
Elevated NET, Calprotectin, and Neopterin Levels Discriminate between Disease Activity in COVID-19, as Evidenced by Need for Hospitalization among Patients in Northern Italy
 , ,
, ,  ,
,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. H3-NET Dual Hybrid ELISA
2.3. Calprotectin Mixed Monoclonal Assay
2.4. Competitive DNase ELISA
2.5. Neopterin
2.6. Complement Activation Products
2.7. Cytokine Detection
2.8. Statistics
3. Results
3.1. Patients
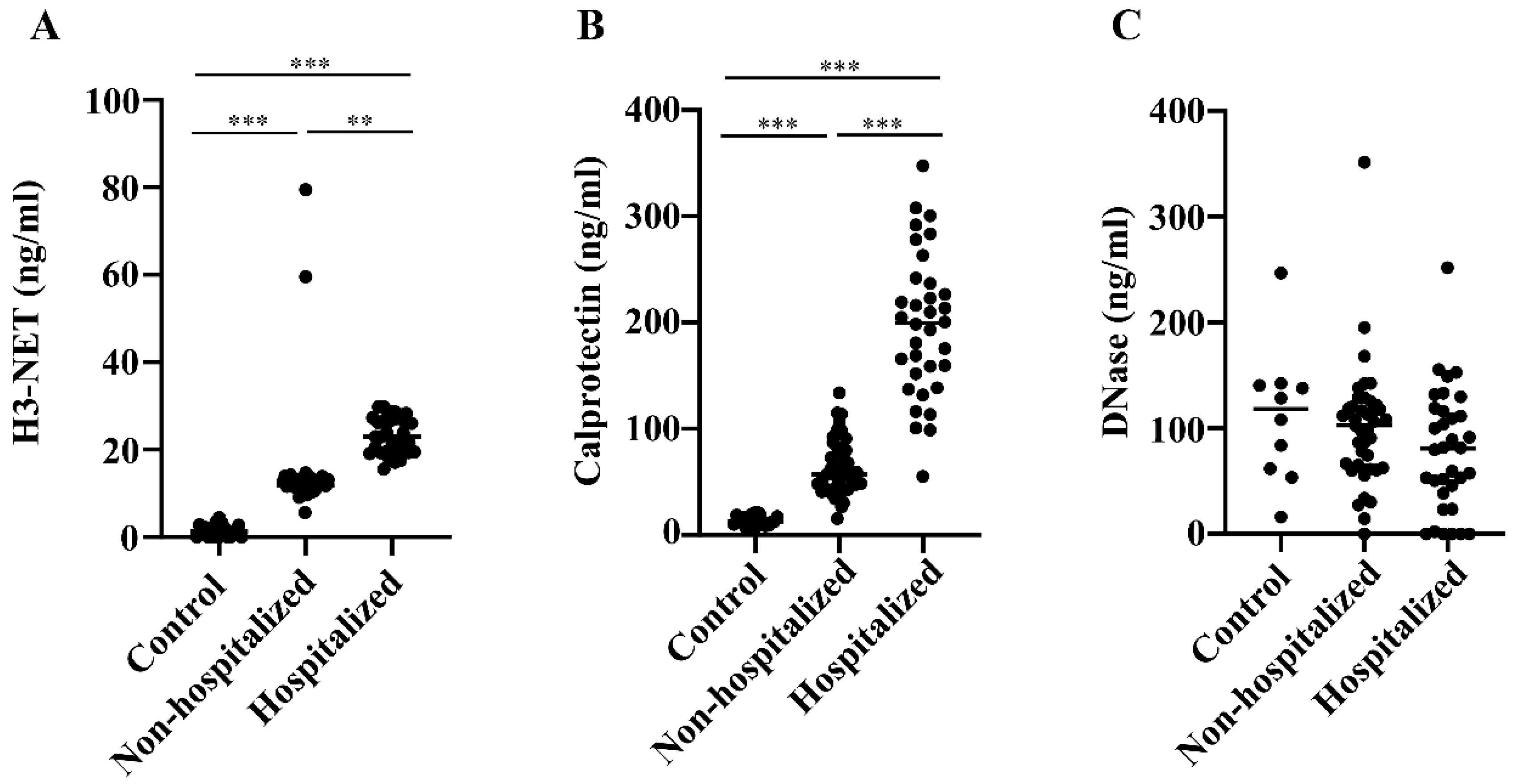
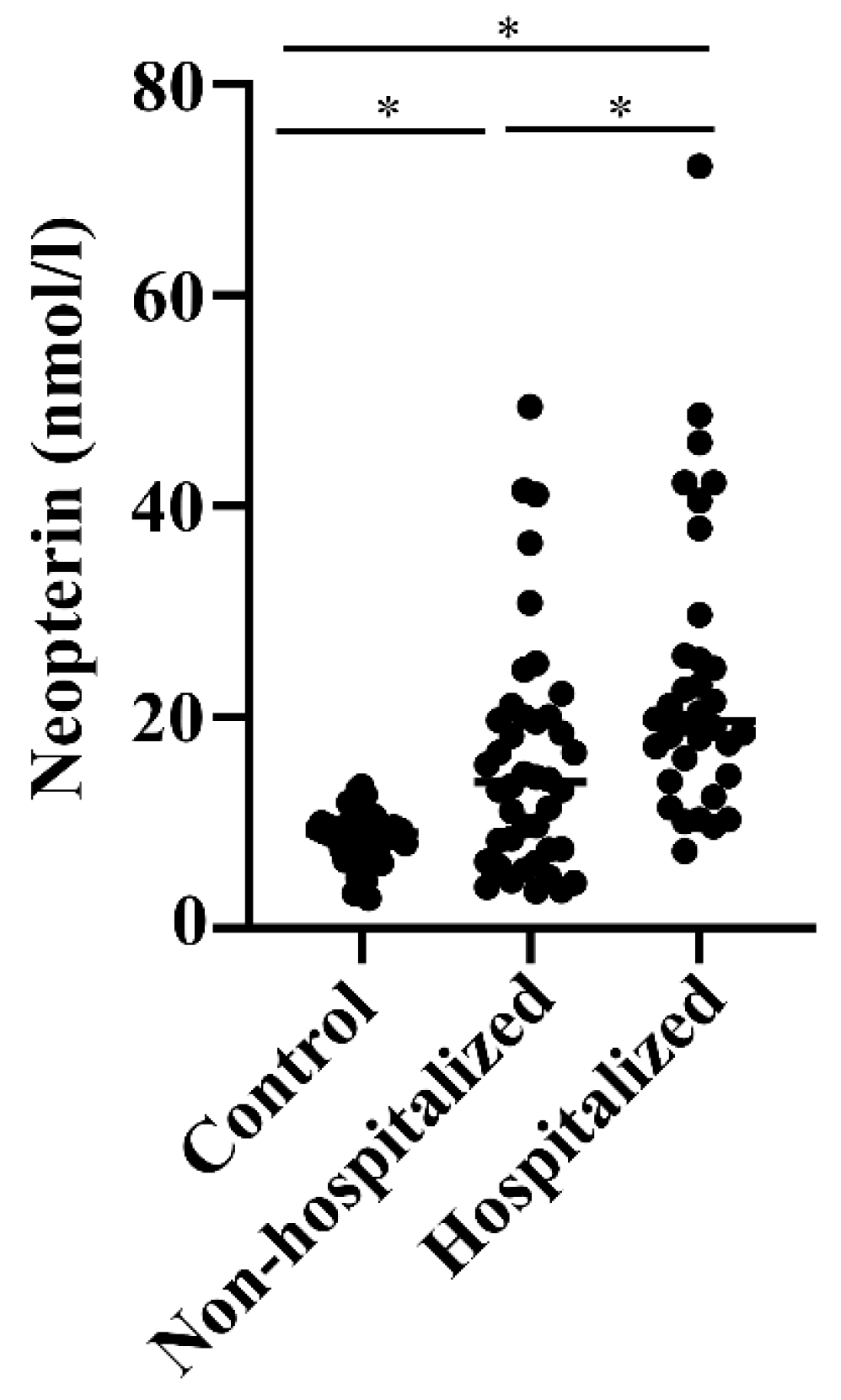
3.2. Levels of NETs, Calprotectin, DNase, and Neopterin in Hospitalized versus Non-Hospitalized COVID-19 Patients
3.3. Correlations between NETs, Calprotectin, Neopterin, and the Other Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Diamond, M.S.; Kanneganti, T.D. Innate immunity: The first line of defense against SARS-CoV-2. Nat. Immunol. 2022, 23, 165–176. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72,314 Cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Knight, J.S.; Caricchio, R.; Casanova, J.L.; Combes, A.J.; Diamond, B.; Fox, S.E.; Hanauer, D.A.; James, J.A.; Kanthi, Y.; Ladd, V.; et al. The intersection of COVID-19 and autoimmunity. J. Clin. Investig. 2021, 131, e154886. [Google Scholar] [CrossRef]
- Stephenson, E.; Reynolds, G.; Botting, R.A.; Calero-Nieto, F.J.; Morgan, M.D.; Tuong, Z.K.; Bach, K.; Sungnak, W.; Worlock, K.B.; Yoshida, M.; et al. Single-cell multi-omics analysis of the immune response in COVID-19. Nat. Med. 2021, 27, 904–916. [Google Scholar] [CrossRef]
- Pastorek, M.; Dúbrava, M.; Celec, P. On the Origin of Neutrophil Extracellular Traps in COVID-19. Front. Immunol. 2022, 13, 821007. [Google Scholar] [CrossRef]
- Morrissey, S.M.; Geller, A.E.; Hu, X.; Tieri, D.; Ding, C.; Klaes, C.K.; Cooke, E.A.; Woeste, M.R.; Martin, Z.C.; Chen, O.; et al. A specific low-density neutrophil population correlates with hypercoagulation and disease severity in hospitalized COVID-19 patients. JCI Insight 2021, 6, e148435. [Google Scholar] [CrossRef]
- Taylor, E.B. Casting a wide NET: An update on uncontrolled NETosis in response to COVID-19 infection. Clin. Sci. 2022, 136, 1047–1052. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, X.; Liu, X. NETosis and Neutrophil Extracellular Traps in COVID-19: Immunothrombosis and Beyond. Front. Immunol. 2022, 13, 838011. [Google Scholar] [CrossRef]
- Meroni, P.L.; Croci, S.; Lonati, P.A.; Pregnolato, F.; Spaggiari, L.; Besutti, G.; Bonacini, M.; Ferrigno, I.; Rossi, A.; Hetland, G.; et al. Complement activation predicts negative outcomes in COVID-19: The experience from Northen Italian patients. Autoimmun. Rev. 2023, 22, 103232. [Google Scholar] [CrossRef]
- Merad, M.; Blish, C.A.; Sallusto, F.; Iwasaki, A. The immunology and immunopathology of COVID-19. Science 2022, 375, 1122–1127. [Google Scholar] [CrossRef]
- Croce, K.; Gao, H.; Wang, Y.; Mooroka, T.; Sakuma, M.; Shi, C.; Sukhova, G.K.; Packard, R.R.; Hogg, N.; Libby, P.; et al. Myeloid-related protein-8/14 is critical for the biological response to vascular injury. Circulation 2009, 120, 427–436. [Google Scholar] [CrossRef]
- Jonsson, M.K.; Sundlisæter, N.P.; Nordal, H.H.; Hammer, H.B.; Aga, A.B.; Olsen, I.C.; Brokstad, K.A.; van der Heijde, D.; Kvien, T.K.; Fevang, B.S.; et al. Calprotectin as a marker of inflammation in patients with early rheumatoid arthritis. Ann. Rheum. Dis. 2017, 76, 2031–2037. [Google Scholar] [CrossRef]
- Mariani, A.; Marsili, M.; Nozzi, M.; Faricelli, R.; Chiarelli, F.; Breda, L. Serum calprotectin: Review of its usefulness and validity in paediatric rheumatic diseases. Clin. Exp. Rheumatol. 2015, 33, 109–114. [Google Scholar]
- Shi, H.; Zuo, Y.; Yalavarthi, S.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.; Woodward, W.; Lezak, S.P.; Lugogo, N.L.; et al. Neutrophil calprotectin identifies severe pulmonary disease in COVID-19. J. Leukoc. Biol. 2021, 109, 67–72. [Google Scholar] [CrossRef]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil extracellular traps contain calprotectin, a cytosolic protein complex involved in host defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef]
- Boneschansker, L.; Inoue, Y.; Oklu, R.; Irimia, D. Capillary plexuses are vulnerable to neutrophil extracellular traps. Integr. Biol. 2016, 8, 149–155. [Google Scholar] [CrossRef]
- Yang, J.; Wu, Z.; Long, Q.; Huang, J.; Hong, T.; Liu, W.; Lin, J. Insights into Immunothrombosis: The Interplay among Neutrophil Extracellular Trap, von Willebrand Factor, and ADAMTS13. Front. Immunol. 2020, 11, 610696. [Google Scholar] [CrossRef]
- Zuo, Y.; Zuo, M.; Yalavarthi, S.; Gockman, K.; Madison, J.A.; Shi, H.; Woodard, W.; Lezak, S.P.; Lugogo, N.L.; Knight, J.S.; et al. Neutrophil extracellular traps and thrombosis in COVID-19. J. Thromb. Thrombolysis 2021, 51, 446–453. [Google Scholar] [CrossRef]
- Jiménez-Alcázar, M.; Rangaswamy, C.; Panda, R.; Bitterling, J.; Simsek, Y.J.; Long, A.T.; Bilyy, R.; Krenn, V.; Renné, C.; Renné, T.; et al. Host DNases prevent vascular occlusion by neutrophil extracellular traps. Science 2017, 358, 1202–1206. [Google Scholar] [CrossRef]
- Hetland, G.; Fagerhol, M.K.; Dimova-Svetoslavova, V.P.; Mirlashari, M.R.; Nguyen, N.T.; Lind, A.; Kolset, S.O.; Søraas, A.V.L.; Nissen-Meyer, L.S.H. Inflammatory markers calprotectin, NETs, syndecan-1 and neopterin in COVID-19 convalescent blood donors. Scand. J. Clin. Lab. Investig. 2022, 82, 481–485. [Google Scholar] [CrossRef]
- Holm, S.; Kared, H.; Michelsen, A.E.; Kong, X.Y.; Dahl, T.B.; Schultz, N.H.; Nyman, T.A.; Fladeby, C.; Seljeflot, I.; Ueland, T.; et al. Immune complexes, innate immunity, and NETosis in ChAdOx1 vaccine-induced thrombocytopenia. Eur. Heart J. 2021, 42, 4064–4072. [Google Scholar] [CrossRef]
- Hirsch, J.; Uzun, G.; Zlamal, J.; Singh, A.; Bakchoul, T. Platelet-neutrophil interaction in COVID-19 and vaccine-induced thrombotic thrombocytopenia. Front. Immunol. 2023, 14, 1186000. [Google Scholar] [CrossRef]
- Leung, H.H.L.; Perdomo, J.; Ahmadi, Z.; Zheng, S.S.; Rashid, F.N.; Enjeti, A.; Ting, S.B.; Chong, J.J.H.; Chong, B.H. NETosis and thrombosis in vaccine-induced immune thrombotic thrombocytopenia. Nat. Commun. 2022, 13, 5206. [Google Scholar] [CrossRef]
- Fagerhol, M.K.; Schultz, N.H.; Mirlashari, M.R.; Wiedmann, M.K.H.; Nissen-Meyer, L.S.H.; Søraas, A.V.L.; Hetland, G. DNase analysed by a novel competitive assay in patients with complications after ChAdOx1 nCoV-19 vaccination and in normal unvaccinated blood donors. Scand. J. Immunol. 2023, 98, e13274. [Google Scholar] [CrossRef]
- Gieseg, S.P.; Baxter-Parker, G.; Lindsay, A. Neopterin, Inflammation, and Oxidative Stress: What Could We Be Missing? Antioxidants 2018, 7, 80. [Google Scholar] [CrossRef]
- Richardson, J.P.; Moyes, D.L.; Ho, J.; Naglik, J.R. Candida innate immunity at the mucosa. Semin. Cell Dev. Biol. 2019, 89, 58–70. [Google Scholar] [CrossRef]
- Nübling, C.M.; Chudy, M.; Volkers, P.; Löwer, J. Neopterin levels during the early phase of human immunodeficiency virus, hepatitis C virus, or hepatitis B virus infection. Transfusion 2006, 46, 1886–1891. [Google Scholar] [CrossRef]
- Mayersbach, P.; Fuchs, D.; Schennach, H. Performance of a fully automated quantitative neopterin measurement assay in a routine voluntary blood donation setting. Clin. Chem. Lab. Med. 2010, 48, 373–377. [Google Scholar] [CrossRef]
- Robertson, J.; Gostner, J.M.; Nilsson, S.; Andersson, L.M.; Fuchs, D.; Gisslen, M. Serum neopterin levels in relation to mild and severe COVID-19. BMC Infect. Dis. 2020, 20, 942. [Google Scholar] [CrossRef]
- Holter, J.C.; Pischke, S.E.; de Boer, E.; Lind, A.; Jenum, S.; Holten, A.R.; Tonby, K.; Barratt-Due, A.; Sokolova, M.; Schjalm, C.; et al. Systemic complement activation is associated with respiratory failure in COVID-19 hospitalized patients. Proc. Natl. Acad. Sci. USA 2020, 117, 25018–25025. [Google Scholar] [CrossRef]
- Carvelli, J.; Demaria, O.; Vély, F.; Batista, L.; Chouaki Benmansour, N.; Fares, J.; Carpentier, S.; Thibult, M.L.; Morel, A.; Remark, R.; et al. Association of COVID-19 inflammation with activation of the C5a-C5aR1 axis. Nature 2020, 588, 146–150. [Google Scholar] [CrossRef]
- Noris, M.; Benigni, A.; Remuzzi, G. The case of complement activation in COVID-19 multiorgan impact. Kidney Int. 2020, 98, 314–322. [Google Scholar] [CrossRef]
- Cugno, M.; Meroni, P.L.; Gualtierotti, R.; Griffini, S.; Grovetti, E.; Torri, A.; Lonati, P.; Grossi, C.; Borghi, M.O.; Novembrino, C.; et al. Complement activation and endothelial perturbation parallel COVID-19 severity and activity. J. Autoimmun. 2021, 116, 102560. [Google Scholar] [CrossRef]
- Besutti, G.; Ottone, M.; Fasano, T.; Pattacini, P.; Iotti, V.; Spaggiari, L.; Bonacini, R.; Nitrosi, A.; Bonelli, E.; Canovi, S.; et al. The value of computed tomography in assessing the risk of death in COVID-19 patients presenting to the emergency room. Eur. Radiol. 2021, 31, 9164–9175. [Google Scholar] [CrossRef]
- Galli, M.G.; Djuric, O.; Besutti, G.; Ottone, M.; Amidei, L.; Bitton, L.; Bonilauri, C.; Boracchia, L.; Campanale, S.; Curcio, V.; et al. Clinical and imaging characteristics of patients with COVID-19 predicting hospital readmission after emergency department discharge: A single-centre cohort study in Italy. BMJ Open 2022, 12, e052665. [Google Scholar] [CrossRef]
- Robinson, M.J.; Tessier, P.; Poulsom, R.; Hogg, N. The S100 family heterodimer, MRP-8/14, binds with high affinity to heparin and heparan sulfate glycosaminoglycans on endothelial cells. J. Biol. Chem. 2002, 277, 3658–3665. [Google Scholar] [CrossRef]
- Longstaff, C.; Varjú, I.; Sótonyi, P.; Szabó, L.; Krumrey, M.; Hoell, A.; Bóta, A.; Varga, Z.; Komorowicz, E.; Kolev, K. Mechanical stability and fibrinolytic resistance of clots containing fibrin, DNA, and histones. J. Biol. Chem. 2013, 288, 6946–6956. [Google Scholar] [CrossRef]
- Fagerhol, M.K.; Rugtveit, J. Heterogeneity of Fecal Calprotectin Reflecting Generation of Neutrophil Extracellular Traps (NETs) in the Gut: New Immunoassays Are Available. J. Mol. Pathol. 2022, 3, 38–51. [Google Scholar] [CrossRef]
- Macor, P.; Durigutto, P.; Mangogna, A.; Bussani, R.; De Maso, L.; D'Errico, S.; Zanon, M.; Pozzi, N.; Meroni, P.L.; Tedesco, F. Multiple-Organ Complement Deposition on Vascular Endothelium in COVID-19 Patients. Biomedicines 2021, 9, 1003. [Google Scholar] [CrossRef]
- Teijeira, A.; Garasa, S.; Ochoa, M.D.C.; Cirella, A.; Olivera, I.; Glez-Vaz, J.; Andueza, M.P.; Migueliz, I.; Alvarez, M.; Rodríguez-Ruiz, M.E.; et al. Differential Interleukin-8 thresholds for chemotaxis and netosis in human neutrophils. Eur. J. Immunol. 2021, 51, 2274–2280. [Google Scholar] [CrossRef]
- Bellmann-Weiler, R.; Lanser, L.; Burkert, F.; Seiwald, S.; Fritsche, G.; Wildner, S.; Schroll, A.; Koppelstätter, S.; Kurz, K.; Griesmacher, A.; et al. Neopterin Predicts Disease Severity in Hospitalized Patients with COVID-19. Open Forum Infect Dis. 2021, 8, ofaa521. [Google Scholar] [CrossRef]
- Ozger, H.S.; Dizbay, M.; Corbacioglu, S.K.; Aysert, P.; Demirbas, Z.; Tunccan, O.G.; Hizel, K.; Bozdayi, G.; Caglar, K. The prognostic role of neopterin in COVID-19 patients. J. Med. Virol. 2021, 93, 1520–1525. [Google Scholar] [CrossRef]
- Brambilla, M.; Canzano, P.; Becchetti, A.; Tremoli, E.; Camera, M. Alterations in platelets during SARS-CoV-2 infection. Platelets 2022, 33, 192–199. [Google Scholar] [CrossRef]
- Cervia-Hasler, C.; Brüningk, S.C.; Hoch, T.; Fan, B.; Muzio, G.; Thompson, R.C.; Ceglarek, L.; Meledin, R.; Westermann, P.; Emmenegger, M.; et al. Persistent complement dysregulation with signs of thromboinflammation in active Long Covid. Science 2024, 383, eadg7942. [Google Scholar] [CrossRef]
- Gallo Marin, B.; Aghagoli, G.; Lavine, K.; Yang, L.; Siff, E.J.; Chiang, S.S.; Salazar-Mather, T.P.; Dumenco, L.; Savaria, M.C.; Aung, S.N.; et al. Predictors of COVID-19 severity: A literature review. Rev. Med. Virol. 2021, 31, 1–10. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Parameter | NETs | Calprotectin | DNase | Neopterin |
|---|---|---|---|---|
| NETs | 1 | r = 0.3894 | r = −0.2628 | r = 0.1939 |
| p = 0.0005 | p = 0.0227 | p = 0.0932 | ||
| Calprotectin | - | 1 | r = −0.1703 | r = 0.2752 |
| - | p = 0.0161 | |||
| DNase | - | - | 1 | r = −0.1050 |
| - | ||||
| Neopterin | - | - | - | 1 |
| Parameter | Neopterin | Calprotectin | NET | DNase |
|---|---|---|---|---|
| CRP | 0.3328 * | - | - | −0.4026 ** |
| C5a | - | - | - | −0.3633 ** |
| SC5b-9 | - | 0.2635 * | - | - |
| BAFF | 0.5362 **** | 0.2472 ^ | - | −0.3422 ** |
| IFNa | 0.3480 * | 0.3061 * | - | - |
| IL-17A | - | - | - | 0.2619 * |
| VEGFR2 | - | - | - | 0.3595 ** |
| Procalcitonin | 0.5200 *** | 0.6032 *** | - | - |
| Ferritin | 0.5226 *** | 0.3895 ** | - | −0.3773 ** |
| Fibrinogen | - | 0.3895 * | - | - |
| AST | 0.6641 **** | 0.3562 ** | - | −0.3228 * |
| LDH | 0.5496 **** | 0.4021 ** | - | - |
| CPK | 0.4146 ** | 0.4439 ** | - | - |
| Troponin HS | 0.6490 **** | - | - | - |
| Leukos | - | 0.2500 ^ | - | - |
| Lymphos | - | −0.3020 * | - | - |
| Monoc | - | −0.3835 ** | - | - |
| NLR | - | - | 0.3737 ** | - |
| PLT | −0.4584 ** | - | −0.2737 * | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hetland, G.; Fagerhol, M.K.; Mirlashari, M.R.; Nissen-Meyer, L.S.H.; Croci, S.; Lonati, P.A.; Bonacini, M.; Salvarani, C.; Marvisi, C.; Bodio, C.; et al. Elevated NET, Calprotectin, and Neopterin Levels Discriminate between Disease Activity in COVID-19, as Evidenced by Need for Hospitalization among Patients in Northern Italy. Biomedicines 2024, 12, 766. https://doi.org/10.3390/biomedicines12040766
Hetland G, Fagerhol MK, Mirlashari MR, Nissen-Meyer LSH, Croci S, Lonati PA, Bonacini M, Salvarani C, Marvisi C, Bodio C, et al. Elevated NET, Calprotectin, and Neopterin Levels Discriminate between Disease Activity in COVID-19, as Evidenced by Need for Hospitalization among Patients in Northern Italy. Biomedicines. 2024; 12(4):766. https://doi.org/10.3390/biomedicines12040766
Chicago/Turabian StyleHetland, Geir, Magne Kristoffer Fagerhol, Mohammad Reza Mirlashari, Lise Sofie Haug Nissen-Meyer, Stefania Croci, Paola Adele Lonati, Martina Bonacini, Carlo Salvarani, Chiara Marvisi, Caterina Bodio, and et al. 2024. "Elevated NET, Calprotectin, and Neopterin Levels Discriminate between Disease Activity in COVID-19, as Evidenced by Need for Hospitalization among Patients in Northern Italy" Biomedicines 12, no. 4: 766. https://doi.org/10.3390/biomedicines12040766
APA StyleHetland, G., Fagerhol, M. K., Mirlashari, M. R., Nissen-Meyer, L. S. H., Croci, S., Lonati, P. A., Bonacini, M., Salvarani, C., Marvisi, C., Bodio, C., Muratore, F., Borghi, M. O., & Meroni, P. L. (2024). Elevated NET, Calprotectin, and Neopterin Levels Discriminate between Disease Activity in COVID-19, as Evidenced by Need for Hospitalization among Patients in Northern Italy. Biomedicines, 12(4), 766. https://doi.org/10.3390/biomedicines12040766