.; Canon, F. Molecular mechanisms of aroma persistence: From noncovalent interactions between aroma compounds and the oral mucosa to metabolization of aroma compounds by saliva and oral cells. Food Chem. 2022, 373, 131467. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.M.C.; Dawes, C. The Surface Area of the Adult Human Mouth and Thickness of the Salivary Film Covering the Teeth and Oral Mucosa. J. Dent. Res. 1987, 66, 1300–1302. [Google Scholar] [CrossRef]
- Pramanik, R.; Osailan, S.M.; Challacombe, S.J.; Urquhart, D.; Proctor, G.B. Protein and mucin retention on oral mucosal surfaces in dry mouth patients. Eur. J. Oral Sci. 2010, 118, 245–253. [Google Scholar] [CrossRef]
- Assy, Z.; Jager, D.H.J.; Brand, H.S.; Bikker, F.J. Salivary film thickness and MUC5B levels at various intra-oral surfaces. Clin. Oral Investig. 2023, 27, 859–869. [Google Scholar] [CrossRef]
- Cárdenas, M.; Elofsson, U.; Lindh, L. Salivary Mucin MUC5B Could Be an Important Component of in Vitro Pellicles of Human Saliva: An In Situ Ellipsometry and Atomic Force Microscopy Study. Biomacromolecules 2007, 8, 1149–1156. [Google Scholar] [CrossRef]
- Faruque, M.; Wanschers, M.; Ligtenberg, A.J.; Laine, M.L.; Bikker, F.J. A review on the role of salivary MUC5B in oral health. J. Oral Biosci. 2022, 64, 392–399. [Google Scholar] [CrossRef]
- Gibbins, H.; Proctor, G.; Yakubov, G.; Wilson, S.; Carpenter, G. Concentration of salivary protective proteins within the bound oral mucosal pellicle. Oral Dis. 2013, 20, 707–713. [Google Scholar] [CrossRef]
- Gibbins, H.L.; Yakubov, G.E.; Proctor, G.B.; Wilson, S.; Carpenter, G.H. What interactions drive the salivary mucosal pellicle formation? Colloids Surf. B Biointerfaces 2014, 120, 184–192. [Google Scholar] [CrossRef]
- Schwartz, M.; Brignot, H.; Feron, G.; Hummel, T.; Zhu, Y.; von Koskull, D.; Heydel, J.-M.; Lirussi, F.; Canon, F.; Neiers, F. Role of human salivary enzymes in bitter taste perception. Food Chem. 2022, 386, 132798. [Google Scholar] [CrossRef] [PubMed]
- Bradway, S.D.; Bergey, E.J.; Scannapieco, F.A.; Ramasubbu, N.; Zawacki, S.; Levine, M.J. Formation of salivary-mucosal pellicle: The role of transglutaminase. Biochem. J. 1992, 284, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Biesbrock, A.R.; Reddy, M.S.; Levine, M.J. Interaction of a salivary mucin-secretory immunoglobulin A complex with mucosal pathogens. Infect. Immun. 1991, 59, 3492–3497. [Google Scholar] [CrossRef] [PubMed]
- Gibbins, H.L.; Proctor, G.B.; Yakubov, G.E.; Wilson, S.; Carpenter, G.H. SIgA Binding to Mucosal Surfaces Is Mediated by Mucin-Mucin Interactions. PLoS ONE 2015, 10, e0119677. [Google Scholar] [CrossRef] [PubMed]
- Aybeke, E.N.; Ployon, S.; Brulé, M.; De Fonseca, B.; Bourillot, E.; Morzel, M.; Lesniewska, E.; Canon, F. Nanoscale Mapping of the Physical Surface Properties of Human Buccal Cells and Changes Induced by Saliva. Langmuir 2019, 35, 12647–12655. [Google Scholar] [CrossRef] [PubMed]
- Ployon, S.; Belloir, C.; Bonnotte, A.; Lherminier, J.; Canon, F.; Morzel, M. The membrane-associated MUC1 improves adhesion of salivary MUC5B on buccal cells. Application to development of an in vitro cellular model of oral epithelium. Arch. Oral Biol. 2016, 61, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Offner, G.D.; Troxler, R.F. Heterogeneity of High-molecular-weight Human Salivary Mucins. Adv. Dent. Res. 2000, 14, 69–75. [Google Scholar] [CrossRef]
- Ukkonen, H.; Pirhonen, P.; Herrala, M.; Mikkonen, J.J.W.; Singh, S.P.; Sormunen, R.; Kullaa, A.M. Oral mucosal epithelial cells express the membrane anchored mucin MUC1. Arch. Oral Biol. 2017, 73, 269–273. [Google Scholar] [CrossRef]
- Levitin, F.; Stern, O.; Weiss, M.; Gil-Henn, C.; Ziv, R.; Prokocimer, Z.; Smorodinsky, N.I.; Rubinstein, D.B.; Wreschner, D.H. The MUC1 SEA Module Is a Self-cleaving Domain. J. Biol. Chem. 2005, 280, 33374–33386. [Google Scholar] [CrossRef]
- Macao, B.; Johansson, D.G.A.; Hansson, G.C.; Härd, T. Autoproteolysis coupled to protein folding in the SEA domain of the membrane-bound MUC1 mucin. Nat. Struct. Mol. Biol. 2006, 13, 71. [Google Scholar] [CrossRef] [PubMed]
- Parry, S.; Silverman, H.S.; McDermott, K.; Willis, A.; Hollingsworth, M.A.; Harris, A. Identification of MUC1 Proteolytic Cleavage Sites In Vivo. Biochem. Biophys. Res. Commun. 2001, 283, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, F.-G.; Müller, S. MUC1: The polymorphic appearance of a human mucin. Glycobiology 2000, 10, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Muller, S.; Alving, K.; Peter-Katalinic, J.; Zachara, N.; Gooley, A.A.; Hanisch, F.G. High density O-glycosylation on tandem repeat peptide from secretory MUC1 of T47D breast cancer cells. J. Biol. Chem. 1999, 274, 18165–18172. [Google Scholar] [CrossRef] [PubMed]
- Imbert, Y.; Darling, D.S.; Jumblatt, M.M.; Foulks, G.N.; Couzin, E.G.; Steele, P.S.; Young, W.W. MUC1 splice variants in human ocular surface tissues: Possible differences between dry eye patients and normal controls. Exp. Eye Res. 2006, 83, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Rupniak, H.T.; Rowlatt, C.; Lane, E.B.; Steele, J.G.; Trejdosiewicz, L.K.; Laskiewicz, B.; Povey, S.; Hill, B.T. Characteristics of four new human cell lines derived from squamous cell carcinomas of the head and neck. J. Natl. Cancer Inst. 1985, 75, 621–635. [Google Scholar] [PubMed]
- Veerman, E.C.I.; Bolscher, J.G.M.; Appelmelk, B.J.; Bloemena, E.; van den Berg, T.K.; Amerongen, A.V.N. A monoclonal antibody directed against high Mr salivary mucins recognizes the SO3-3Galβ1-3GlcNAc moiety of sulfo-Lewisa: A histochemical survey of human and rat tissue. Glycobiology 1997, 7, 37–43. [Google Scholar] [CrossRef]
- Mathurin, J.; Deniset-Besseau, A.; Bazin, D.; Dartois, E.; Wagner, M.; Dazzi, A. Photothermal AFM-IR spectroscopy and imaging: Status, challenges, and trends. J. Appl. Phys. 2022, 131, 010901. [Google Scholar] [CrossRef]
- Dazzi, A.; Glotin, F.; Carminati, R. Theory of infrared nanospectroscopy by photothermal induced resonance. J. Appl. Phys. 2010, 107, 124519. [Google Scholar] [CrossRef]
- Dazzi, A.; Prater, C.B. AFM-IR: Technology and Applications in Nanoscale Infrared Spectroscopy and Chemical Imaging. Chem. Rev. 2017, 117, 5146–5173. [Google Scholar] [CrossRef] [PubMed]
- Custovic, I.; Pocholle, N.; Bourillot, E.; Lesniewska, E.; Piétrement, O. Infrared nanospectroscopic imaging of DNA molecules on mica surface. Sci. Rep. 2022, 12, 18972. [Google Scholar] [CrossRef] [PubMed]
- Maryjose, N.; Custovic, I.; Chaabane, L.; Lesniewska, E.; Piétrement, O.; Chambin, O.; Assifaoui, A. Core-shell polygalacturonate magnetic iron oxide nanoparticles: Synthesis, characterization, and functionalities. Int. J. Biol. Macromol. 2022, 220, 360–370. [Google Scholar] [CrossRef]
- Ruggeri, F.S.; Habchi, J.; Chia, S.; Horne, R.I.; Vendruscolo, M.; Knowles, T.P.J. Infrared nanospectroscopy reveals the molecular interaction fingerprint of an aggregation inhibitor with single Aβ42 oligomers. Nat. Commun. 2021, 12, 688. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Yang, X.; Ji, Z.; Zhu, L.; Ma, N.; Chen, D.; Jia, X.; Tang, J.; Cao, Y. DFT-Calculated IR Spectrum Amide I, II, and III Band Contributions of N-Methylacetamide Fine Components. ACS Omega 2020, 5, 8572–8578. [Google Scholar] [CrossRef] [PubMed]
- Movasaghi, Z.; Rehman, S.; ur Rehman, D.I. Fourier Transform Infrared (FTIR) Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2008, 43, 134–179. [Google Scholar] [CrossRef]
- Durak, R.; Ciak, B.; Durak, T. Highly Efficient Use of Infrared Spectroscopy (ATR-FTIR) to Identify Aphid Species. Biology 2022, 11, 1232. [Google Scholar] [CrossRef]
- Fujioka, N.; Morimoto, Y.; Arai, T.; Kikuchi, M. Discrimination between normal and malignant human gastric tissues by Fourier transform infrared spectroscopy. Cancer Detect. Prev. 2004, 28, 32–36. [Google Scholar] [CrossRef]
- Barth, A. Infrared spectroscopy of proteins. Biochim. Biophys. Acta (BBA)—Bioenerg. 2007, 1767, 1073–1101. [Google Scholar] [CrossRef]
- Paluszkiewicz, C.; Kwiatek, W.M. Analysis of human cancer prostate tissues using FTIR microspectroscopy and SRIXE techniques. J. Mol. Struct. 2001, 565–566, 329–334. [Google Scholar] [CrossRef]
- Barchi, J.J.; Strain, C.N. The effect of a methyl group on structure and function: Serine vs. threonine glycosylation and phosphorylation. Front. Mol. Biosci. 2023, 10, 1117850. [Google Scholar] [CrossRef] [PubMed]
- Barchi Jr, J.J. Mucin-Type Glycopeptide Structure in Solution: Past, Present, and Future. Biopolymers 2013, 99, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Naganagowda, G.A.; Gururaja, T.E.L.; Satyanarayana, J.; Levine, M.J. NMR analysis of human salivary mucin (MUC7) derived O-linked model glycopeptides: Comparison of structural features and carbohydrate–peptide interactions. J. Pept. Res. 1999, 54, 290–310. [Google Scholar] [CrossRef]
- Derenne, A.; Derfoufi, K.-M.; Cowper, B.; Delporte, C.; Goormaghtigh, E. FTIR spectroscopy as an analytical tool to compare glycosylation in therapeutic monoclonal antibodies. Anal. Chim. Acta 2020, 1112, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Khajehpour, M.; Dashnau, J.L.; Vanderkooi, J.M. Infrared spectroscopy used to evaluate glycosylation of proteins. Anal. Biochem. 2006, 348, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Asikainen, P.; Ruotsalainen, T.J.; Mikkonen, J.J.W.; Koistinen, A.; ten Bruggenkate, C.; Kullaa, A.M. The defence architecture of the superficial cells of the oral mucosa. Med. Hypotheses 2012, 78, 790–792. [Google Scholar] [CrossRef]
- Demouveaux, B.; Gouyer, V.; Gottrand, F.; Narita, T.; Desseyn, J.-L. Gel-forming mucin interactome drives mucus viscoelasticity. Adv. Colloid Interface Sci. 2018, 252, 69–82. [Google Scholar] [CrossRef]
- Kang, J.-H.; Kim, Y.-Y.; Chang, J.-Y.; Kho, H.-S. Relationships between oral MUC1 expression and salivary hormones in burning mouth syndrome. Arch. Oral Biol. 2017, 78, 58–64. [Google Scholar] [CrossRef]
- Gendler, S.J.; Lancaster, C.A.; Taylor-Papadimitriou, J.; Duhig, T.; Peat, N.; Burchell, J.; Pemberton, L.; Lalani, E.N.; Wilson, D. Molecular cloning and expression of human tumor-associated polymorphic epithelial mucin. J. Biol. Chem. 1990, 265, 15286–15293. [Google Scholar] [CrossRef]
- Govindarajan, B.; Gipson, I.K. Membrane-tethered mucins have multiple functions on the ocular surface. Exp. Eye Res. 2010, 90, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Linden, S.K.; Sutton, P.; Karlsson, N.G.; Korolik, V.; McGuckin, M.A. Mucins in the mucosal barrier to infection. Mucosal Immunol. 2008, 1, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Lindén, S.K.; Sheng, Y.H.; Every, A.L.; Miles, K.M.; Skoog, E.C.; Florin, T.H.J.; Sutton, P.; McGuckin, M.A. MUC1 Limits Helicobacter pylori Infection both by Steric Hindrance and by Acting as a Releasable Decoy. PLoS Pathog. 2009, 5, e1000617. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.D. The Cytoplasmic Tail of MUC1: A Very Busy Place. Sci. Signal. 2008, 1, pe35. [Google Scholar] [CrossRef]
- Schmidt, P.; Dybal, J.; Trchová, M. Investigations of the hydrophobic and hydrophilic interactions in polymer–water systems by ATR FTIR and Raman spectroscopy. Vib. Spectrosc. 2006, 42, 278–283. [Google Scholar] [CrossRef]
- Wang, L.; Li, X.; Bruno, L.S.; Oppenheim, F.G.; Troxler, R.F.; Offner, G.D. MUC1 and MUC5B can form a protective mucin scaffold. In Proceedings of the 82nd IADR (International Association for Dental Research) General Session, Honolulu, HI, USA, 10–13 March 2004. [Google Scholar]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nivet, C.; Custovic, I.; Avoscan, L.; Bikker, F.J.; Bonnotte, A.; Bourillot, E.; Briand, L.; Brignot, H.; Heydel, J.-M.; Herrmann, N.; et al. Development of New Models of Oral Mucosa to Investigate the Impact of the Structure of Transmembrane Mucin-1 on the Mucosal Pellicle Formation and Its Physicochemical Properties. Biomedicines 2024, 12, 139. https://doi.org/10.3390/biomedicines12010139
Nivet C, Custovic I, Avoscan L, Bikker FJ, Bonnotte A, Bourillot E, Briand L, Brignot H, Heydel J-M, Herrmann N, et al. Development of New Models of Oral Mucosa to Investigate the Impact of the Structure of Transmembrane Mucin-1 on the Mucosal Pellicle Formation and Its Physicochemical Properties. Biomedicines. 2024; 12(1):139. https://doi.org/10.3390/biomedicines12010139
Chicago/Turabian StyleNivet, Clément, Irma Custovic, Laure Avoscan, Floris J. Bikker, Aline Bonnotte, Eric Bourillot, Loïc Briand, Hélène Brignot, Jean-Marie Heydel, Noémie Herrmann, and et al. 2024. "Development of New Models of Oral Mucosa to Investigate the Impact of the Structure of Transmembrane Mucin-1 on the Mucosal Pellicle Formation and Its Physicochemical Properties" Biomedicines 12, no. 1: 139. https://doi.org/10.3390/biomedicines12010139
APA StyleNivet, C., Custovic, I., Avoscan, L., Bikker, F. J., Bonnotte, A., Bourillot, E., Briand, L., Brignot, H., Heydel, J. -M., Herrmann, N., Lelièvre, M., Lesniewska, E., Neiers, F., Piétrement, O., Schwartz, M., Belloir, C., & Canon, F. (2024). Development of New Models of Oral Mucosa to Investigate the Impact of the Structure of Transmembrane Mucin-1 on the Mucosal Pellicle Formation and Its Physicochemical Properties. Biomedicines, 12(1), 139. https://doi.org/10.3390/biomedicines12010139