

Innovative Biomedical and Technological Strategies for the Control of Bacterial Growth and Infections

 ,
,
Abstract
:1. Introduction
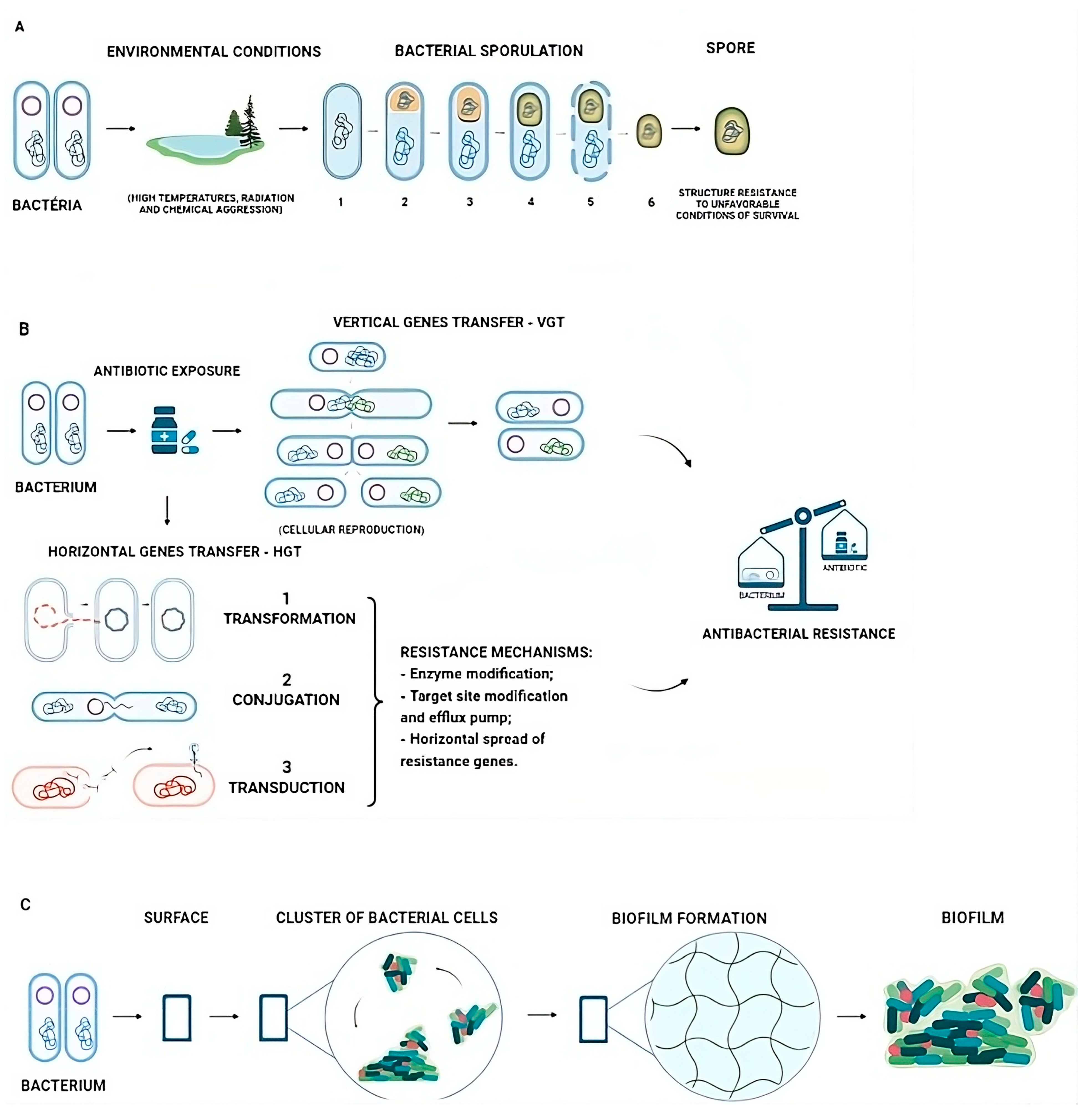
2. Bacterial Resistant Mechanisms and Antibiotics

3. Innovations and Strategies in the Development of New Antibiotics
3.1. Use of Genomics and Resistant Genes for Antibiotics
3.2. Use of Micro/Nanoparticles for Antibiotics
3.2.1. Microparticles
3.2.2. Nanoparticles

{kind=link}
{kind=link}
{kind=link}
| Study | Encapsulating Agent | Antibiotic | Encapsulation Method | Particle Size | Bacteria(s) Target | Antibacterial Outcome |
|---|---|---|---|---|---|---|
| Oliveira et al., 2017 [54] | Silver nitrate (AgNO3) | Ampicillin | Colloidal dispersion | 93 nm | Escherichia coli | The antibiotic interrupted the cell wall synthesis, inhibiting enzymes, and the silver nanoparticles acted as a cytotoxic agent for the bacteria. |
| Knoblauch et al., 2021 [66] | Brominated carbon (BrCND) | No antibiotic | Nanoincorporation (via carbon halogenation with bromine) | 365 nm | Staphylococcus aureus and Listeria monocytogenes and Escherichia coli | The brominated carbon nanoparticles showed antimicrobial activity through the innovative method of photodynamic inactivation of bacteria with membrane rupture and release of reactive nitrogen species (synergical damage). |
| Armênia et al., 2018 [56] | Iron oxide (FeO) | Teicoplanin | Co-precipitation | 10.5 nm | Staphylococcus aureus, Enterococcus faecalis and Bacillus subtilis and Escherichia coli | The isolated antibiotic presented antimicrobial activity in the short term, while the nanoparticles showed promising and prolonged antimicrobial activity because they caused a synergical effect with membrane and DNA damage. |
| Zheng et al., 2019 [57] | Gold (Au) | Daptomycin | Aggregation–Inducted Emission | 190 nm | Staphylococcus aureus | The nanoparticles showed an enhanced bactericidal synergistic effect with the ability to disrupt bacterial membranes and produce ROS. |
| Chen et al., 2013 [60] | Graphene oxide (GO) | No antibiotic | Nanoprecipitation | 300–600 nm | Xanthomonas oryzae pv. oryzae | Due to their sharp edges, the nanoparticles have shown superior bactericidal capacity at extremely low concentrations by rupturing the cell membrane. |
| El-Alfy et al., 2020 [63] | Chitosan | Tetracycline, Gentamicin, and Ciprofloxacin | Ionic gelation | 3–4 nm | Gram-positive and Gram-negative bacteria | The three nanoantibiotics effectively acted on the inhibition of the growth of Gram-positive and Gram-negative bacteria due to chemical damage due to a charge imbalance. |
| Gaspar et al., 2018 [64] | Polylactic-co-glycolic acid (PLGA) | Vancomycin and Meropenem | Double emulsion–solvent evaporation | 284.2 nm | Staphylococcus aureus and Pseudomonas aeruginosa | The nanoparticles of both antibiotics showed effective and efficient antibacterial action compared to the isolated actives, probably causing chemical damage. |
3.3. Use of Computational Simulation for Antibiotics
In Silico Studies and Antimicrobial Peptides (AMPs)
3.4. Other Strategies for Applying In Silico Studies
4. Natural Antimicrobials and New Strategies
4.1. Polymeric Films
4.2. Alternative Medical Solutions
4.3. Probiotics
4.4. Bacteriophages
4.5. Vaccines
4.6. Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Rossolini, G.M.; Arena, F.; Pecile, P.; Pollini, S. Update on the antibiotic resistance crisis. Curr. Opin. Pharmacol. 2014, 18, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Khan, A.U. Global economic impact of antibiotic resistance: A review. J. Glob. Antimicrob. Resist. 2019, 19, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, G. Detection and various environmental factors of antibiotic resistance gene horizontal transfer. Environ. Res. 2022, 212, 113267. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Giacobbe, D.; Giamarellou, H.; Viscoli, C.; Daikos, G.; Dimopoulos, G.; De Rosa, F.; Giamarellos-Bourboulis, E.; Rossolini, G.; Righi, E.; et al. Management of KPC-producing Klebsiella pneumoniae infections. Clin. Microbiol. Infect. 2018, 24, 133–144. [Google Scholar] [CrossRef]
- Antimicrobial Resistance Collaborators. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Giacobbe, D.R.; Giani, T.; Bassetti, M.; Marchese, A.; Viscoli, C.; Rossolini, G.M. Rapid microbiological tests for bloodstream infections due to multidrug resistant Gram-negative bacteria: Therapeutic implications. Clin. Microbiol. Infect. 2020, 26, 713–722. [Google Scholar] [CrossRef] [PubMed]
- Savjani, J.K.; Gajjar, A.K.; Savjani, K.T. Mechanisms of resistance: Useful tool to design antibacterial agents for drug-resistant bacteria. Mini. Rev. Med. Chem. 2009, 9, 194–205. [Google Scholar] [CrossRef]
- Garvey, M. Bacteriophages and the one health approach to combat multidrug resistance: Is this the way? Antibiotics 2020, 9, 414. [Google Scholar] [CrossRef]
- Zohra, T.; Numan, M.; Ikram, A.; Salman, M.; Khan, T.; Din, M.; Salman, M.; Farooq, A.; Amir, A.; Ali, M. Cracking the Challenge of Antimicrobial Drug Resistance with CRISPR/Cas9, Nanotechnology and Other Strategies in ESKAPE Pathoges. Microorganisms 2021, 9, 954. [Google Scholar] [CrossRef]
- Ndagi, U.; Falaki, A.A.; Abdullahi, M.; Lawal, M.M.; Soliman, M.E. Antibiotic resistance: Bioinformatics-based understanding as a functional strategy for drug design. RSC Adv. 2020, 10, 18451–18468. [Google Scholar] [CrossRef]
- Ngara, T.R.; Zhang, H. Recent advances in function-based metagenomic screening. Genom. Proteom. Bioinform. 2018, 16, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Sizar, O.; Leslie, S.W.; Unakal, C.G. Gram Positive Bacteria. Stat Pearls [Internet]. 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK470553/ (accessed on 8 November 2022).
- Yang, D.C.; Blair, K.M.; Salama, N.R. Staying in shape: The impact of cell shape on bacterial survival in diverse environments. Microbiol. Mol. Biol. Rev. 2016, 80, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Segev, E.; Smith, Y.; Ben-Yehuda, S. RNA dynamics in aging bacterial spores. Cell 2012, 148, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.F.; Zhang, S.; Pan, B.; Liu, X.; Feng, L.S. 4-Quinolone derivatives and their activities against Gram positive pathogens. Eur. J. Med. Chem. 2018, 143, 710–723. [Google Scholar] [CrossRef] [PubMed]
- Klobucar, K.; Brown, E.D. New potentiators of ineffective antibiotics: Targeting the Gram-negative outer membrane to overcome intrinsic resistance. Curr. Opin. Chem. Biol. 2022, 66, 102099. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson-Palme, J.; Kristiansson, E.; Larsson, D.G.J. Environmental factors influencing the development and spread of antibiotic resistance. FEMS Microbiol. Rev. 2017, 42, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Borodovich, T.; Shkoporov, A.N.; Ross, R.P.; Hill, C. Phage-mediated horizontal gene transfer and its implications for the human gut microbiome. Gastroenterol. Rep. 2022, 10. [Google Scholar] [CrossRef]
- Kannan, M.; Rajarathinam, K.; Venkatesan, S.; Dheeba, B.; Maniraj, A. Silver Iodide Nanoparticles as an Antibiofilm Agent—A Case Study on Gram-Negative Biofilm-Forming Bacteria. In Nanostructures for Antimicrobial Therapy; Elsevier: Amsterdam, The Netherlands, 2017; pp. 435–456. [Google Scholar]
- Tocheva, E.I.; Ortega, D.R.; Jensen, G.J. Sporulation, bacterial cell envelopes and the origin of life. Nat. Rev. Microbiol. 2016, 14, 535–542. [Google Scholar] [CrossRef]
- Cag, Y.; Caskurlu, H.; Fan, Y.; Cao, B.; Vahaboglu, H. Resistance mechanisms. Ann. Transl. Med. 2016, 4, 326. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Yang, X.; Li, J.; Lv, N.; Liu, F.; Wu, J.; Lin, I.Y.; Wu, N.; Weimer, B.C.; Gao, G.F.; et al. The bacterial mobile resistome transfer network connecting the animal and human microbiomes. Appl. Environ. Microbiol. 2016, 82, 6672–6681. [Google Scholar] [CrossRef] [PubMed]
- Watkins, R.R.; Bonomo, R.A. Overview: Global and Local Impact of Antibiotic Resistance. Infect. Dis. Clin. N. Am. 2016, 30, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Bouki, C.; Venieri, D.; Diamadopoulos, E. Detection and fate of antibiotic resistant bacteria in wastewater treatment plants: A review. Ecotoxicol. Environ. Saf. 2013, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pawlowski, A.C.; Johnson, J.W.; Wright, G.D. Evolving medicinal chemistry strategies in antibiotic discovery. Curr. Opin. Biotechnol. 2016, 42, 108–117. [Google Scholar] [CrossRef]
- Yao, G.; Cao, C.; Zhang, M.; Kwok, L.Y.; Zhang, H.; Zhang, W. Lactobacillus casei Zhang exerts probiotic effects to antibiotic-treated rats. Comput. Struct. Biotechnol. J. 2021, 19, 5888–5897. [Google Scholar] [CrossRef]
- World Health Organization. WHO Model List of Essential Medicines for Children: 6th List [Internet] 2017. Available online: https://apps.who.int/iris/handle/10665/273825 (accessed on 10 November 2022).
- World Health Organization Team. 2021 AWaRe Classification [Internet]. 2021. Available online: https://www.who.int/publications/i/item/2021-aware-classification (accessed on 10 November 2022).
- Brooks, B.D.; Brooks, A.E. Therapeutic strategies to combat antibiotic resistance. Adv. Drug Deliv. Rev. 2014, 78, 14–27. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial antibiotic resistance: The most critical pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Ernst, C.M.; Peschel, A. MprF-mediated daptomycin resistance. Int. J. Med. Microbiol. 2019, 309, 359–363. [Google Scholar] [CrossRef]
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef]
- Lambert, P.A. Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. J. R. Soc. Med. 2002, 95, 22–26. [Google Scholar] [PubMed]
- Handelsman, J. Metagenomics: Application of genomics to uncultured microorganisms. Microbiol. Mol. Biol. Rev. 2004, 68, 669–685. [Google Scholar] [CrossRef]
- Chernov, V.M.; Chernova, O.A.; Mouzykantov, A.A.; Lopukhov, L.L.; Aminov, R.I. Omics of antimicrobials and antimicrobial resistance. Expert Opin. Drug Discov. 2019, 14, 455–468. [Google Scholar] [CrossRef] [PubMed]
- Garza, D.R.; Dutilh, B.E. From cultured to uncultured genome sequences: Metagenomics and modeling microbial ecosystems. Cell. Mol. Life Sci. 2015, 72, 4287–4308. [Google Scholar] [CrossRef] [PubMed]
- Zipperer, A.; Konnerth, M.C.; Laux, C.; Berscheid, A.; Janek, D.; Weidenmaier, C.; Burian, M.; Schilling, N.A.; Slavetinsky, C.; Mareschal, M.; et al. Human commensals producing a novel antibiotic impair pathogen colonization. Nature 2016, 535, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Hover, B.M.; Kim, S.H.; Katz, M.; Charlop-Powers, Z.; Owen, J.G.; Ternei, M.A.; Maniko, J.; Estrela, A.B.; Molina, H.; Park, S.; et al. Culture-independent discovery of the malacidins as calcium dependent antibiotics with activity against multidrug-resistant Gram-positive pathogens. Nat. Microbiol. 2018, 3, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Nikolaou, E.; Hubbard, A.M.; Botelho, J.; Marschall, T.A.M.; Ferreira, D.M.; Roberts, A.P. Antibiotic resistance is associated with integrative and conjugative elements and genomic islands in naturally circulating Streptococcus pneumoniae isolates from adults in Liverpool, UK. Genes 2020, 11, 625. [Google Scholar] [CrossRef]
- Harrison, E.M.; Ba, X.; Coll, F.; Blane, B.; Restif, O.; Carvell, H.; Köser, C.U.; Jamrozy, D.; Reuter, S.; Lovering, A.; et al. Genomic identification of cryptic susceptibility to penicillins and β-lactamase inhibitors in methicillin-resistant Staphylococcus aureus. Nat. Microbiol. 2019, 4, 1680–1691. [Google Scholar] [CrossRef]
- Genilloud, O. Natural products discovery and potential for new antibiotics. Curr. Opin. Microbiol. 2019, 51, 81–87. [Google Scholar] [CrossRef]
- Lewis, K. New approaches to antimicrobial discovery. Biochem. Pharmacol. 2017, 134, 87–98. [Google Scholar] [CrossRef]
- Ali Said, F.; Bousserrhine, N.; Alphonse, V.; Michely, L.; Belbekhouche, S. Antibiotic loading and development of antibacterial capsules by using porous CaCO3 microparticles as starting material. Int. J. Pharm. 2020, 579, 119175. [Google Scholar] [CrossRef] [PubMed]
- Josino, M.A.A.; da Silva, C.R.; Neto, J.B.d.A.; Barroso, F.D.D.; da Silva, L.J.; Cavalcanti, B.C.; de Moraes, M.O.; Brito, D.H.A.; Ricardo, N.M.P.S.; Júnior, H.V.N. Development and in vitro evaluation of microparticles of fluoxetine in galactomannan against biofilms of S. aureus methicilin resistant. Carbohydr. Polym. 2021, 252, 117184. [Google Scholar] [CrossRef] [PubMed]
- Rezić, I.; Škoc, M.S.; Majdak, M.; Jurić, S.; Stracenski, K.S.; Vlahoviček-Kahlina, K.; Vinceković, M. ICP-MS Determination of Antimicrobial Metals in Microcapsules. Molecules 2022, 27, 3219. [Google Scholar] [CrossRef] [PubMed]
- Kost, B.; Kunicka-Styczyńska, A.; Plucińska, A.; Rajkowska, K.; Basko, M.; Brzeziński, M. Microfluidic preparation of antimicrobial microparticles composed of l-lactide/1,3-dioxolane (co)polymers loaded with quercetin. Food Chem. 2022, 396, 133639. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Silva, H.R.L.; Junior, E.C.; Serafim, J.A.; Meneguin, A.B.; Chorilli, M.; Macedo, W.C.; Teixeira, S.R.; Guastalli, E.A.L.; Soares, N.M.; et al. Alginate-based microparticles coated with HPMCP/AS cellulose-derivatives enable the Ctx(Ile21)-Ha antimicrobial peptide application as a feed additive. Int. J. Biol. Macromol. 2021, 183, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Sousa, J.; Pais, A.; Vitorino, C. Nanomedicine: Principles, properties, and regulatory issues. Front. Chem. 2018, 6, 360. [Google Scholar] [CrossRef] [PubMed]
- Mamun, M.M.; Sorinolu, A.J.; Munir, M.; Vejerano, E.P. Nanoantibiotics: Functions and properties at the nanoscale to combat antibiotic resistance. Front. Chem. 2021, 9, 687660. [Google Scholar] [CrossRef]
- Liu, W.; Wang, R.; Vedarethinam, V.; Huang, L.; Qian, K. Advanced materials for precise detection and antibiotic-free inhibition of bacteria. Mater. Today Adv. 2022, 13, 100204. [Google Scholar] [CrossRef]
- Herrera, G.; Peña-Bahamonde, J.; Paudel, S.; Rodrigues, D.F. The role of nanomaterials and antibiotics in microbial resistance and environmental impact: An overview. Curr. Opin. Chem. Eng. 2021, 33, 100707. [Google Scholar] [CrossRef]
- Oliveira, J.A.; Saito, A.; Bido, A.T.; Kobarg, J.; Stassen, H.K.; Cardoso, M.B. Defeating bacterial resistance and preventing mammalian cells toxicity through rational design of antibiotic-functionalized nanoparticles. Sci. Rep. 2017, 7, 1326. [Google Scholar] [CrossRef]
- Zhang, S.; Lin, L.; Huang, X.; Lu, Y.G.; Zheng, D.L.; Feng, Y. Antimicrobial properties of metal nanoparticles and their oxide materials and their applications in oral biology. J. Nanomater. 2022, 2022, 2063265. [Google Scholar] [CrossRef]
- Armenia, I.; Marcone, G.L.; Berini, F.; Orlandi, V.T.; Pirrone, C.; Martegani, E.; Gornati, R.; Bernardini, G.; Marinelli, F. Magnetic nanoconjugated teicoplanin: A novel tool for bacterial infection site targeting. Front. Microbiol. 2018, 9, 2270. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, W.; Chen, Y.; Li, C.; Jiang, H.; Wang, X. Conjugating gold nanoclusters and antimicrobial peptides: From aggregation-induced emission to antibacterial synergy. J. Colloid. Interface Sci. 2019, 546, 1–10. [Google Scholar] [CrossRef]
- Mohammed, H.; Kumar, A.; Bekyarova, E.; Al-Hadeethi, Y.; Zhang, X.; Chen, M.; Ansari, M.S.; Cochis, A.; Rimondini, L. Antimicrobial Mechanisms and Effectiveness of Graphene and Graphene-Functionalized Biomaterials. A Scope Review. Front. Bioeng. Biotechnol. 2020, 8, 465. [Google Scholar] [CrossRef]
- Zou, X.; Zhang, L.; Wang, Z.; Luo, Y. Mechanisms of the Antimicrobial Activities of Graphene Materials. J. Am. Chem. Soc. 2016, 138, 2064–2077. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, X.; Han, H. A new function of graphene oxide emerges: Inactivating phytopathogenic bacterium Xanthomonas oryzae pv. Oryzae. J. Nanopart. Res. 2013, 15, 1658. [Google Scholar] [CrossRef]
- Je, J.Y.; Park, P.J.; Kim, B.; Kim, S.K. Antihypertensive activity of chitin derivatives. Biopolymers 2006, 83, 250–254. [Google Scholar] [CrossRef]
- Kohda, N.; Inoue, S.; Noda, T.; Saito, T. Effects of a chitosan intake on the fecal excretion of dioxins and fat in rats. Biosci. Biotechnol. Biochem. 2012, 76, 1544–1548. [Google Scholar] [CrossRef]
- El-Alfy, E.A.; El-Bisi, M.K.; Taha, G.M.; Ibrahim, H.M. Preparation of biocompatible chitosan nanoparticles loaded by tetracycline, gentamycin and ciprofloxacin as novel drug delivery system for improvement the antibacterial properties of cellulose based fabrics. Int. J. Biol. Macromol. 2020, 161, 1247–1260. [Google Scholar] [CrossRef]
- Gaspar, L.M.D.A.C.; Dórea, A.C.S.; Droppa-Almeida, D.; de Melo Silva, I.S.; Montoro, F.E.; Alves, L.L.; Hernández-Macedo, M.-L.; Padilha, F.F. Development and characterization of PLGA nanoparticles containing antibiotics. J. Nanopart. Res. 2018, 20, 289. [Google Scholar] [CrossRef]
- Sengar, P.; Garcia-Tapia, K.; Chauhan, K.; Jain, A.; Juarez-Moreno, K.; Borbón-Nuñez, H.A.; Tiznado, H.; Contreras, O.E.; Hirata, G.A. Dual-photosensitizer coupled nanoscintillator capable of producing type I and type II ROS for next generation photodynamic therapy. J. Colloid. Interface Sci. 2019, 536, 586–597. [Google Scholar] [CrossRef] [PubMed]
- Knoblauch, R.; Harvey, A.; Ra, E.; Greenberg, K.M.; Lau, J.; Hawkins, E.; Geddes, C.D. Antimicrobial carbon nanodots: Photodynamic inactivation and dark antimicrobial effects on bacteria by brominated carbon nanodots. Nanoscale 2021, 7, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chen, Z.; Gao, Z.; Tammina, S.K.; Yang, Y. Nanozymes used for antimicrobials and their applications. Colloids Surf. B Biointerfaces 2020, 195, 111252. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Zhu, S.; Liu, Y.; Yin, W.; Gu, Z.; Zhao, Y. An overview of the use of nanozymes in antibacterial applications. Chem. Eng. J. 2021, 418, 129431. [Google Scholar] [CrossRef]
- Naskar, A.; Kim, K. Nanomaterials as delivery vehicles and components of new strategies to combat bacterial infections: Advantages and limitations. Microorganisms 2019, 7, 356. [Google Scholar] [CrossRef]
- Lei, R.; Hou, J.; Chen, Q.; Yuan, W.; Cheng, B.; Sun, Y.; Jin, Y.; Ge, L.; Ben-Sasson, S.A.; Chen, J.; et al. Self-assembling myristoylated human α-defensin 5 as a next-generation nanobiotics potentiates therapeutic efficacy in bacterial infection. Acs. Nano. 2018, 12, 5284–5296. [Google Scholar] [CrossRef]
- Elfadil, D.; Elkhatib, W.F.; El-sayyad, G.S. Promising advances in nanobiotic-based formulations for drug specific targeting against multidrug resistant microbes and biofilm-associated infections. Microb. Pathog. 2022, 170, 105721. [Google Scholar] [CrossRef]
- Yin, K.; Wang, Q.; Luv, M.; Chen, L. Microorganism remediation strategies towards heavy metals. Chem. Eng. J. 2019, 360, 1553–1563. [Google Scholar] [CrossRef]
- Pallen, M.J. Microbial bioinformatics 2020. Microb. Biotechnol. 2016, 9, 681–686. [Google Scholar] [CrossRef]
- Fricke, W.F.; Rasko, D.A. Bacterial genome sequencing in the clinic: Bioinformatic challenges and solutions. Nat. Rev. Genet. 2014, 15, 49–55. [Google Scholar] [CrossRef]
- Peredo-Lovillo, A.; Hernández-Mendoza, A.; Vallejo-Cordoba, B.; Romero-Luna, H.E. Conventional and in silico approaches to select promising food-derived bioactive peptides: A review. Food Chem. X. 2021, 13, 100183. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.E.; Ameen, F.; Aabed, K.; Suliman, R.S.; Alghamdi, S.S.; Safhi, F.A.; Alshaya, D.S.; Alafari, H.A.; Jalal, A.S.; Alosaimi, A.A.; et al. In-silico predicting as a tool to develop plant-based biomedicines and nanoparticles: Lycium shawii metabolites. Biomed. Pharmacother. 2022, 150, 113008. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhu, B.; Wang, W.; Tan, H.; Yin, H. Characterization of a new oligoalginate lyase from marine bacterium Vibrio sp. Int. J. Biol. Macromol. 2018, 112, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Felnagle, E.A.; Jackson, E.E.; Chan, Y.A.; Podevels, A.M.; Berti, A.D.; McMahon, M.D.; Thomas, M.G. Nonribosomal peptide synthetases involved in the production of medically relevant natural products. Mol. Pharm. 2008, 5, 191–211. [Google Scholar] [CrossRef] [PubMed]
- Khabthani, S.; Rolain, J.M.; Merhej, V. In silico/in vitro strategies leading to the discovery of new nonribosomal peptide and polyketide antibiotics active against human pathogens. Microorganisms 2021, 5, 2297. [Google Scholar] [CrossRef]
- Tracanna, V.; de Jong, A.; Medema, M.H.; Kuipers, O.P. Mining prokaryotes for antimicrobial compounds: From diversity to function. FEMS Microbiol. Rev. 2017, 41, 417–429. [Google Scholar] [CrossRef]
- Idakwo, G.; Luttrell, J.; Chen, M.; Hong, H.; Zhou, Z.; Gong, P.; Zhang, C. A review on machine learning methods for in silico toxicity prediction. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2018, 36, 169–191. [Google Scholar] [CrossRef]
- Zwickl, C.M.; Graham, J.C.; Jolly, R.A.; Bassan, A.; Ahlberg, E.; Amberg, A.; Anger, L.T.; Beilke, L.; Bellion, P.; Brigo, A.; et al. Principles and procedures for assessment of acute toxicity incorporating in silico methods. Comput. Toxicol. 2022, 24, 100237. [Google Scholar] [CrossRef]
- Detunji, C.O.; Mitembo, W.P.; Egbuna, C.; Rao, G.M.N. In Silico Modeling as a Tool to Predict and Characterize Plant Toxicity. In Poisonous Plants and Phytochemicals in Drug Discovery; Wiley: Hoboken, NJ, USA, 2020; pp. 367–378. [Google Scholar] [CrossRef]
- Segalin, J.; Arsand, J.B.; Jank, L.; Schwalm, C.S.; Streit, L.; Pizzolato, T.M. In silico toxicity evaluation for transformation products of antimicrobials, from aqueous photolysis degradation. Sci. Total Environ. 2022, 828, 154109. [Google Scholar] [CrossRef]
- da Cunha, N.B.; Cobacho, N.B.; Viana, J.F.; Lima, L.A.; Sampaio, K.B.; Dohms, S.S.; Ferreira, A.C.; de la Fuente-Núñez, C.; Costa, F.F.; Franco, O.L.; et al. The next generation of antimicrobial peptides (AMPs) as molecular therapeutic tools for the treatment of diseases with social and economic impacts. Drug Discov. Today 2017, 22, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Meher, P.K.; Sahu, T.K.; Saini, V.; Rao, A.R. Predicting antimicrobial peptides with improved accuracy by incorporating the compositional, physico-chemical and structural features into Chou’s general PseAAC. Sci. Rep. 2017, 7, 42362. [Google Scholar] [CrossRef] [PubMed]
- Espeche, J.C.; Martínez, M.; Maturana, P.; Cutró, A.; Semorile, L.; Maffia, P.C.; Hollmann, A. Unravelling the mechanism of action of “de novo” designed peptide P1 with model membranes and gram-positive and gram-negative bacteria. Arch. Biochem. Biophys. 2020, 693, 108549. [Google Scholar] [CrossRef]
- Savelyeva, A.; Ghavami, S.; Davoodpour, P.; Asoodeh, A.; Los, M.J. An overview of Brevinin superfamily: Structure, function and clinical perspectives. Adv. Exp. Med. Biol. 2014, 818, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.P.; Hancock, R.E. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Wang, G. Database-Guided Discovery of Potent Peptides to Combat HIV-1 or Superbugs. Pharmaceuticals 2013, 6, 728–758. [Google Scholar] [CrossRef]
- Seo, M.D.; Won, H.S.; Kim, J.H.; Mishig-Ochir, T.; Lee, B.J. Antimicrobial peptides for therapeutic applications: A review. Molecules 2012, 18, 12276–12286. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD2: The updated antimicrobial peptide database and its application in peptide design. Nucleic. Acids. Res. 2009, 37, 933–937. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.H.; Ribeiro, S.M.; Nolasco, D.O.; De La Fuente-Núñez, C.; Felício, M.R.; Gonçalves, S.; Matos, C.O.; Liao, L.M.; Santos, N.C.; Hancock, R.E.; et al. A polyalanine peptide derived from polar fish with anti-infectious activities. Sci. Rep. 2016, 6, 21385, Erratum in Sci. Rep. 2016, 20, 28216. [Google Scholar] [CrossRef]
- Porto, W.F.; Irazazabal, L.; Alves, E.S.F.; Ribeiro, S.M.; Matos, C.O.; Pires, Á.S.; Fensterseifer, I.C.M.; Miranda, V.J.; Haney, E.F.; Humblot, V.; et al. In silico optimization of a guava antimicrobial peptide enables combinatorial exploration for peptide design. Nat. Commun. 2018, 9, 1490. [Google Scholar] [CrossRef]
- Erdem Büyükkiraz, M.; Kesmen, Z. Antimicrobial peptides (AMPs): A promising class of antimicrobial compounds. J. Appl. Microbiol. 2022, 132, 1573–1596. [Google Scholar] [CrossRef]
- Torrent, M.; Andreu, D.; Nogués, V.M.; Boix, E. Connecting peptide physicochemical and antimicrobial properties by a rational prediction model. PLoS ONE 2011, 6, 16968. [Google Scholar] [CrossRef]
- Fernandes, F.C.; Rigden, D.J.; Franco, O.L. Prediction of antimicrobial peptides based on the adaptive neuro-fuzzy inference system application. Biopolymers 2012, 98, 280–287. [Google Scholar] [CrossRef]
- Ong, Z.Y.; Wiradharma, N.; Yang, Y.Y. Strategies employed in the design and optimization of synthetic antimicrobial peptide amphiphiles with enhanced therapeutic potentials. Adv. Drug Deliv. Rev. 2014, 78, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, P.; Glossop, H.; Meikle, T.G.; Aburto-Medina, A.; Conn, C.E.; Sarojini, V.; Valery, C. Molecular engineering of antimicrobial peptides: Microbial targets, peptide motifs and translation opportunities. Biophys. Rev. 2021, 13, 35–69. [Google Scholar] [CrossRef]
- Veltri, D.; Kamath, U.; Shehu, A. Deep learning improves antimicrobial peptide recognition. Bioinformatics 2018, 34, 2740–2747. [Google Scholar] [CrossRef] [PubMed]
- Dawood, H.M.; Ibrahim, R.S.; Shawky, E.; Hammoda, H.M.; Metwally, A.M. Integrated in silico-in vitro strategy for screening of some traditional Egyptian plants for human aromatase inhibitors. J. Ethnopharmacol. 2018, 224, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.P.; Rees, S.; Kalindjian, S.B.; Philpott, K.L. Principles of early drug discovery. Br J. Pharmacol. 2011, 162, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Miethke, M.; Pieroni, M.; Weber, T.; Brönstrup, M.; Hammann, P.; Halby, L.; Arimondo, P.B.; Glaser, P.; Aigle, B.; Bode, H.B.; et al. Towards the sustainable discovery and development of new antibiotics. Nat. Rev. Chem. 2021, 5, 726–749. [Google Scholar] [CrossRef]
- Reddy, A.S.; Zhang, S. Polypharmacology: Drug discovery for the future. Expert Rev. Clin. Pharmacol. 2013, 6, 41–47. [Google Scholar] [CrossRef]
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Rozenbaum, R.T.; Su, L.; Umerska, A.; Eveillard, M.; Håkansson, J.; Mahlapuu, M.; Huang, F.; Liu, J.; Zhang, Z.; Shi, L.; et al. Antimicrobial synergy of monolaurin lipid nanocapsules with adsorbed antimicrobial peptides against Staphylococcus aureus biofilms in vitro is absent in vivo. J. Control. Release. 2019, 293, 73–83. [Google Scholar] [CrossRef]
- Sakthivel, M.; Palani, P. Isolation, purification and characterization of antimicrobial protein from seedlings of Bauhinia purpurea L. Int. J. Biol. Macromol. 2016, 86, 390–401. [Google Scholar] [CrossRef]
- Anumudu, C.; Hart, A.; Miri, T.; Onyeaka, H. Recent Advances in the Application of the Antimicrobial Peptide Nisin in the Inactivation of Spore-Forming Bacteria in Foods. Molecules 2021, 26, 5552. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, A.M.d.S.; Segundo, V.H.d.O.; Aguiar, A.J.F.C.; Piuvezam, G.; Passos, T.S.; Damasceno, K.S.F.d.S.F.d.S.C.; Morais, A.H.d.A. Antibacterial action mechanisms and mode of trypsin inhibitors: A systematic review. J. Enzym. Inhib. Med. Chem. 2022, 37, 749–759. [Google Scholar] [CrossRef]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.W.; Harper, D.; et al. Alternatives to antibiotics-a pipeline portfolio review. Lancet. Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Nisa, M.; Dar, R.A.; Fomda, B.A.; Nazir, R. Combating food spoilage and pathogenic microbes via bacteriocins: A natural and eco-friendly substitute to antibiotics. Food Control. 2023, 149, 109710. [Google Scholar] [CrossRef]
- Taylor, A.W.; Coveney, J.; Ward, P.R.; Henderson, J.; Meyer, S.B.; Pilkington, R.; Gill, T.K. Fruit and vegetable consumption—the influence of aspects associated with trust in food and safety and quality of food. Public. Health Nutr. 2012, 15, 208–217. [Google Scholar] [CrossRef]
- Ministério da Saúde (BR). Surtos de Doenças de Transmissão Hídrica e Alimentar no Brasil. Informe 2022 [Internet]. Brasília: MS; 2022 14 p. Available online: https://www.gov.br/saude/pt-br/assuntos/saude-de-a-a-z/d/dtha/publicacoes/surtos-de-doencas-de-transmissao-hidrica-e-alimentar-no-brasil-informe-2022 (accessed on 14 November 2022).
- Jeevahan, J.; Chandrasekaran, M. Nanoedible films for food packaging: A review. J. Mater. Sci. 2019, 54, 12290–12318. [Google Scholar] [CrossRef]
- Gul, O.; Saricaoglu, F.T.; Besir, A.; Atalar, I.; Yazici, F. Effect of ultrasound treatment on the properties of nano-emulsion films obtained from hazelnut meal protein and clove essential oil. Ultrason. Sonochem. 2018, 41, 466–474. [Google Scholar] [CrossRef]
- Bourbon, A.I.; Costa, M.J.; Maciel, L.C.; Pastrana, L.; Vicente, A.A.; Cerqueira, M.A. Active carboxymethylcellulose-based edible films: Influence of free and encapsulated curcumin on films’ properties. Foods 2021, 10, 1512. [Google Scholar] [CrossRef]
- Bonilla, J.; Atarés, L.; Vargas, M.; Chiralt, A. Properties of wheat starch film-forming dispersions and films as affected by chitosan addition. J. Food. Eng. 2013, 114, 303–312. [Google Scholar] [CrossRef]
- Kumar, M.R. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Mateescu, M.; Baixe, S.; Garnier, T.; Jierry, L.; Ball, V.; Haikel, Y.; Metz-Boutigue, M.H.; Nardin, M.; Schaaf, P.; Etienne, O.; et al. Antibacterial Peptide-Based Gel for Prevention of Medical Implanted-Device Infection. PLoS ONE 2015, 10, 0145143. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; Costa, F.; Pirttilä, A.M.; Tejesvi, M.V.; Martins, M.C.L. Prevention of urinary catheter-associated infections by coating antimicrobial peptides from crowberry endophytes. Sci. Rep. 2019, 9, 10753. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.K.; Heo, S.J.; Peredo, A.P.; Mauck, R.L.; Dodge, G.R.; Lee, D. Stretch-responsive adhesive microcapsules for strain-regulated antibiotic release from fabric wound dressings. Biomater. Sci. 2021, 9, 5136–5143. [Google Scholar] [CrossRef]
- Gomes, R.N.; Borges, I.; Pereira, A.T.; Maia, A.F.; Pestana, M.; Magalhães, F.D.; Pinto, A.M.; Gonçalves, I.C. Antimicrobial graphene nanoplatelets coatings for silicone catheters. Carbon 2018, 139, 635–647. [Google Scholar] [CrossRef]
- Kabirian, F.; Brouki Milan, P.; Zamanian, A.; Heying, R.; Mozafari, M. Nitric oxide-releasing vascular grafts: A therapeutic strategy to promote angiogenic activity and endothelium regeneration. Acta Biomater. 2019, 92, 82–91. [Google Scholar] [CrossRef]
- Murugan, D.; Rangasamy, L. The use of antimicrobial biomaterials as a savior from post-operative vascular graft-related infections: A review. Results Eng. 2022, 16, 100662. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Forssten, S.; Hibberd, A.A.; Lyra, A.; Stahl, B. Probiotic approach to prevent antibiotic resistance. Ann. Med. 2016, 48, 246–255. [Google Scholar] [CrossRef]
- Homan, M.; Orel, R. Are probiotics useful in Helicobacter pylori eradication? World J. Gastroenterol. 2015, 21, 10644–10653. [Google Scholar] [CrossRef]
- Wu, M.; Dong, Q.; Ma, Y.; Yang, S.; Aslam, M.Z.; Liu, Y.; Li, Z. Potential antimicrobial activities of probiotics and their derivatives against Listeria monocytogenes in food field: A review. Food Res. Int. 2022, 160, 111733. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Yang, H. using probiotics as supplementation for helicobacter pylori antibiotic therapy. Int. J. Mol. Sci. 2020, 21, 1136. [Google Scholar] [CrossRef] [PubMed]
- Martín, I.; Rodríguez, A.; Delgado, J.; Córdoba, J.J. Strategies for Biocontrol of Listeria monocytogenes Using Lactic Acid Bacteria and Their Metabolites in Ready-to-Eat Meat- and Dairy-Ripened Products. Foods 2022, 11, 542. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yang, H. Effect of Bifidobacterium breve in Combination With Different Antibiotics on Clostridium difficile. Front. Microbiol. 2018, 9, 2953. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Meng, L.; Li, Y.; Huang, H. Strategies for applying probiotics in the antibiotic management of Clostridioides difficile infection. Food Funct. 2023, 14, 8711–8733. [Google Scholar] [CrossRef] [PubMed]
- Ling Lou, X.; Luo, Q.; He, Z.; Sun, M.; Sun, J. Recent advances in bacteriophage-based therapeutics: Insight into the post-antibiotic era. Acta Pharm. Sin. B 2022, 12, 4348–4364. [Google Scholar] [CrossRef]
- Xia, H.; Yang, H.; Yan, N.; Hou, W.; Wang, H.; Wang, X.; Wang, H.; Zhou, M. Bacteriostatic effects of phage F23s1 and its endolysin on Vibrio parahaemolyticus. J. Basic Microbiol. 2022, 62, 963–974. [Google Scholar] [CrossRef]
- Donlan, R.M. Preventing Biofilms of Clinically Relevant Organisms Using Bacteriophage. Trends Microbiol. 2009, 17, 66–72. [Google Scholar] [CrossRef]
- Micoli, F.; Bagnoli, F.; Rappuoli, R.; Serruto, D. The role of vaccines in combatting antimicrobial resistance. Nat. Rev. Microbiol. 2021, 19, 287–302. [Google Scholar] [CrossRef]
- Vekemans, J.; Hasso-Agopsowicz, M.; Kang, G.; Hausdorff, W.P.; Fiore, A.; Tayler, E.; Klemm, E.J.; Laxminarayan, R.; Srikantiah, P.; Friede, M.; et al. Leveraging vaccines to reduce antibiotic use and prevent antimicrobial resistance: A world health organization action framework. Clin. Infect. Dis. 2021, 73, 1011–1017. [Google Scholar] [CrossRef]
- Kennedy, D.A.; Read, A.F. Why the evolution of vaccine resistance is less of a concern than the evolution of drug resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 12878–12886. [Google Scholar] [CrossRef]
- Bagnoli, F.; Payne, D.J. Reaction: Alternative modalities to address antibiotic-resistant pathogens. Chem 2017, 3, 369–372. [Google Scholar] [CrossRef]
- David, E.B.; Steven, B.; David, S.; Rino, R. Antimicrobial resistance and the role of vaccines. Proc. Natl. Acad. Sci. 2018, 115, 12868–12871. [Google Scholar] [CrossRef]
- Bergmann, I.E.; Malirat, V.; Pedemonte, A.; Maradei, E. Challenges in foot-and-mouth disease virus strain selection as an input to attain broad vaccine intraserotype cross-protection. Expert Rev. Vaccines 2021, 20, 13–22. [Google Scholar] [CrossRef]
- Buckley, B.; Henschke, N.; Bergman, H.; Skidmore, B.; Klemm, E.; Villanueva, G.; Garritty, C.; Paul, M. Impact of vaccination on antibiotic usage: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2019, 25, 1213–1225. [Google Scholar] [CrossRef] [PubMed]
- Greene, A.C. CRISPR-Based Antibacterials: Transforming bacterial defense into offense. Trends Biotechnol. 2018, 36, 127–130. [Google Scholar] [CrossRef] [PubMed]
- Donohoue, P.D.; Barrangou, R.; May, A.P. Advances in industrial biotechnology using CRISPR-Cas Systems. Trends Biotechnol. 2018, 36, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Citorik, R.J.; Mimee, M.; Lu, T.K. Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases. Nat. Biotechnol. 2014, 32, 1141–1145. [Google Scholar] [CrossRef]
- Fuente-Núñez, C.; Lu, T.K. CRISPR–Cas9 technology: Applications in genome engineering, development of sequence-specific antimicrobials, and future prospects. Integr. Biol. 2017, 9, 109–122. [Google Scholar] [CrossRef]
- Drulis-Kawa, Z.; Majkowska-Skrobek, G.; Maciejewska, B.; Delatte, A.S.; Lavigne, R. Learning from Bacteriophages—Advantages and Limitations of Phage and Phage-Encoded Protein Applications. Curr. Protein Pept. Sci. 2012, 13, 699–722. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, J.; Xing, Y.; Zhao, Y.; Liu, Y. Development of delivery strategies for CRISPR-Cas9 genome editing. BMEMat 2023, 1, e12025. [Google Scholar] [CrossRef]


| Mechanism of Action | Main Target Bacteria * | Main Antibiotics |
|---|---|---|
| Cell Wall They inhibit the synthesis of bacterial peptidoglycan cell walls. They act on enzymes called penicillin-binding proteins (PBPs) involved in cross-linking bacterial cell walls. The beta-lactam ring portion of these antibiotics irreversibly binds to PBPs, inhibiting the cross-linking of peptidoglycan and triggering bacterial death via autolysis. | Staphylococcus aureus, Streptococcus pneumoniae, Listeria monocytogenes, Pseudomonas aeruginosa, and Acinetobacter. | Penicillin, Carbapenem, and Cephalosporin |
| Staphylococcus epidermidis, Staphylococcus haemolyticus and Staphylococcus aureus | Vancomycin and Teicoplanin | |
| Cell Membrane They bind to phospholipids in the cytoplasmic membrane, altering their barrier function. | Pseudomonas aeruginosa Staphylococcus aureus, and Bacillus subtilis | Colomycin, Colistin, and Daptomycin |
| Inhibition of Protein Synthesis Binding to a susceptible organism’s 30S or 50S ribosomal subunit interferes with the binding of aminoacyl-tRNA to the mRNA/ribosome complex, thus interrupting bacterial protein synthesis. | Aerobic Gram-negative bacteria | Neomycin, Streptomycin, Kanamycin, Tobramycin, and Amikacin |
| Oxytetracycline and Chlortetracycline | ||
| Streptococcus pneumoniae, Streptococcus pyogenes, Haemophilus influenzae and Moraxella catarrhalis | Macrolides, Lincosamides, and Streptogramin B | |
| Chloramphenicol | ||
| Nucleic Acids They directly inhibit bacterial DNA synthesis by inhibiting two enzymes: topoisomerase II (DNA gyrase—catalyzes the negative supercoiling of closed double-stranded circular DNA) and topoisomerase IV (the unwinding of DNA after chromosomal duplication). | Gram-negative bacteria | Ciprofloxacin, Gemifloxacin, Levofloxacin, Moxifloxacin, and Ofloxacin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matias, L.L.R.; Damasceno, K.S.F.d.S.C.; Pereira, A.S.; Passos, T.S.; Morais, A.H.d.A. Innovative Biomedical and Technological Strategies for the Control of Bacterial Growth and Infections. Biomedicines 2024, 12, 176. https://doi.org/10.3390/biomedicines12010176
Matias LLR, Damasceno KSFdSC, Pereira AS, Passos TS, Morais AHdA. Innovative Biomedical and Technological Strategies for the Control of Bacterial Growth and Infections. Biomedicines. 2024; 12(1):176. https://doi.org/10.3390/biomedicines12010176
Chicago/Turabian StyleMatias, Lídia Leonize Rodrigues, Karla Suzanne Florentino da Silva Chaves Damasceno, Annemberg Salvino Pereira, Thaís Souza Passos, and Ana Heloneida de Araujo Morais. 2024. "Innovative Biomedical and Technological Strategies for the Control of Bacterial Growth and Infections" Biomedicines 12, no. 1: 176. https://doi.org/10.3390/biomedicines12010176
APA StyleMatias, L. L. R., Damasceno, K. S. F. d. S. C., Pereira, A. S., Passos, T. S., & Morais, A. H. d. A. (2024). Innovative Biomedical and Technological Strategies for the Control of Bacterial Growth and Infections. Biomedicines, 12(1), 176. https://doi.org/10.3390/biomedicines12010176