

PBMCs as Tool for Identification of Novel Immunotherapy Biomarkers in Lung Cancer
 , , , ,
, , , ,  , ,
, , 

 ,
, 
 ,
,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Treatment and Study Assessments
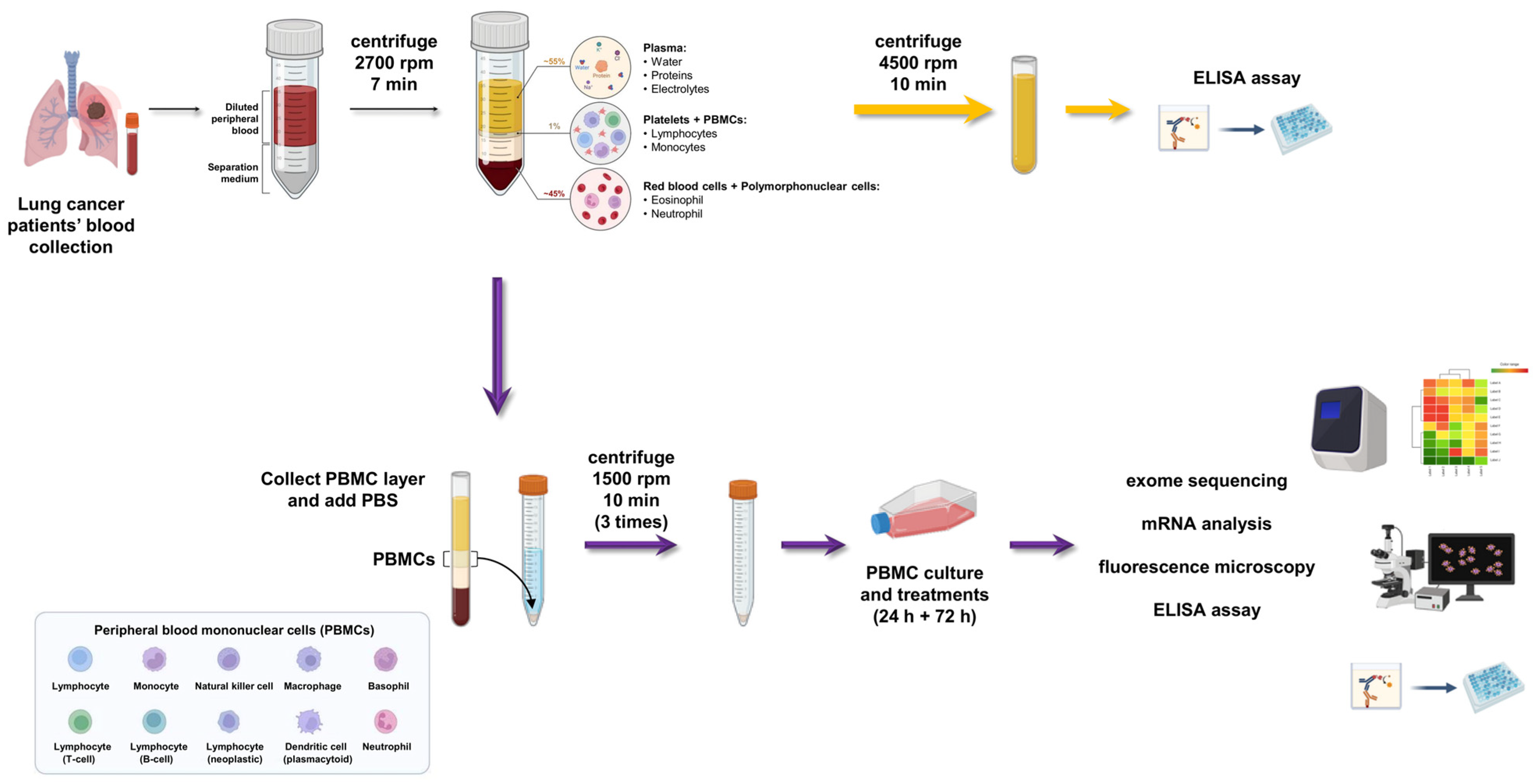
2.2. Peripheral Blood Mononuclear Cells (PBMC) Isolation: Step-By-Step Method
2.3. Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. RNA Extraction and cDNA Synthesis
2.5. Gene Expression Analysis by Quantitative qRT-PCR
2.6. Next-Generation Sequencing (NGS)
2.7. Generation of 3D Spheroid Culture
2.8. Co-Localization of 3D Tumor Spheroids and LC Patient-Derived PBMCs by Immunofluorescence (IF)
2.9. Flow Cytometry
2.10. Multiplex Cytokine Assay
2.11. Statistical Analysis
3. Results
3.1. Study Population, Patient Characteristics and Clinical Responses
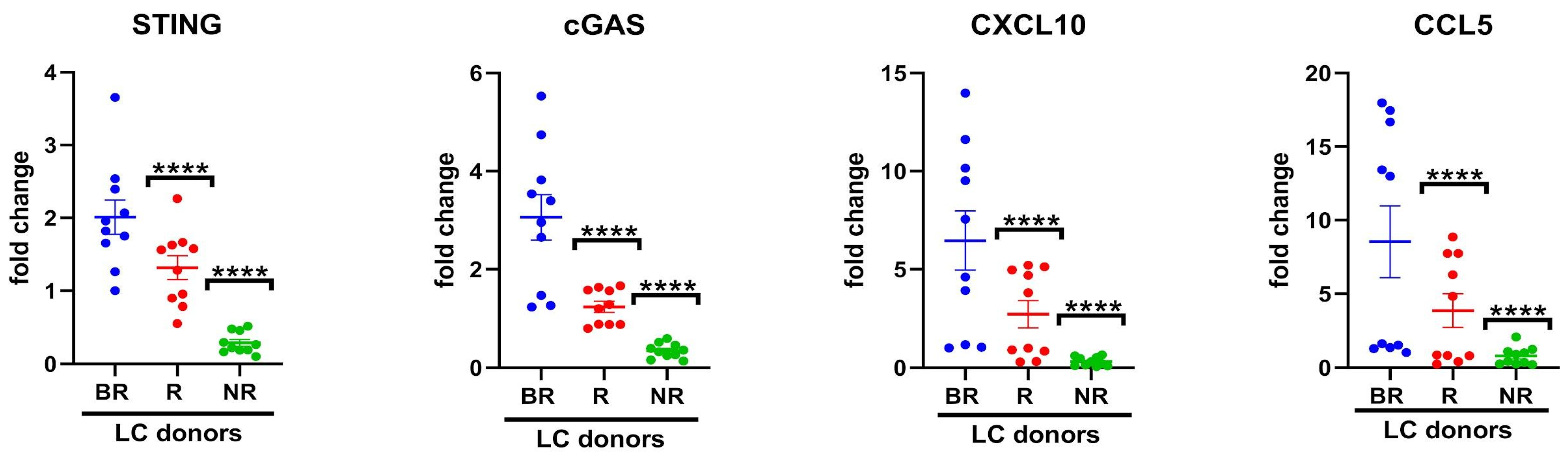
3.2. Association of Cytosolic DNA Sensor cGAS/STING Levels in Peripheral Blood with Response to Anti-PD-L1 Blockade
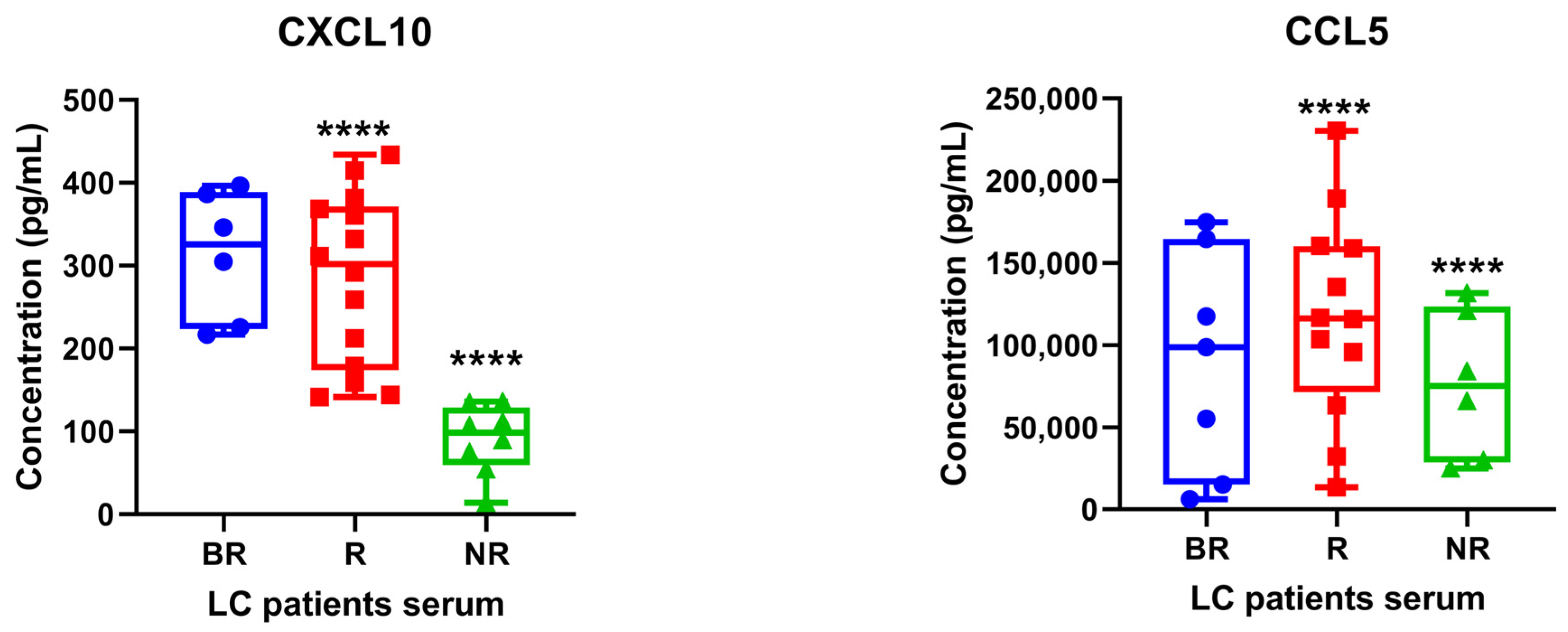
3.3. Determination of STING-Related Serum Cytokines in Patients with Lung Cancer under Anti-PD1/PD-L1 Therapy
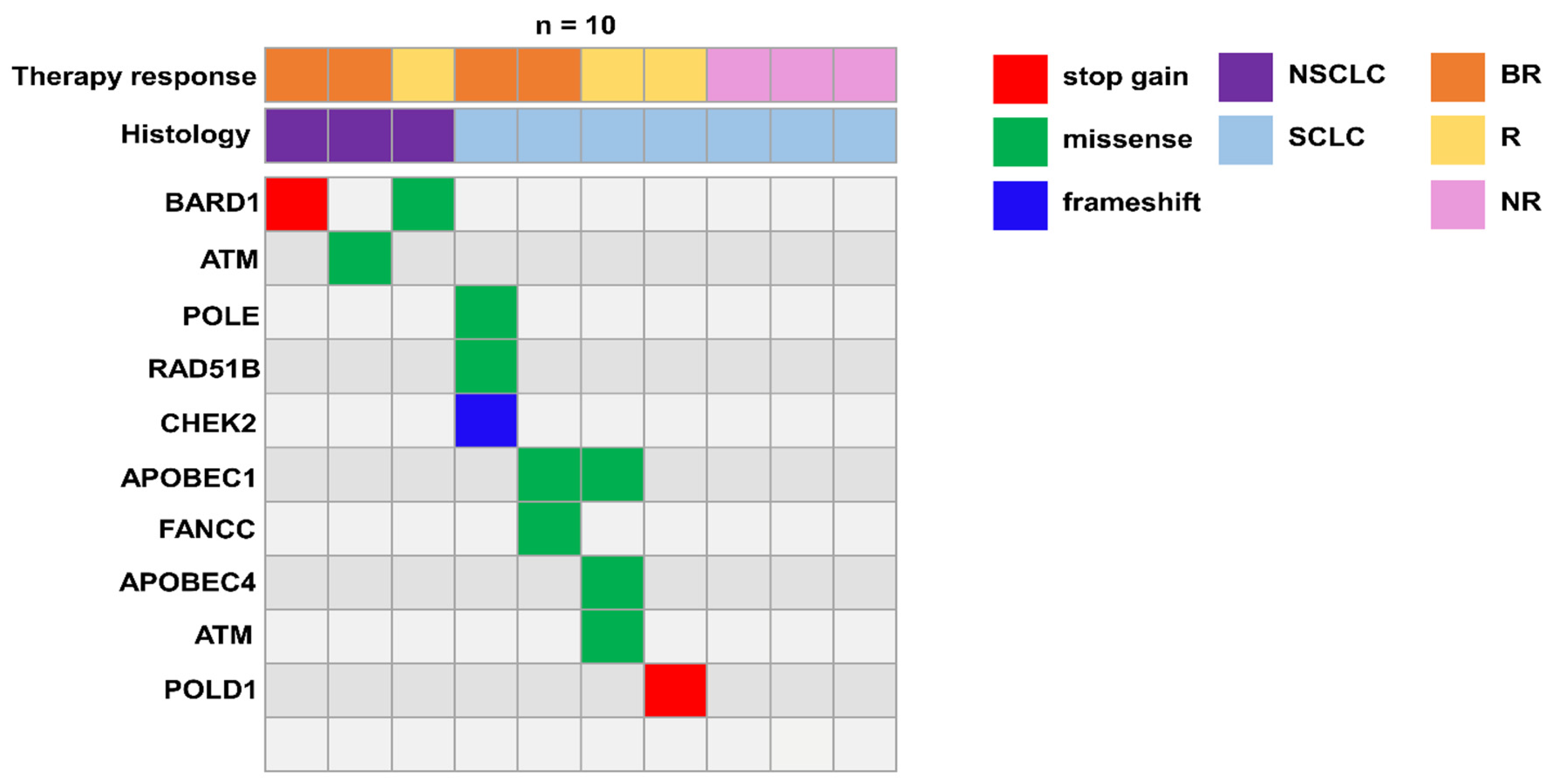
3.4. Germline Variants in DNA Damage Response (DDR) Genes Potentially Implicated in Immune Response
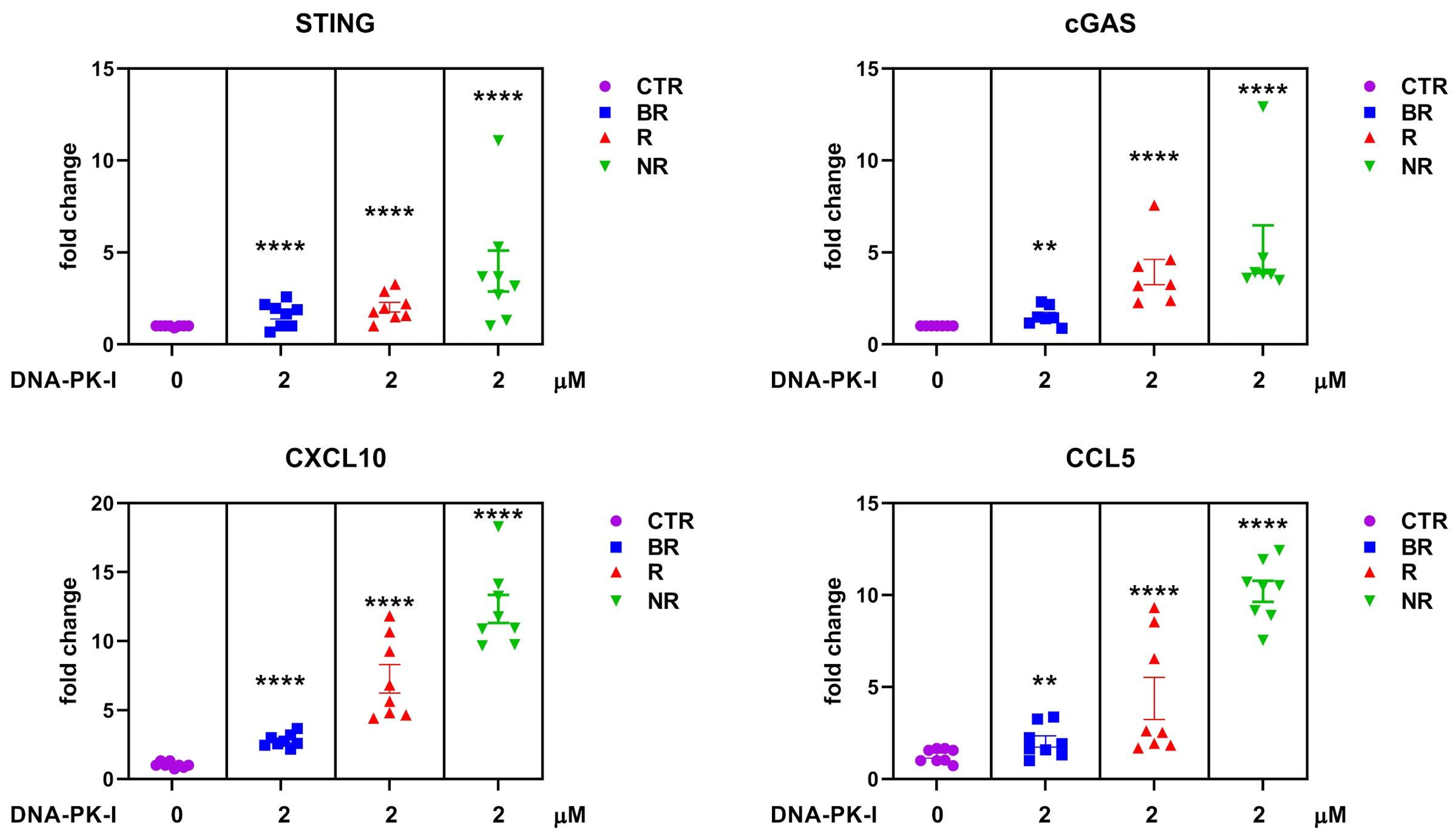
3.5. Effect of DNA-Dependent Protein Kinase (DNA-PK) Inhibitor on Cytosolic DNA Sensor cGAS/STING in Peripheral Blood of LC with Different Response to Anti-PD-L1 Blockade
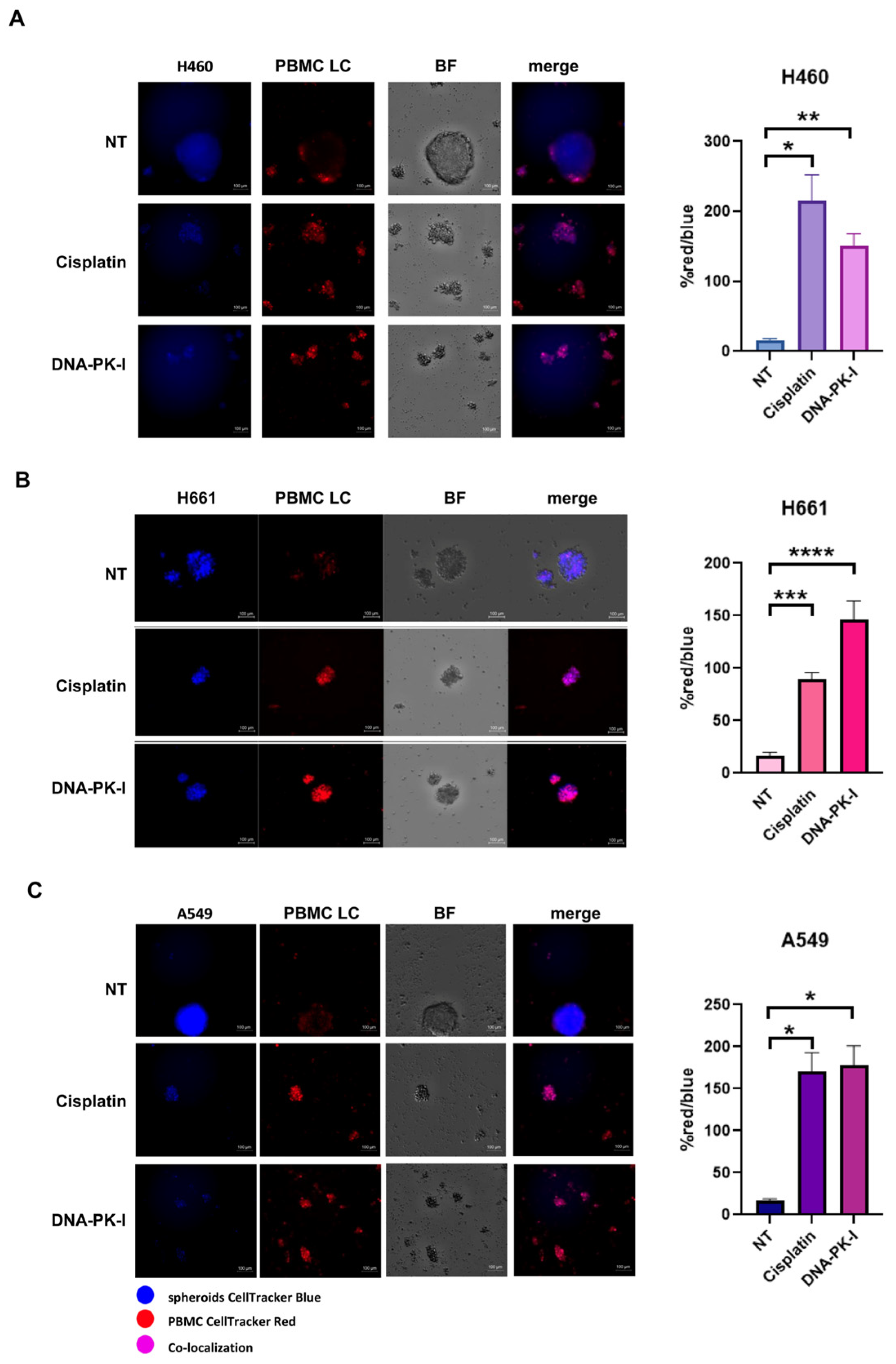
3.6. Antitumoral Activity and Innate Immune Cells Effect of DNA-PK Inhibitor
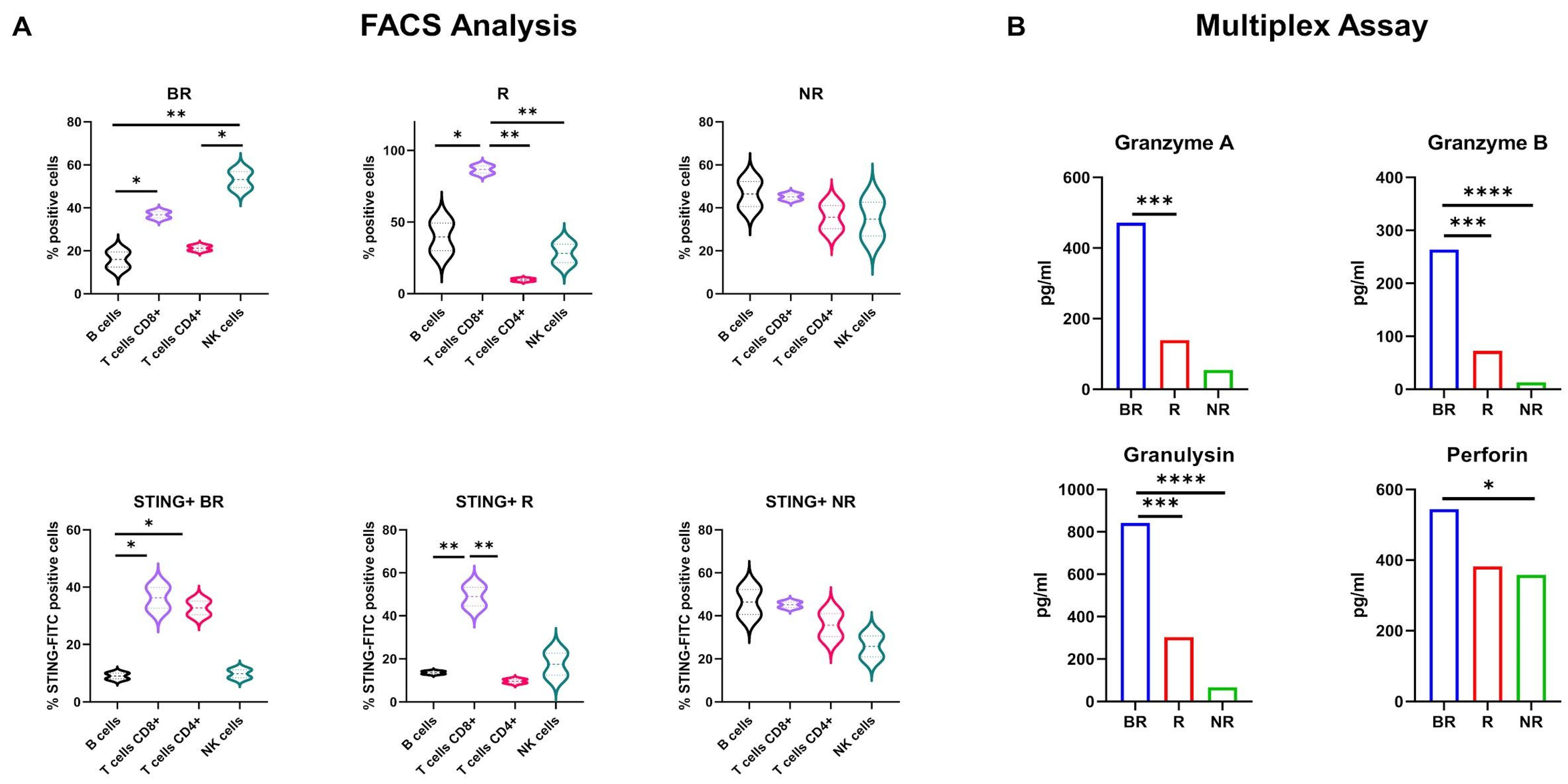
3.7. Subpopulation Analysis of STING Expression in LC Patient-Derived PBMCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lahiri, A.; Maji, A.; Potdar, P.D.; Singh, N.; Parikh, P.; Bisht, B.; Mukherjee, A.; Paul, M.K. Lung cancer immunotherapy: Progress, pitfalls, and promises. Mol. Cancer 2023, 22, 40. [Google Scholar] [CrossRef] [PubMed]
- Horvath, L.; Thienpont, B.; Zhao, L.; Wolf, D.; Pircher, A. Overcoming immunotherapy resistance in non-small cell lung cancer (NSCLC)—Novel approaches and future outlook. Mol. Cancer 2020, 19, 141. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Chen, M.; Zhang, R.; Zhang, W.; Wang, C.; Zhang, Y.; Wei, X.; Guan, Y.; Liu, J.; Feng, K.; et al. Manganese is critical for antitumor immune responses via cGAS-STING and improves the efficacy of clinical immunotherapy. Cell Res. 2020, 30, 966–979. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.W.; Higashi, R.M.; Song, H.; Daneshmandi, S.; Mahan, A.L.; Purdom, M.S.; Bocklage, T.J.; Pittman, T.A.; He, D.; Wang, C.; et al. Innate immune activation by checkpoint inhibition in human patient-derived lung cancer tissues. eLife 2021, 10, e69578. [Google Scholar] [CrossRef] [PubMed]
- Della Corte, C.M.; Sen, T.; Gay, C.M.; Ramkumar, K.; Diao, L.; Cardnell, R.J.; Rodriguez, B.L.; Stewart, C.A.; Papadimitrakopoulou, V.A.; Gibson, L.; et al. STING Pathway Expression Identifies NSCLC with an Immune-Responsive Phenotype. J. Thorac. Oncol. 2020, 15, 777–791. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Park, Y.S.; Lee, H.W.; Park, T.Y.; Lee, J.K.; Heo, E.Y. Peripheral lymphocyte count as a surrogate marker of immune checkpoint inhibitor therapy outcomes in patients with non-small-cell lung cancer. Sci. Rep. 2022, 12, 626. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Cho, J.; Ku, B.M.; Koh, J.; Sun, J.M.; Lee, S.H.; Ahn, J.S.; Cheon, J.; Min, Y.J.; Park, S.H.; et al. The First-week Proliferative Response of Peripheral Blood PD-1. Clin. Cancer Res. 2019, 25, 2144–2154. [Google Scholar] [CrossRef] [PubMed]
- Della Corte, C.M.; Fasano, M.; Ciaramella, V.; Cimmino, F.; Cardnell, R.; Gay, C.M.; Ramkumar, K.; Diao, L.; Di Liello, R.; Viscardi, G.; et al. Anti-tumor activity of cetuximab plus avelumab in non-small cell lung cancer patients involves innate immunity activation: Findings from the CAVE-Lung trial. J. Exp. Clin. Cancer Res. 2022, 41, 109. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S.; et al. Pembrolizumab versus Chemotherapy for PD-L1-Positive Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef]
- Novello, S.; Kowalski, D.M.; Luft, A.; Gümüş, M.; Vicente, D.; Mazières, J.; Rodríguez-Cid, J.; Tafreshi, A.; Cheng, Y.; Lee, K.H.; et al. Pembrolizumab Plus Chemotherapy in Squamous Non-Small-Cell Lung Cancer: 5-Year Update of the Phase III KEYNOTE-407 Study. J. Clin. Oncol. 2023, 41, 1999–2006. [Google Scholar] [CrossRef]
- Horn, L.; Mansfield, A.S.; Szczęsna, A.; Havel, L.; Krzakowski, M.; Hochmair, M.J.; Huemer, F.; Losonczy, G.; Johnson, M.L.; Nishio, M.; et al. First-Line Atezolizumab plus Chemotherapy in Extensive-Stage Small-Cell Lung Cancer. N. Engl. J. Med. 2018, 379, 2220–2229. [Google Scholar] [CrossRef] [PubMed]
- Della Corte, C.M.; Barra, G.; Ciaramella, V.; Di Liello, R.; Vicidomini, G.; Zappavigna, S.; Luce, A.; Abate, M.; Fiorelli, A.; Caraglia, M.; et al. Antitumor activity of dual blockade of PD-L1 and MEK in NSCLC patients derived three-dimensional spheroid cultures. J. Exp. Clin. Cancer Res. 2019, 38, 253. [Google Scholar] [CrossRef] [PubMed]
- Fasano, M.; Della Corte, C.M.; Di Liello, R.; Barra, G.; Sparano, F.; Viscardi, G.; Iacovino, M.L.; Paragliola, F.; Famiglietti, V.; Ciaramella, V.; et al. Induction of natural killer antibody-dependent cell cytotoxicity and of clinical activity of cetuximab plus avelumab in non-small cell lung cancer. ESMO Open 2020, 5, e000753. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, H.; Caeser, R.; Chavan, S.S.; Zhan, Y.A.; Chow, A.; Manoj, P.; Uddin, F.; Kitai, H.; Qu, R.; Hayatt, O.; et al. WEE1 inhibition enhances the antitumor immune response to PD-L1 blockade by the concomitant activation of STING and STAT1 pathways in SCLC. Cell Rep. 2022, 39, 110814. [Google Scholar] [CrossRef]
- Schindler, H.; Lusky, F.; Daniello, L.; Elshiaty, M.; Gaissmaier, L.; Benesova, K.; Souto-Carneiro, M.; Angeles, A.K.; Janke, F.; Eichhorn, F.; et al. Serum cytokines predict efficacy and toxicity, but are not useful for disease monitoring in lung cancer treated with PD-(L)1 inhibitors. Front. Oncol. 2022, 12, 1010660. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Xi, Y.; Yuan, Z.; Wang, B.; Hu, S.; Fang, C.; Cai, Y.; Fu, X.; Li, L. IFN-γ activates the tumor cell-intrinsic STING pathway through the induction of DNA damage and cytosolic dsDNA formation. Oncoimmunology 2022, 11, 2044103. [Google Scholar] [CrossRef] [PubMed]
- Sorscher, S.; LoPiccolo, J.; Heald, B.; Chen, E.; Bristow, S.L.; Michalski, S.T.; Nielsen, S.M.; Lacoste, A.; Keyder, E.; Lee, H.; et al. Rate of Pathogenic Germline Variants in Patients with Lung Cancer. JCO Precis. Oncol. 2023, 7, e2300190. [Google Scholar] [CrossRef] [PubMed]
- Garmezy, B.; Gheeya, J.; Lin, H.Y.; Huang, Y.; Kim, T.; Jiang, X.; Thein, K.Z.; Pilié, P.G.; Zeineddine, F.; Wang, W.; et al. Clinical and Molecular Characterization of. JCO Precis. Oncol. 2022, 6, e2100267. [Google Scholar] [CrossRef] [PubMed]
- Pecori, R.; Di Giorgio, S.; Paulo Lorenzo, J.; Nina Papavasiliou, F. Functions and consequences of AID/APOBEC-mediated DNA and RNA deamination. Nat. Rev. Genet. 2022, 23, 505–518. [Google Scholar] [CrossRef]
- Guerreiro, I.M.; Barros-Silva, D.; Lopes, P.; Cantante, M.; Cunha, A.L.; Lobo, J.; Antunes, L.; Rodrigues, A.; Soares, M.; Henrique, R.; et al. Levels as a Potential Predictive Biomarker for PD-1 Blockade Response in Non-Small Cell Lung Cancer. J. Clin. Med. 2020, 9, 1000. [Google Scholar] [CrossRef]
- Yi, R.; Lin, A.; Cao, M.; Xu, A.; Luo, P.; Zhang, J. ATM Mutations Benefit Bladder Cancer Patients Treated with Immune Checkpoint Inhibitors by Acting on the Tumor Immune Microenvironment. Front. Genet. 2020, 11, 933. [Google Scholar] [CrossRef] [PubMed]
- Concannon, K.; Morris, B.B.; Gay, C.M.; Byers, L.A. Combining targeted DNA repair inhibition and immune-oncology approaches for enhanced tumor control. Mol. Cell 2023, 83, 660–680. [Google Scholar] [CrossRef] [PubMed]
- Pardo, J.; Aguilo, J.I.; Anel, A.; Martin, P.; Joeckel, L.; Borner, C.; Wallich, R.; Müllbacher, A.; Froelich, C.J.; Simon, M.M. The biology of cytotoxic cell granule exocytosis pathway: Granzymes have evolved to induce cell death and inflammation. Microbes Infect. 2009, 11, 452–459. [Google Scholar] [CrossRef]
- Boivin, W.A.; Cooper, D.M.; Hiebert, P.R.; Granville, D.J. Intracellular versus extracellular granzyme B in immunity and disease: Challenging the dogma. Lab. Investig. 2009, 89, 1195–1220. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.T.; Fang, Y.; Wei, Q.; Shi, H.; Tan, H.; Deng, Y.; Zeng, Z.; Qiu, J.; Chen, C.; Sun, L.; et al. Tumor-targeted delivery of a STING agonist improves cancer immunotherapy. Proc. Natl. Acad. Sci. USA 2022, 119, e2214278119. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.C.; Chiang, S.F.; Chang, H.Y.; Chen, W.T.; Yang, P.C.; Chen, T.W.; Liang, J.A.; Shiau, A.C.; Ke, T.W.; Clifford Chao, K.S. Engineered sTRAIL-armed MSCs overcome STING deficiency to enhance the therapeutic efficacy of radiotherapy for immune checkpoint blockade. Cell Death Dis. 2022, 13, 610. [Google Scholar] [CrossRef]
- Byers, L.A.; Wang, J.; Nilsson, M.B.; Fujimoto, J.; Saintigny, P.; Yordy, J.; Giri, U.; Peyton, M.; Fan, Y.H.; Diao, L.; et al. Proteomic profiling identifies dysregulated pathways in small cell lung cancer and novel therapeutic targets including PARP1. Cancer Discov. 2012, 2, 798–811. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Sequence | Reverse Sequence |
|---|---|---|
| 18S | 5′-CGCCGCTAGAGGTGAAATTC-3′ | 3′-CTTTCGCTCTGGTCCGTCTT-5′ |
| STING | 5′-CAGGCACTGAACATCCTCCT-3′ | 3′-ATATACAGCCGCTGGCTCAC-5′ |
| cGAS | 5′-CTCCACGAAGCCAAGACCTC-3′ | 3′-GCGGCTGAGCTTCAACTTCT-5′ |
| CCL5 | 5′-CTCCCCATATTCCTCGGACA-3′ | 3′-CTCTGGGTTGGCACACACTT-5′ |
| CXCL10 | 5′-GGTGAGAAGAGATGTCTGAATCC-3 | 3′-GTCCATCCTTGGAAGCACTGCA-5′ |
| 1L-IO (n = 3) | ICT (n = 25) | |
|---|---|---|
| Samples (n) | ||
| BR R NR | 2 1 - | 5 12 8 |
| Age (mean, range) | 66.00 (60–75) | 66.68 (52–83) |
| Sex (n, %) | ||
| Female Male | - 3 (100.00%) | 6 (24.00%) 19 (76.00%) |
| Histology (n, %) | ||
| SCLC NSCLC | - 3 (100.00%) | 15 (60.00%) 10 (40.00%) |
| BR | ||||||
|---|---|---|---|---|---|---|
| STING vs. CXCL10 | STING vs. CCL5 | STING vs. cGAS | cGAS vs. CXCL10 | cGAS vs. CCL5 | CXCL10 vs. CCL5 | |
| r value | 0.6705 | 0.5582 | 0.3118 | 0.4053 | 0.1737 | 0.8777 |
| 95% CI | 0.07070 to 0.9142 | −0.1102 to 0.8789 | −0.3955 to 0.7869 | −0.3012 to 0.8245 | −0.5119 to 0.7241 | 0.5545 to 0.9708 |
| p value | 0.0339 | 0.0936 | 0.3805 | 0.2452 | 0.6314 | 0.0008 |
| R | ||||||
| STING vs. CXCL10 | STING vs. CCL5 | STING vs. cGAS | cGAS vs. CXCL10 | cGAS vs. CCL5 | CXCL10 vs. CCL5 | |
| r value | 0.4070 | 0.4382 | 0.7053 | 0.8643 | 0.8796 | 0.9051 |
| 95% CI | −0.2994 to 0.8252 | −0.2643 to 0.8369 | 0.1362 to 0.9244 | 0.5149 to 0.9675 | 0.5602 to 0.9713 | 0.6405 to 0.9776 |
| p value | 0.2431 | 0.2052 | 0.0227 | 0.0013 | 0.0008 | 0.0003 |
| NR | ||||||
| STING vs. CXCL10 | STING vs. CCL5 | STING vs. cGAS | cGAS vs. CXCL10 | cGAS vs. CCL5 | CXCL10 vs. CCL5 | |
| r value | −0.1854 | 0.1827 | 0.3602 | 0.08317 | 0.5195 | −0.4806 |
| 95% CI | −0.7298 to 0.5030 | −0.5050 to 0.7285 | −0.3485 to 0.8068 | −0.5767 to 0.6773 | −0.1637 to 0.8659 | −0.8523 to 0.2137 |
| p value | 0.6082 | 0.6134 | 0.3066 | 0.8193 | 0.1238 | 0.1598 |
| Patient ID | Genes | HGVSP | Consequence | Sift Prediction | PolyPhen Prediction | Variant Read Frequency | ClinVar |
|---|---|---|---|---|---|---|---|
| NSCLC_1 | BARD1 | p.(Arg406Ter) | stop_gained | - | - | 0.48421052 | VCV000229677.13:pathogenic |
| NSCLC_2 | ATM | p.(Ser333Phe) | missense_variant | deleterious | probably damaging | 0.47340426 | VCV000127471.16:uncertain significance |
| NSCLC_3 | BARD1 | p.(Asp673Tyr) | missense_variant | deleterious | probably damaging | 0.55279505 | VCV000482778.3:uncertain significance |
| SCLC_1 | CHEK2 POLE RAD51B | p.(Asn105IlefsTer11) p.(Lys425Arg) p.(Lys243Arg) | frameshift_variant missense_variant missense_variant | - deleterious deleterious | - probably damaging probably damaging | 0.19469027 0.46486485 0.16216215 | - VCV000224587.7:uncertain significance - |
| SCLC_2 | APOBEC1 FANCC | p.(Glu41Gln) p.(Asp195Val) | missense_variant missense_variant | deleterious deleterious | - probably damaging | 0.53571427 0.556701 | - VCV000134305.15:uncertain significance |
| SCLC_3 | APOBEC4 ATM APOBEC1 | p.(Arg183Trp) p.(Ser49Cys) p.(Pro108Ser) | missense_variant missense_variant missense_variant | tolerated deleterious deleterious | - possibly damaging - | 0.43418014 0.5242967 0.50517243 | - VCV000003048.16:uncertain significance - |
| SCLC_4 | POLD1 | p.(Gln53Ter) | stop_gained | - | - | 0.44516128 | - |
| SCLC_5 | - | - | - | - | - | - | - |
| SCLC_6 | - | - | - | - | - | - | - |
| SCLC_7 | - | - | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Rosa, C.; Iommelli, F.; De Rosa, V.; Ercolano, G.; Sodano, F.; Tuccillo, C.; Amato, L.; Tirino, V.; Ariano, A.; Cimmino, F.; et al. PBMCs as Tool for Identification of Novel Immunotherapy Biomarkers in Lung Cancer. Biomedicines 2024, 12, 809. https://doi.org/10.3390/biomedicines12040809
De Rosa C, Iommelli F, De Rosa V, Ercolano G, Sodano F, Tuccillo C, Amato L, Tirino V, Ariano A, Cimmino F, et al. PBMCs as Tool for Identification of Novel Immunotherapy Biomarkers in Lung Cancer. Biomedicines. 2024; 12(4):809. https://doi.org/10.3390/biomedicines12040809
Chicago/Turabian StyleDe Rosa, Caterina, Francesca Iommelli, Viviana De Rosa, Giuseppe Ercolano, Federica Sodano, Concetta Tuccillo, Luisa Amato, Virginia Tirino, Annalisa Ariano, Flora Cimmino, and et al. 2024. "PBMCs as Tool for Identification of Novel Immunotherapy Biomarkers in Lung Cancer" Biomedicines 12, no. 4: 809. https://doi.org/10.3390/biomedicines12040809
APA StyleDe Rosa, C., Iommelli, F., De Rosa, V., Ercolano, G., Sodano, F., Tuccillo, C., Amato, L., Tirino, V., Ariano, A., Cimmino, F., di Guida, G., Filosa, G., di Liello, A., Ciardiello, D., Martinelli, E., Troiani, T., Napolitano, S., Martini, G., Ciardiello, F., ... Della Corte, C. M. (2024). PBMCs as Tool for Identification of Novel Immunotherapy Biomarkers in Lung Cancer. Biomedicines, 12(4), 809. https://doi.org/10.3390/biomedicines12040809