
Decoding Ribosome Heterogeneity: A New Horizon in Cancer Therapy



Abstract
:1. Introduction
2. Ribosome Heterogeneity in Normal Cellular Function
3. Oncoribosome

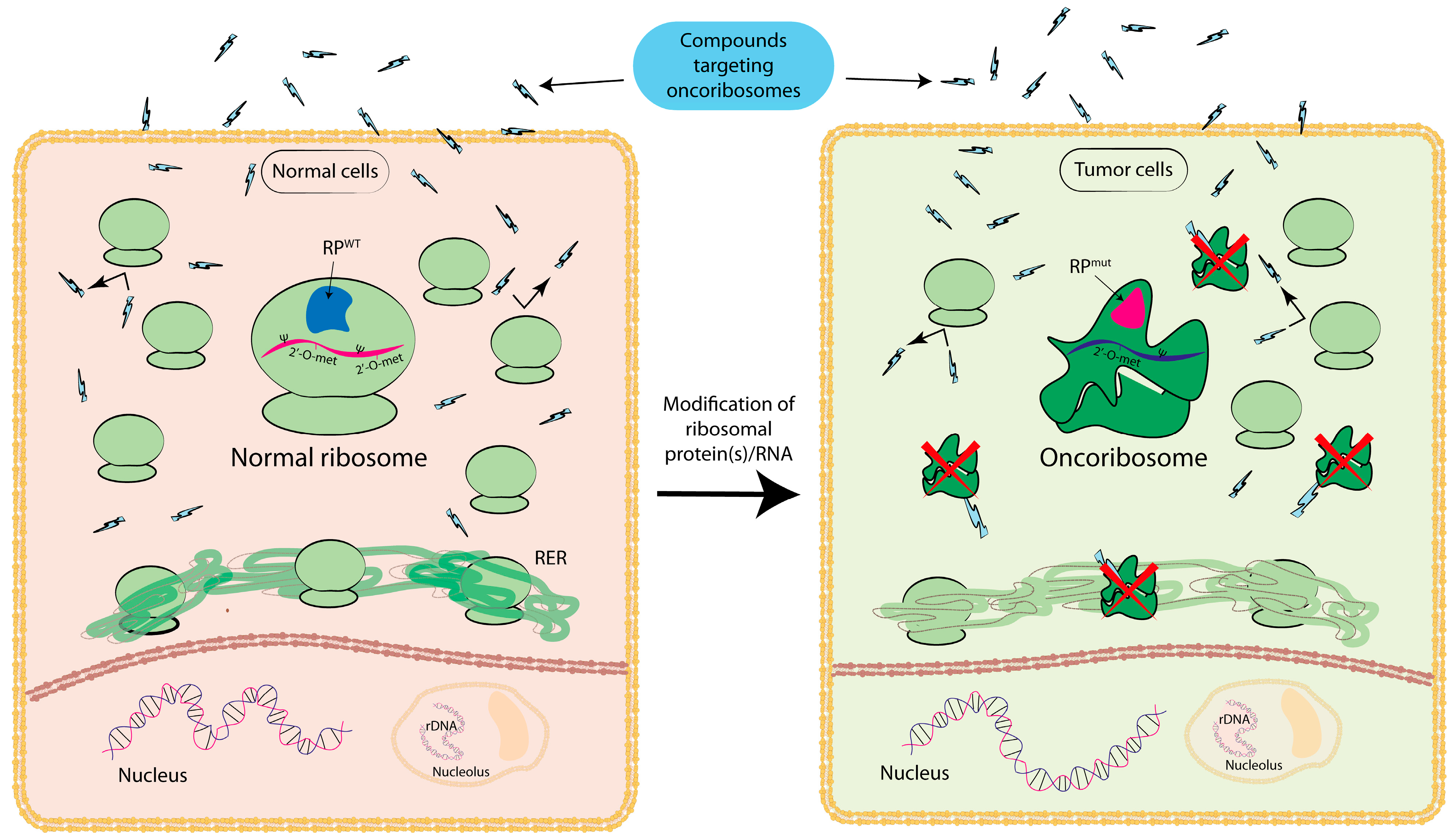{kind=link}
| Ribosomal Component | Type of Change | Ribosomal Protein | Phenotypic Consequences |
|---|---|---|---|
| Ribosomal proteins | Mutation(s) | RPL5 and RPL11 | Forty-nine cancer types [61]. |
| RPL10 | T-ALL and breast cancer [54,63]. | ||
| RPS20 | Colorectal cancer [56]. | ||
| RPL22 | Endometrial, colorectal, gastric, breast, and non-small-cell lung cancer [64]. | ||
| RPL23A | Uterine cancer [61]. | ||
| RPS15 | Primary chronic lymphocytic leukemia [62]. | ||
| Expression Level Variation | Increased/decreased RPS9, RPS14, RPL5, RPL10, RPL11, RPL15, and RPL39 | Initiation, progression, promotion of metastasis, regulation of translation, and cell cycles in circulating breast tumor cells [65]. | |
| Increased RPL17 | Colorectal cancer [66]. | ||
| Increased RPS3 | Impacts protein synthesis regulation and cancer progression in colorectal cancer [67]. | ||
| Increased RPS6 and RPL19 | Poor prognosis in cancer [40]. | ||
| Ribosomal RNA | Mutation(s) | 5S rRNA tip | Impairs the interaction between Rpf2–Rrs1 and 5S rRNA [68]. |
| 18S rRNA (1248.U) | Present in 45.9% of CRC patients and across >22 cancer types [69]. | ||
| Post-Translational Modification | 2’-O-Methylation | ||
| Pseudouridylation |
| ||
| Expression Level Variation | Increased rRNA levels | ||
| Structural Alteration | Truncation of 23S rRNA | Dysfunctional ribosome [79]. | |
| Environmental Influence | Therapeutic agents |
4. Ribosome Diversity as an Entry Point for Targeted Therapy
5. Discussion and Conclusions
Funding
Conflicts of Interest
References
- Lafontaine, D.L.J.; Tollervey, D. The Function and Synthesis of Ribosomes. Nat. Rev. Mol. Cell Biol. 2001, 2, 514–520. [Google Scholar] [CrossRef]
- Noller, H.F. RNA Structure: Reading the Ribosome. Science 2005, 309, 1508–1514. [Google Scholar] [CrossRef]
- Steitz, T.A. A Structural Understanding of the Dynamic Ribosome Machine. Nat. Rev. Mol. Cell Biol. 2008, 9, 242–253. [Google Scholar] [CrossRef]
- Bastide, A.; David, A. The Ribosome, (Slow) Beating Heart of Cancer (Stem) Cell. Oncogenesis 2018, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Nag, S.; Zhang, X.; Wang, M.H.; Wang, H.; Zhou, J.; Zhang, R. Ribosomal Proteins and Human Diseases: Pathogenesis, Molecular Mechanisms, and Therapeutic Implications. Med. Res. Rev. 2015, 35, 225–285. [Google Scholar] [CrossRef]
- Dolezal, J.M.; Dash, A.P.; Prochownik, E.V. Diagnostic and Prognostic Implications of Ribosomal Protein Transcript Expression Patterns in Human Cancers. BMC Cancer 2018, 18, 275. [Google Scholar] [CrossRef] [PubMed]
- Jiao, L.; Liu, Y.; Yu, X.-Y.; Pan, X.; Zhang, Y.; Tu, J.; Song, Y.-H.; Li, Y. Ribosome Biogenesis in Disease: New Players and Therapeutic Targets. Signal Transduct. Target. Ther. 2023, 8, 15. [Google Scholar] [CrossRef] [PubMed]
- Aspesi, A.; Ellis, S.R. Rare Ribosomopathies: Insights into Mechanisms of Cancer. Nat. Rev. Cancer 2019, 19, 228–238. [Google Scholar] [CrossRef]
- Kang, J.; Brajanovski, N.; Chan, K.T.; Xuan, J.; Pearson, R.B.; Sanij, E. Ribosomal Proteins and Human Diseases: Molecular Mechanisms and Targeted Therapy. Signal Transduct. Target. Ther. 2021, 6, 323. [Google Scholar] [CrossRef]
- Szaflarski, W.; Leśniczak-Staszak, M.; Sowiński, M.; Ojha, S.; Aulas, A.; Dave, D.; Malla, S.; Anderson, P.; Ivanov, P.; Lyons, S.M. Early rRNA Processing Is a Stress-Dependent Regulatory Event Whose Inhibition Maintains Nucleolar Integrity. Nucleic Acids Res. 2022, 50, 1033–1051. [Google Scholar] [CrossRef]
- Sulima, S.O.; Hofman, I.J.F.; De Keersmaecker, K.; Dinman, J.D. How Ribosomes Translate Cancer. Cancer Discov. 2017, 7, 1069–1087. [Google Scholar] [CrossRef]
- Taoka, M.; Nobe, Y.; Yamaki, Y.; Yamauchi, Y.; Ishikawa, H.; Takahashi, N.; Nakayama, H.; Isobe, T. The Complete Chemical Structure of Saccharomyces Cerevisiae rRNA: Partial Pseudouridylation of U2345 in 25S RRNA by SnoRNA SnR9. Nucleic Acids Res. 2016, 44, 8951–8961. [Google Scholar] [CrossRef]
- Barozzi, C.; Zacchini, F.; Corradini, A.G.; Morara, M.; Serra, M.; De Sanctis, V.; Bertorelli, R.; Dassi, E.; Montanaro, L. Alterations of Ribosomal RNA Pseudouridylation in Human Breast Cancer. NAR Cancer 2023, 5, zcad026. [Google Scholar] [CrossRef] [PubMed]
- Polikanov, Y.S.; Melnikov, S.V.; Söll, D.; Steitz, T.A. Structural Insights into the Role of rRNA Modifications in Protein Synthesis and Ribosome Assembly. Nat. Struct. Mol. Biol. 2015, 22, 342–344. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Montanaro, L. Turning Uridines around: Role of rRNA Pseudouridylation in Ribosome Biogenesis and Ribosomal Function. Biomolecules 2018, 8, 38. [Google Scholar] [CrossRef]
- Barozzi, C.; Zacchini, F.; Asghar, S.; Montanaro, L. Ribosomal RNA Pseudouridylation: Will Newly Available Methods Finally Define the Contribution of This Modification to Human Ribosome Plasticity? Front. Genet. 2022, 13, 920987. [Google Scholar] [CrossRef] [PubMed]
- Guo, H. Specialized Ribosomes and the Control of Translation. Biochem. Soc. Trans. 2018, 46, 855–869. [Google Scholar] [CrossRef]
- Dinman, J.D. Pathways to Specialized Ribosomes: The Brussels Lecture. J. Mol. Biol. 2016, 428, 2186–2194. [Google Scholar] [CrossRef]
- Genuth, N.R.; Barna, M. The Discovery of Ribosome Heterogeneity and Its Implications for Gene Regulation and Organismal Life. Mol. Cell 2018, 71, 364–374. [Google Scholar] [CrossRef]
- Wang, Q.; Mao, Y. Principles, Challenges, and Advances in Ribosome Profiling: From Bulk to Low-Input and Single-Cell Analysis. Adv. Biotechnol. 2023, 1, 6. [Google Scholar] [CrossRef]
- Xue, S.; Barna, M. Specialized Ribosomes: A New Frontier in Gene Regulation and Organismal Biology. Nat. Rev. Mol. Cell Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef]
- Segev, N.; Gerst, J.E. Specialized Ribosomes and Specific Ribosomal Protein Paralogs Control Translation of Mitochondrial Proteins. J. Cell Biol. 2018, 217, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Lafontaine, D.L.J. ‘View From A Bridge’: A New Perspective on Eukaryotic rRNA Base Modification. Trends Biochem. Sci. 2015, 40, 560–575. [Google Scholar] [CrossRef]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-Wide. Mol. Cell 2017, 67, 71–83.e7. [Google Scholar] [CrossRef]
- Fusco, C.M.; Desch, K.; Dörrbaum, A.R.; Wang, M.; Staab, A.; Chan, I.C.W.; Vail, E.; Villeri, V.; Langer, J.D.; Schuman, E.M. Neuronal Ribosomes Exhibit Dynamic and Context-Dependent Exchange of Ribosomal Proteins. Nat. Commun. 2021, 12, 6127. [Google Scholar] [CrossRef] [PubMed]
- Norris, K.; Hopes, T.; Aspden, J.L. Ribosome Heterogeneity and Specialization in Development. WIREs RNA 2021, 12, e1644. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, N.; Pusic, A.; Stumpf, C.R.; Shimizu, K.; Hsieh, A.C.; Xue, S.; Ishijima, J.; Shiroishi, T.; Barna, M. Ribosome-Mediated Specificity in Hox mRNA Translation and Vertebrate Tissue Patterning. Cell 2011, 145, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, J.C.; Zavolan, M. Patterns of Ribosomal Protein Expression Specify Normal and Malignant Human Cells. Genome Biol. 2016, 17, 236. [Google Scholar] [CrossRef]
- Locati, M.D.; Pagano, J.F.B.; Girard, G.; Ensink, W.A.; Van Olst, M.; Van Leeuwen, S.; Nehrdich, U.; Spaink, H.P.; Rauwerda, H.; Jonker, M.J.; et al. Expression of Distinct Maternal and Somatic 5.8S, 18S, and 28S rRNA Types during Zebrafish Development. RNA 2017, 23, 1188–1199. [Google Scholar] [CrossRef]
- Bellodi, C.; Kopmar, N.; Ruggero, D. Deregulation of Oncogene-Induced Senescence and P53 Translational Control in X-Linked Dyskeratosis Congenita. EMBO J. 2010, 29, 1865–1876. [Google Scholar] [CrossRef]
- Sinturel, F.; Gerber, A.; Mauvoisin, D.; Wang, J.; Gatfield, D.; Stubblefield, J.J.; Green, C.B.; Gachon, F.; Schibler, U. Diurnal Oscillations in Liver Mass and Cell Size Accompany Ribosome Assembly Cycles. Cell 2017, 169, 651–663.e14. [Google Scholar] [CrossRef]
- Rothschild, D.; Susanto, T.T.; Sui, X.; Spence, J.P.; Rangan, R.; Genuth, N.R.; Sinnott-Armstrong, N.; Wang, X.; Pritchard, J.K.; Barna, M. Diversity of Ribosomes at the Level of rRNA Variation Associated with Human Health and Disease. bioRxiv 2023. [Google Scholar] [CrossRef]
- Parks, M.M.; Kurylo, C.M.; Dass, R.A.; Bojmar, L.; Lyden, D.; Vincent, C.T.; Blanchard, S.C. Variant Ribosomal RNA Alleles Are Conserved and Exhibit Tissue-Specific Expression. Sci. Adv. 2018, 4, eaao0665. [Google Scholar] [CrossRef]
- Karamysheva, Z.N.; Tikhonova, E.B.; Grozdanov, P.N.; Huffman, J.C.; Baca, K.R.; Karamyshev, A.; Denison, R.B.; Macdonald, C.C.; Zhang, K.; Karamyshev, A.L. Polysome Profiling in Leishmania, Human Cells and Mouse Testis. J. Vis. Exp. 2018, 2018, e57600. [Google Scholar] [CrossRef]
- Rabany, O.; Nachmani, D. Small Nucleolar (Sno)RNA: Therapy Lays in Translation. Noncoding RNA 2023, 9, 35. [Google Scholar] [CrossRef] [PubMed]
- Stedman, A.; Beck-Cormier, S.; Le Bouteiller, M.; Raveux, A.; Vandormael-Pournin, S.; Coqueran, S.; Lejour, V.; Jarzebowski, L.; Toledo, F.; Robine, S.; et al. Ribosome Biogenesis Dysfunction Leads to P53-Mediated Apoptosis and Goblet Cell Differentiation of Mouse Intestinal Stem/Progenitor Cells. Cell Death Differ. 2015, 22, 1865–1876. [Google Scholar] [CrossRef]
- Turi, Z.; Lacey, M.; Mistrik, M.; Moudry, P. Impaired Ribosome Biogenesis: Mechanisms and Relevance to Cancer and Aging. Aging 2019, 11, 2512–2540. [Google Scholar] [CrossRef] [PubMed]
- Venturi, G.; Montanaro, L. How Altered Ribosome Production Can Cause or Contribute to Human Disease: The Spectrum of Ribosomopathies. Cells 2020, 9, 2300. [Google Scholar] [CrossRef] [PubMed]
- Catez, F.; Dalla Venezia, N.; Marcel, V.; Zorbas, C.; Lafontaine, D.L.J.; Diaz, J.-J. Ribosome Biogenesis: An Emerging Druggable Pathway for Cancer Therapeutics. Biochem. Pharmacol. 2019, 159, 74–81. [Google Scholar] [CrossRef]
- Elhamamsy, A.R.; Metge, B.J.; Alsheikh, H.A.; Shevde, L.A.; Samant, R.S. Ribosome Biogenesis: A Central Player in Cancer Metastasis and Therapeutic Resistance. Cancer Res. 2022, 82, 2344–2353. [Google Scholar] [CrossRef]
- Zisi, A.; Bartek, J.; Lindström, M.S. Targeting Ribosome Biogenesis in Cancer: Lessons Learned and Way Forward. Cancers 2022, 14, 2126. [Google Scholar] [CrossRef]
- Gilles, A.; Frechin, L.; Natchiar, K.; Biondani, G.; von Loeffelholz, O.; Holvec, S.; Malaval, J.-L.; Winum, J.-Y.; Klaholz, B.P.; Peyron, J.-F. Targeting the Human 80S Ribosome in Cancer: From Structure to Function and Drug Design for Innovative Adjuvant Therapeutic Strategies. Cells 2020, 9, 629. [Google Scholar] [CrossRef] [PubMed]
- Romaniello, D.; Gelfo, V.; Pagano, F.; Ferlizza, E.; Sgarzi, M.; Mazzeschi, M.; Morselli, A.; Miano, C.; D’Uva, G.; Lauriola, M. Senescence-Associated Reprogramming Induced by Interleukin-1 Impairs Response to EGFR Neutralization. Cell Mol. Biol. Lett. 2022, 27, 20. [Google Scholar] [CrossRef] [PubMed]
- Sulima, S.; Kampen, K.; De Keersmaecker, K. Cancer Biogenesis in Ribosomopathies. Cells 2019, 8, 229. [Google Scholar] [CrossRef]
- Gianni, F.; Belver, L.; Ferrando, A. The Genetics and Mechanisms of T-Cell Acute Lymphoblastic Leukemia. Cold Spring Harb. Perspect. Med. 2020, 10, a035246. [Google Scholar] [CrossRef] [PubMed]
- De Keersmaecker, K.; Sulima, S.O.; Dinman, J.D. Ribosomopathies and the Paradox of Cellular Hypo- to Hyperproliferation. Blood 2015, 125, 1377–1382. [Google Scholar] [CrossRef]
- Amsterdam, A.; Sadler, K.C.; Lai, K.; Farrington, S.; Bronson, R.T.; Lees, J.A.; Hopkins, N. Many Ribosomal Protein Genes Are Cancer Genes in Zebrafish. PLoS Biol. 2004, 2, E139. [Google Scholar] [CrossRef]
- Hofman, I.J.F.; Van Duin, M.; De Bruyne, E.; Fancello, L.; Mulligan, G.; Geerdens, E.; Garelli, E.; Mancini, C.; Lemmens, H.; Delforge, M.; et al. RPL5 on 1p22.1 Is Recurrently Deleted in Multiple Myeloma and Its Expression Is Linked to Bortezomib Response. Leukemia 2017, 31, 1706–1714. [Google Scholar] [CrossRef]
- Landau, D.A.; Tausch, E.; Taylor-Weiner, A.N.; Stewart, C.; Reiter, J.G.; Bahlo, J.; Kluth, S.; Bozic, I.; Lawrence, M.; Böttcher, S.; et al. Mutations Driving CLL and Their Evolution in Progression and Relapse. Nature 2015, 526, 525–530. [Google Scholar] [CrossRef]
- Xu, X.; Xiong, X.; Sun, Y. The Role of Ribosomal Proteins in the Regulation of Cell Proliferation, Tumorigenesis, and Genomic Integrity. Sci. China Life Sci. 2016, 59, 656–672. [Google Scholar] [CrossRef]
- Teng, T.; Mercer, C.A.; Hexley, P.; Thomas, G.; Fumagalli, S. Loss of Tumor Suppressor RPL5/RPL11 Does Not Induce Cell Cycle Arrest but Impedes Proliferation Due to Reduced Ribosome Content and Translation Capacity. Mol. Cell Biol. 2013, 33, 4660–4671. [Google Scholar] [CrossRef]
- Oršolić, I.; Bursać, S.; Jurada, D.; Drmić Hofman, I.; Dembić, Z.; Bartek, J.; Mihalek, I.; Volarević, S. Cancer-Associated Mutations in the Ribosomal Protein L5 Gene Dysregulate the HDM2/P53-Mediated Ribosome Biogenesis Checkpoint. Oncogene 2020, 39, 3443–3457. [Google Scholar] [CrossRef] [PubMed]
- Girardi, T.; Vereecke, S.; Sulima, S.O.; Khan, Y.; Fancello, L.; Briggs, J.W.; Schwab, C.; De Beeck, J.O.; Verbeeck, J.; Royaert, J.; et al. The T-Cell Leukemia-Associated Ribosomal RPL10 R98S Mutation Enhances JAK-STAT Signaling. Leukemia 2018, 32, 809–819. [Google Scholar] [CrossRef]
- De Keersmaecker, K.; Atak, Z.K.; Li, N.; Vicente, C.; Patchett, S.; Girardi, T.; Gianfelici, V.; Geerdens, E.; Clappier, E.; Porcu, M.; et al. Exome Sequencing Identifies Mutation in CNOT3 and Ribosomal Genes RPL5 and RPL10 in T-Cell Acute Lymphoblastic Leukemia. Nat. Genet. 2013, 45, 186–190. [Google Scholar] [CrossRef]
- Liu, Y.; Easton, J.; Shao, Y.; Maciaszek, J.; Wang, Z.; Wilkinson, M.R.; McCastlain, K.; Edmonson, M.; Pounds, S.B.; Shi, L.; et al. The Genomic Landscape of Pediatric and Young Adult T-Lineage Acute Lymphoblastic Leukemia. Nat. Genet. 2017, 49, 1211–1218. [Google Scholar] [CrossRef]
- Nieminen, T.T.; O’Donohue, M.F.; Wu, Y.; Lohi, H.; Scherer, S.W.; Paterson, A.D.; Ellonen, P.; Abdel-Rahman, W.M.; Valo, S.; Mecklin, J.P.; et al. Germline Mutation of RPS20, Encoding a Ribosomal Protein, Causes Predisposition to Hereditary Nonpolyposis Colorectal Carcinoma without DNA Mismatch Repair Deficiency. Gastroenterology 2014, 147, 595–598. [Google Scholar] [CrossRef]
- Rao, S.; Cai, K.Q.; Stadanlick, J.E.; Greenberg-Kushnir, N.; Solanki-Patel, N.; Lee, S.Y.; Fahl, S.P.; Testa, J.R.; Wiest, D.L. Ribosomal Protein Rpl22 Controls the Dissemination of T-Cell Lymphoma. Cancer Res. 2016, 76, 3387–3396. [Google Scholar] [CrossRef] [PubMed]
- Novetsky, A.P.; Zighelboim, I.; Thompson, D.M.; Powell, M.A.; Mutch, D.G.; Goodfellow, P.J. Frequent Mutations in the RPL22 Gene and Its Clinical and Functional Implications. Gynecol. Oncol. 2013, 128, 470–474. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Tuominen, I.; van Dijk-Bos, K.; Sanjabi, B.; van der Sluis, T.; van der Zee, A.G.; Hollema, H.; Zazula, M.; Sijmons, R.H.; Aaltonen, L.A.; et al. High Frequency of RPL22 Mutations in Microsatellite-Unstable Colorectal and Endometrial Tumors. Hum. Mutat. 2014, 35, 1442–1445. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, N.; Bertrand, D.; Hillmer, A.M.; Zang, Z.J.; Yao, F.; Jacques, P.É.; Teo, A.S.M.; Cutcutache, I.; Zhang, Z.; Lee, W.H.; et al. Whole-Genome Reconstruction and Mutational Signatures in Gastric Cancer. Genome Biol. 2012, 13, R115. [Google Scholar] [CrossRef]
- Fancello, L.; Kampen, K.R.; Hofman, I.J.F.; Verbeeck, J.; De Keersmaecker, K. The Ribosomal Protein Gene RPL5 Is a Haploinsufficient Tumor Suppressor in Multiple Cancer Types. Oncotarget 2017, 8, 14462–14478. [Google Scholar] [CrossRef]
- Ntoufa, S.; Gerousi, M.; Laidou, S.; Psomopoulos, F.; Tsiolas, G.; Moysiadis, T.; Papakonstantinou, N.; Mansouri, L.; Anagnostopoulos, A.; Stavrogianni, N.; et al. RPS15 Mutations Rewire RNA Translation in Chronic Lymphocytic Leukemia. Blood Adv. 2021, 5, 2788–2792. [Google Scholar] [CrossRef]
- Sulima, S.O.; Kampen, K.R.; Vereecke, S.; Pepe, D.; Fancello, L.; Verbeeck, J.; Dinman, J.D.; De Keersmaecker, K. Ribosomal Lesions Promote Oncogenic Mutagenesis. Cancer Res. 2019, 79, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Fang, Z.; Liao, P.; Zhou, X.; Xiong, J.; Zeng, S.; Lu, H. Cancer-Mutated Ribosome Protein L22 (RPL22/EL22) Suppresses Cancer Cell Survival by Blocking P53-MDM2 Circuit. Oncotarget 2017, 8, 90651–90661. [Google Scholar] [CrossRef] [PubMed]
- Lüchtenborg, A.M.; Metzger, P.; Cosenza Contreras, M.; Oria, V.; Biniossek, M.L.; Lindner, F.; Fröhlich, K.; Malyi, A.; Erbes, T.; Gensch, N.; et al. Krüppel-like Factor 7 Influences Translation and Pathways Involved in Ribosomal Biogenesis in Breast Cancer. Breast Cancer Res. 2022, 24, 65. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.J.; Seo, Y.R.; Seo, D.; Park, S.Y.; Seo, J.H.; Jeon, E.H.; Kim, S.W.; Park, K.U.; Koo, D.B.; Kim, S.; et al. RPL17 Promotes Colorectal Cancer Proliferation and Stemness through ERK and NEK2/β-Catenin Signaling Pathways. J. Cancer 2022, 13, 2570–2583. [Google Scholar] [CrossRef] [PubMed]
- El Khoury, W.; Nasr, Z. Deregulation of Ribosomal Proteins in Human Cancers. Biosci. Rep. 2021, 41, BSR20211577. [Google Scholar] [CrossRef] [PubMed]
- Thoms, M.; Mitterer, V.; Kater, L.; Falquet, L.; Beckmann, R.; Kressler, D.; Hurt, E. Suppressor Mutations in Rpf2–Rrs1 or Rpl5 Bypass the Cgr1 Function for Pre-Ribosomal 5S RNP-Rotation. Nat. Commun. 2018, 9, 4094. [Google Scholar] [CrossRef] [PubMed]
- Babaian, A.; Rothe, K.; Girodat, D.; Minia, I.; Djondovic, S.; Milek, M.; Spencer Miko, S.E.; Wieden, H.J.; Landthaler, M.; Morin, G.B.; et al. Loss of M1acp3Ψ Ribosomal RNA Modification Is a Major Feature of Cancer. Cell Rep. 2020, 31, 107611. [Google Scholar] [CrossRef] [PubMed]
- Nombela, P.; Miguel-López, B.; Blanco, S. The Role of M6A, M5C and Ψ RNA Modifications in Cancer: Novel Therapeutic Opportunities. Mol. Cancer 2021, 20, 18. [Google Scholar] [CrossRef]
- Janin, M.; Coll-SanMartin, L.; Esteller, M. Disruption of the RNA Modifications That Target the Ribosome Translation Machinery in Human Cancer. Mol. Cancer 2020, 19, 70. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Yi, H.; Mao, C.; Meng, Q.; Wu, X.; Li, S.; Xue, J. Research Progress of RNA Methylation Modification in Colorectal Cancer. Front. Pharmacol. 2022, 13, 903699. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Aroua, N.; Liu, Y.; Rohde, C.; Cheng, J.; Wirth, A.K.; Fijalkowska, D.; Göllner, S.; Lotze, M.; Yun, H.; et al. A Dynamic rRNA Ribomethylome Drives Stemness in Acute Myeloid Leukemia. Cancer Discov. 2023, 13, 332–347. [Google Scholar] [CrossRef]
- Jansson, M.D.; Häfner, S.J.; Altinel, K.; Tehler, D.; Krogh, N.; Jakobsen, E.; Andersen, J.V.; Andersen, K.L.; Schoof, E.M.; Ménard, P.; et al. Regulation of Translation by Site-Specific Ribosomal RNA Methylation. Nat. Struct. Mol. Biol. 2021, 28, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Yoon, A.; Peng, G.; Brandenburg, Y.; Zollo, O.; Xu, W.; Rego, E.; Ruggero, D. Impaired Control of IRES-Mediated Translation in X-Linked Dyskeratosis Congenita. Science 2006, 312, 902–906. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, Y.; Lv, Q.; Zhang, J.; Wang, Q.; Gao, F.; Hou, H.; Zhang, H.; Zhang, W.; Li, L. Overexpression of Ribosomal RNA in the Development of Human Cervical Cancer Is Associated with rDNA Promoter Hypomethylation. PLoS ONE 2016, 11, e0163340. [Google Scholar] [CrossRef] [PubMed]
- Uemura, M.; Zheng, Q.; Koh, C.M.; Nelson, W.G.; Yegnasubramanian, S.; De Marzo, A.M. Overexpression of Ribosomal RNA in Prostate Cancer Is Common but Not Linked to rDNA Promoter Hypomethylation. Oncogene 2012, 31, 1254–1263. [Google Scholar] [CrossRef] [PubMed]
- Karahan, G.; Sayar, N.; Gozum, G.; Bozkurt, B.; Konu, O.; Yulug, I.G. Relative Expression of rRNA Transcripts and 45S RDNA Promoter Methylation Status Are Dysregulated in Tumors in Comparison with Matched-Normal Tissues in Breast Cancer. Oncol. Rep. 2015, 33, 3131–3145. [Google Scholar] [CrossRef]
- Willi, J.; Küpfer, P.; Eviquoz, D.; Fernandez, G.; Katz, A.; Leumann, C.; Polacek, N. Oxidative Stress Damages rRNA inside the Ribosome and Differentially Affects the Catalytic Center. Nucleic Acids Res. 2018, 46, 1945–1957. [Google Scholar] [CrossRef] [PubMed]
- Butler, P.; Pascheto, I.; Lizzi, M.; St-Onge, R.; Lanner, C.; Guo, B.; Masilamani, T.; Pritzker, L.B.; Kovala, A.T.; Parissenti, A.M. RNA Disruption Is a Widespread Phenomenon Associated with Stress-Induced Cell Death in Tumour Cells. Sci. Rep. 2023, 13, 1711. [Google Scholar] [CrossRef]
- Therizols, G.; Bash-Imam, Z.; Panthu, B.; Machon, C.; Vincent, A.; Ripoll, J.; Nait-Slimane, S.; Chalabi-Dchar, M.; Gaucherot, A.; Garcia, M.; et al. Alteration of Ribosome Function upon 5-Fluorouracil Treatment Favors Cancer Cell Drug-Tolerance. Nat. Commun. 2022, 13, 173. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, J.; Dang, Q.; Wang, X.; Chen, J.; Lin, X.; Yang, N.; Du, J.; Shi, H.; Liu, Y.; et al. Ribosomal Protein RPL5 Regulates Colon Cancer Cell Proliferation and Migration through MAPK/ERK Signaling Pathway. BMC Mol. Cell Biol. 2022, 23, 48. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhang, L.; Zhou, D.; Zhang, J.; Lin, Q.; Guan, W.; Zhang, J.; Ren, W.; Xu, G. Biological Function of Ribosomal Protein L10 on Cell Behavior in Human Epithelial Ovarian Cancer. J. Cancer 2018, 9, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Chabronova, A.; Akker, G.G.; Ripmeester, E.G.; Housmans, B.A.; Cremers, A.; Surtel, D.A.; Wichapong, K.; Rhijn, L.W.; Marchand, V.; Motorin, Y.; et al. Evidence of Specialized Ribosomes in Osteoarthritic Chondrocytes. Osteoarthr. Cartil. 2021, 29, S313–S314. [Google Scholar] [CrossRef]
- Breznak, S.M.; Peng, Y.; Deng, L.; Kotb, N.M.; Flamholz, Z.; Rapisarda, I.T.; Martin, E.T.; LaBarge, K.A.; Fabris, D.; Gavis, E.R.; et al. H/ACA SnRNP–Dependent Ribosome Biogenesis Regulates Translation of Polyglutamine Proteins. Sci. Adv. 2023, 9, eade5492. [Google Scholar] [CrossRef] [PubMed]
- Sloan, K.E.; Warda, A.S.; Sharma, S.; Entian, K.-D.; Lafontaine, D.L.J.; Bohnsack, M.T. Tuning the Ribosome: The Influence of rRNA Modification on Eukaryotic Ribosome Biogenesis and Function. RNA Biol. 2017, 14, 1138–1152. [Google Scholar] [CrossRef] [PubMed]
- Decatur, W.A.; Fournier, M.J. rRNA Modifications and Ribosome Function. Trends Biochem. Sci. 2002, 27, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Monaco, P.; Marcel, V.; Diaz, J.-J.; Catez, F. 2′-O-Methylation of Ribosomal RNA: Towards an Epitranscriptomic Control of Translation? Biomolecules 2018, 8, 106. [Google Scholar] [CrossRef]
- Marcel, V.; Kielbassa, J.; Marchand, V.; Natchiar, K.S.; Paraqindes, H.; Nguyen Van Long, F.; Ayadi, L.; Bourguignon-Igel, V.; Lo Monaco, P.; Monchiet, D.; et al. Ribosomal RNA 2′O-Methylation as a Novel Layer of Inter-Tumour Heterogeneity in Breast Cancer. NAR Cancer 2020, 2, zcaa036. [Google Scholar] [CrossRef] [PubMed]
- Motorin, Y.; Quinternet, M.; Rhalloussi, W.; Marchand, V. Constitutive and Variable 2′-O-Methylation (Nm) in Human Ribosomal RNA. RNA Biol. 2021, 18, 88–97. [Google Scholar] [CrossRef]
- Schosserer, M.; Minois, N.; Angerer, T.B.; Amring, M.; Dellago, H.; Harreither, E.; Calle-Perez, A.; Pircher, A.; Gerstl, M.P.; Pfeifenberger, S.; et al. Methylation of Ribosomal RNA by NSUN5 Is a Conserved Mechanism Modulating Organismal Lifespan. Nat. Commun. 2015, 6, 6158. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Galbiati, A.; Treré, D.; Montanaro, L. The Importance of Being (Slightly) Modified: The Role of rRNA Editing on Gene Expression Control and Its Connections with Cancer. Biochim. et Biophys. Acta Rev. Cancer 2016, 1866, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Birkedal, U.; Christensen-Dalsgaard, M.; Krogh, N.; Sabarinathan, R.; Gorodkin, J.; Nielsen, H. Profiling of Ribose Methylations in RNA by High-Throughput Sequencing. Angew. Chem. Int. Ed. 2015, 54, 451–455. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Sharma, S.; Watzinger, P.; Hartmann, J.D.; Kötter, P.; Entian, K.D. Mapping of Complete Set of Ribose and Base Modifications of Yeast rRNA by RP-HPLC and Mung Bean Nuclease Assay. PLoS ONE 2016, 11, e0168873. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, V.; Plunkett, W.; Cortes, J.E. Omacetaxine: A Protein Translation Inhibitor for Treatment of Chronic Myelogenous Leukemia. Clin. Cancer Res. 2014, 20, 1735–1740. [Google Scholar] [CrossRef]
- Chen, R.; Guo, L.; Chen, Y.; Jiang, Y.; Wierda, W.G.; Plunkett, W. Homoharringtonine Reduced Mcl-1 Expression and Induced Apoptosis in Chronic Lymphocytic Leukemia. Blood 2011, 117, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-J.; Malina, A.; Pelletier, J. C-Myc and EIF4F Constitute a Feedforward Loop That Regulates Cell Growth: Implications for Anticancer Therapy. Cancer Res. 2009, 69, 7491–7494. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.J.; Cencic, R.; Mills, J.R.; Robert, F.; Pelletier, J. C-Myc and EIF4F Are Components of a Feedforward Loop That Links Transcription and Translation. Cancer Res. 2008, 68, 5326–5334. [Google Scholar] [CrossRef]
- Tong, H.; Ren, Y.; Zhang, F.; Jin, J. Homoharringtonine Affects the JAK2-STAT5 Signal Pathway through Alteration of Protein Tyrosine Kinase Phosphorylation in Acute Myeloid Leukemia Cells. Eur. J. Haematol. 2008, 81, 259–266. [Google Scholar] [CrossRef]
- Lü, S.; Wang, J. Homoharringtonine and Omacetaxine for Myeloid Hematological Malignancies. J. Hematol. Oncol. 2014, 7, 2. [Google Scholar] [CrossRef]
- Yan, F.; Yu, Y.; Chow, D.C.; Palzkill, T.; Madoux, F.; Hodder, P.; Chase, P.; Griffin, P.R.; O’Malley, B.W.; Lonard, D.M. Identification of Verrucarin a as a Potent and Selective Steroid Receptor Coactivator-3 Small Molecule Inhibitor. PLoS ONE 2014, 9, e95243. [Google Scholar] [CrossRef] [PubMed]
- Palanivel, K.; Kanimozhi, V.; Kadalmani, B.; Akbarsha, M.A. Verrucarin A, a Protein Synthesis Inhibitor, Induces Growth Inhibition and Apoptosis in Breast Cancer Cell Lines MDA-MB-231 and T47D. Biotechnol. Lett. 2013, 35, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Woldemichael, G.M.; Turbyville, T.J.; Vasselli, J.R.; Linehan, W.M.; Mcmahon, J.B. Lack of a Functional VHL Gene Product Sensitizes Renal Cell Carcinoma Cells to the Apoptotic Effects of the Protein Synthesis Inhibitor Verrucarin A. Neoplasia 2012, 14, 771–777. [Google Scholar] [CrossRef]
- Oda, T.; Namikoshi, M.; Akano, K.; Kobayashi, H.; Honma, Y.; Kasahara, T. Verrucarin A Inhibition of MAP Kinase Activation in a PMA-Stimulated Promyelocytic Leukemia Cell Line. Mar. Drugs 2005, 3, 64–73. [Google Scholar] [CrossRef]
- Myasnikov, A.G.; Kundhavai Natchiar, S.; Nebout, M.; Hazemann, I.; Imbert, V.; Khatter, H.; Peyron, J.F.; Klaholz, B.P. Structure-Function Insights Reveal the Human Ribosome as a Cancer Target for Antibiotics. Nat. Commun. 2016, 7, 12856. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gelfo, V.; Venturi, G.; Zacchini, F.; Montanaro, L. Decoding Ribosome Heterogeneity: A New Horizon in Cancer Therapy. Biomedicines 2024, 12, 155. https://doi.org/10.3390/biomedicines12010155
Gelfo V, Venturi G, Zacchini F, Montanaro L. Decoding Ribosome Heterogeneity: A New Horizon in Cancer Therapy. Biomedicines. 2024; 12(1):155. https://doi.org/10.3390/biomedicines12010155
Chicago/Turabian StyleGelfo, Valerio, Giulia Venturi, Federico Zacchini, and Lorenzo Montanaro. 2024. "Decoding Ribosome Heterogeneity: A New Horizon in Cancer Therapy" Biomedicines 12, no. 1: 155. https://doi.org/10.3390/biomedicines12010155
APA StyleGelfo, V., Venturi, G., Zacchini, F., & Montanaro, L. (2024). Decoding Ribosome Heterogeneity: A New Horizon in Cancer Therapy. Biomedicines, 12(1), 155. https://doi.org/10.3390/biomedicines12010155