
HGF Modulates Actin Cytoskeleton Remodeling and Contraction in Testicular Myoid Cells

 ,
, 


Abstract
:
1. Introduction
2. Results
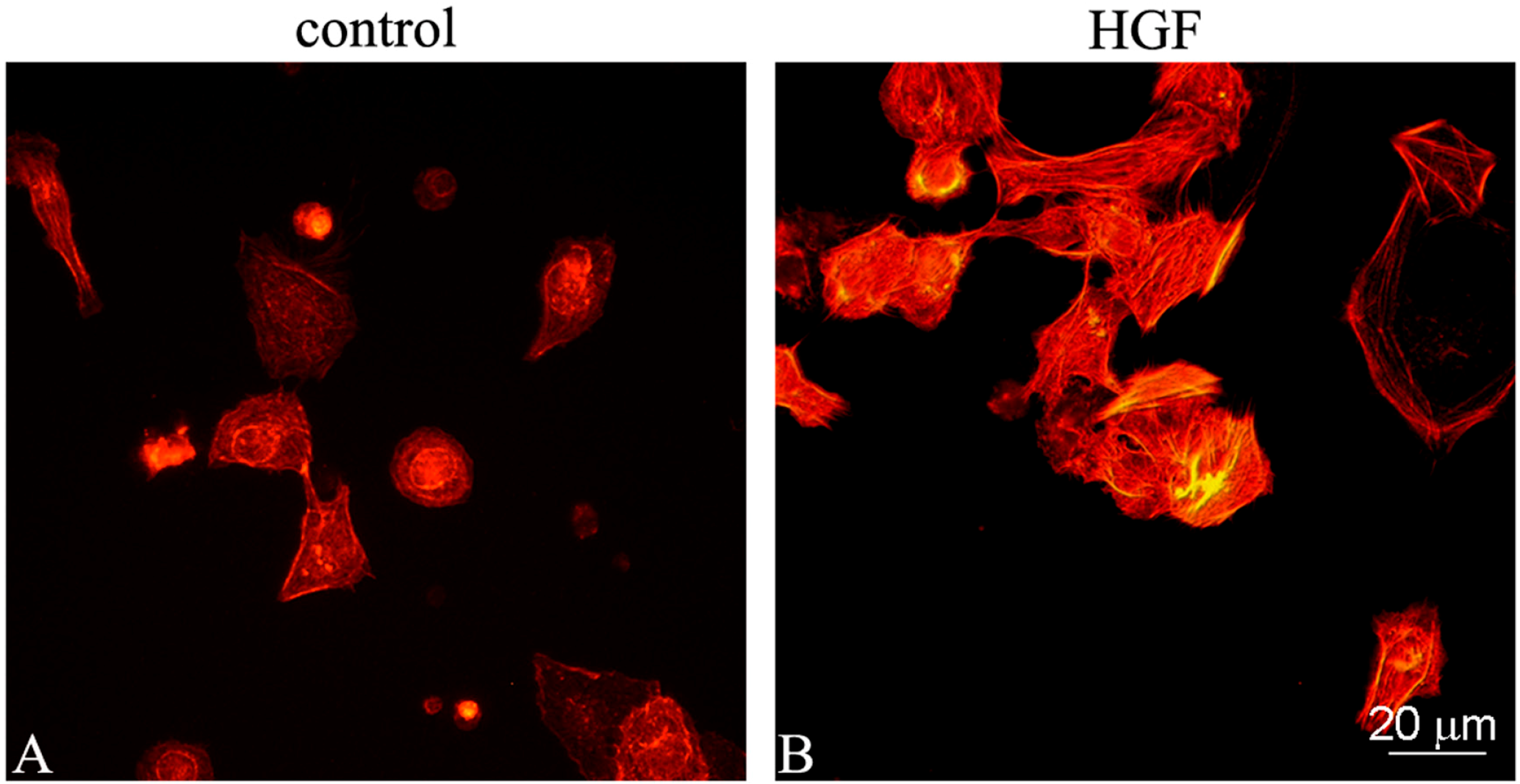
2.1. Effect of HGF on Cell Shape and the Actin Cytoskeleton of Cultured Myoid Cells

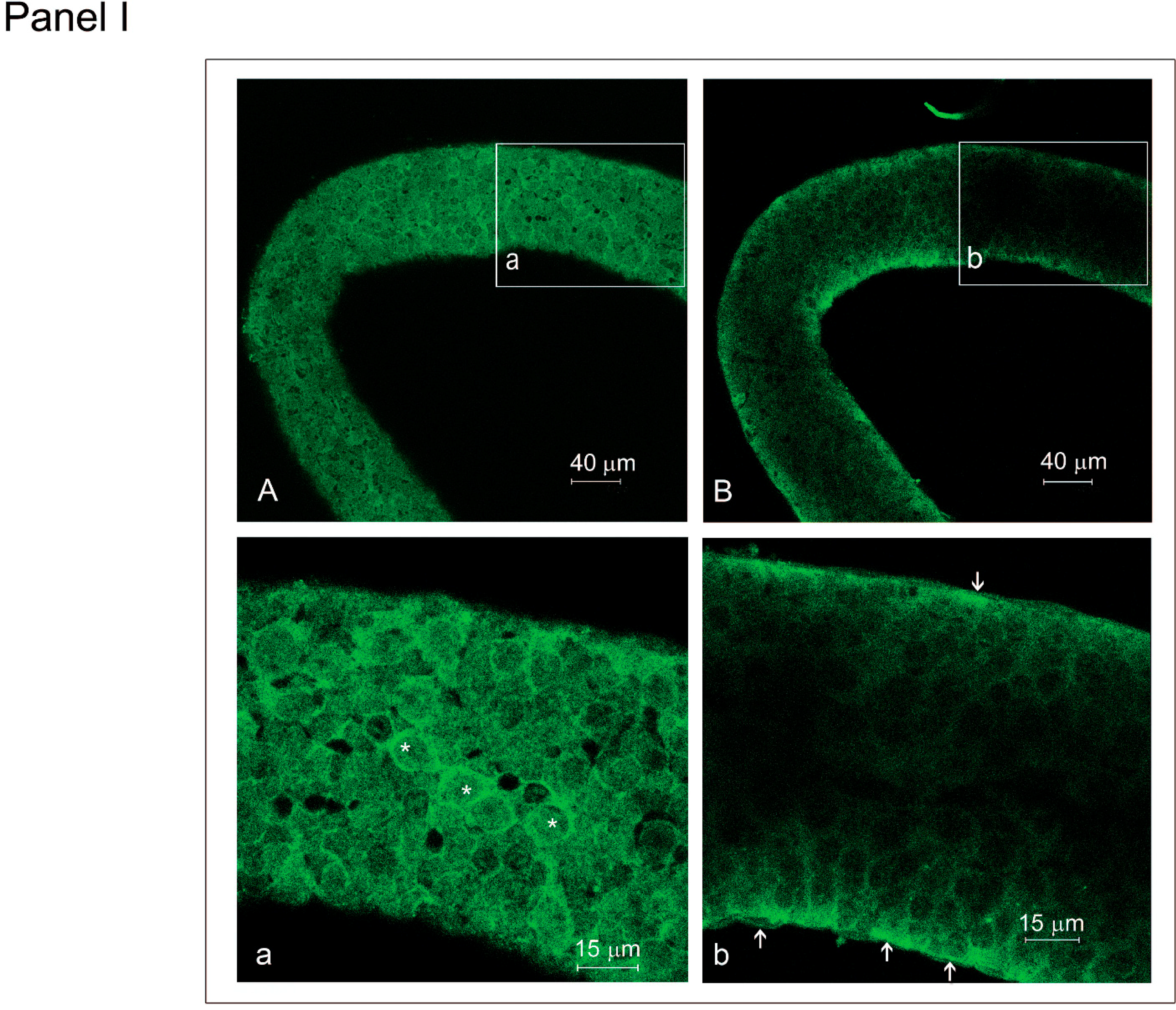
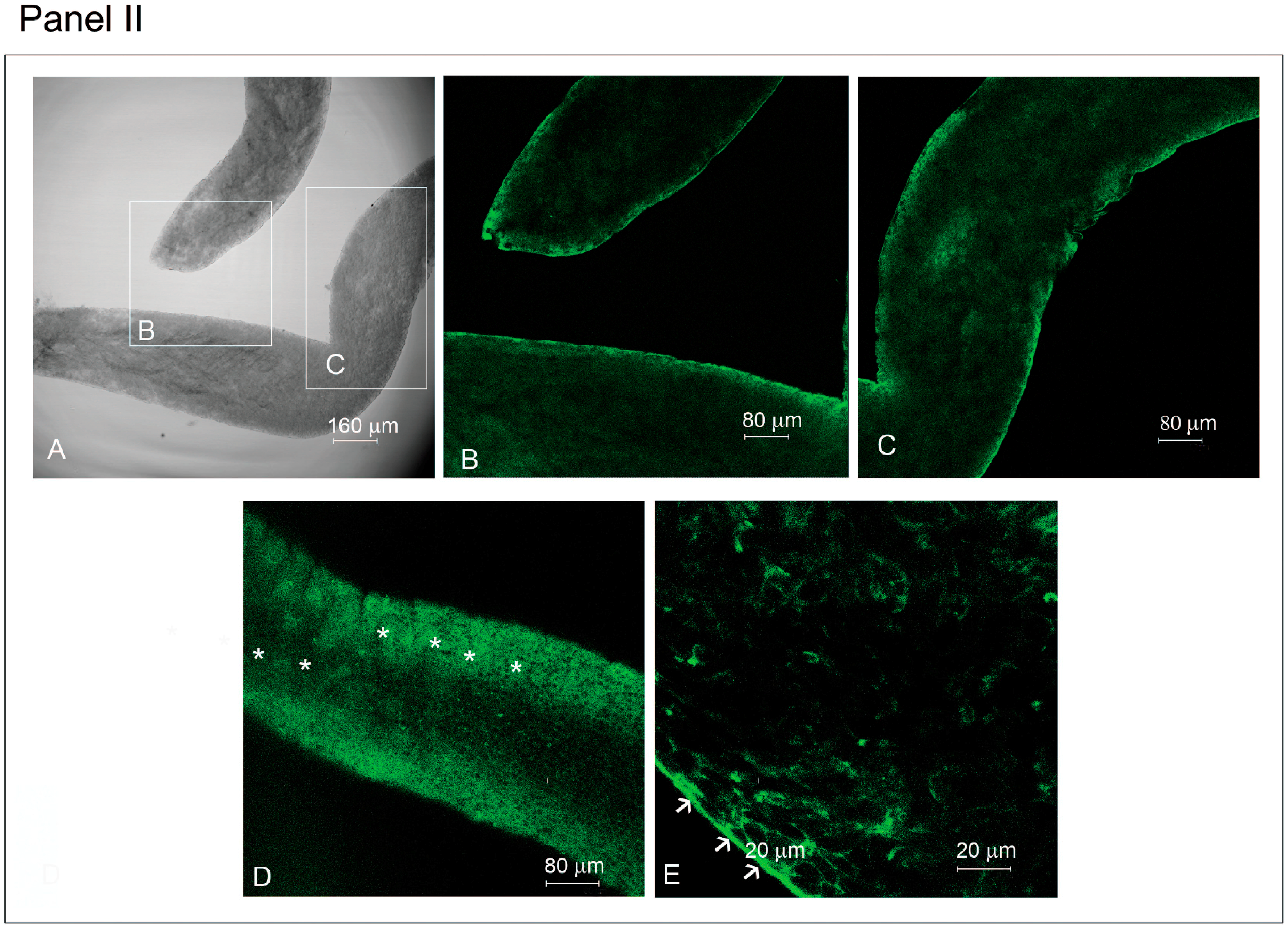
2.2. Effect of HGF on Myoid Cells Organized in Isolated Seminiferous Tubules



2.3. Presence of MET


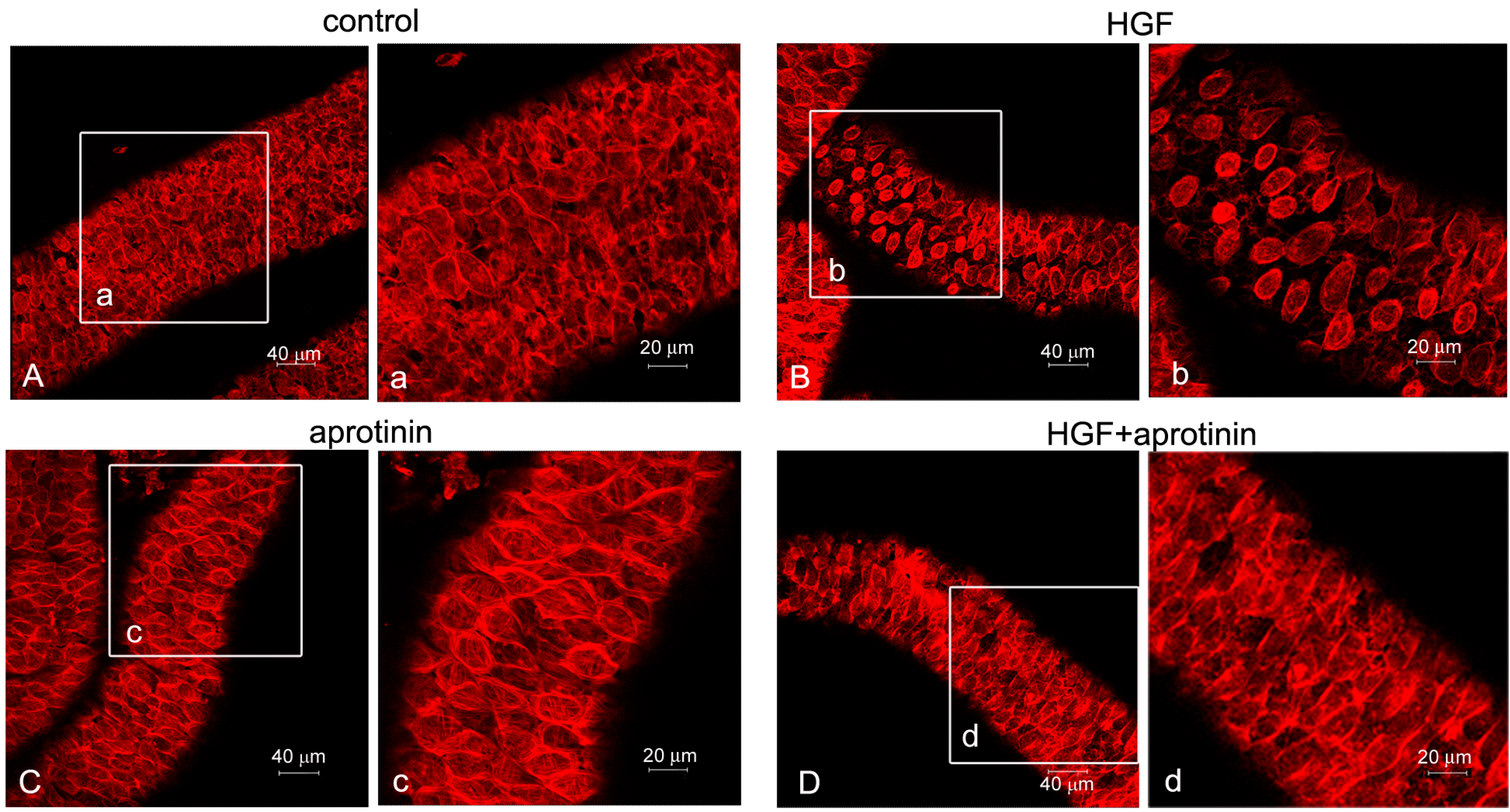
2.4. Effect of Inhibition of PA Activity on Actin Cytoskeleton Remodeling
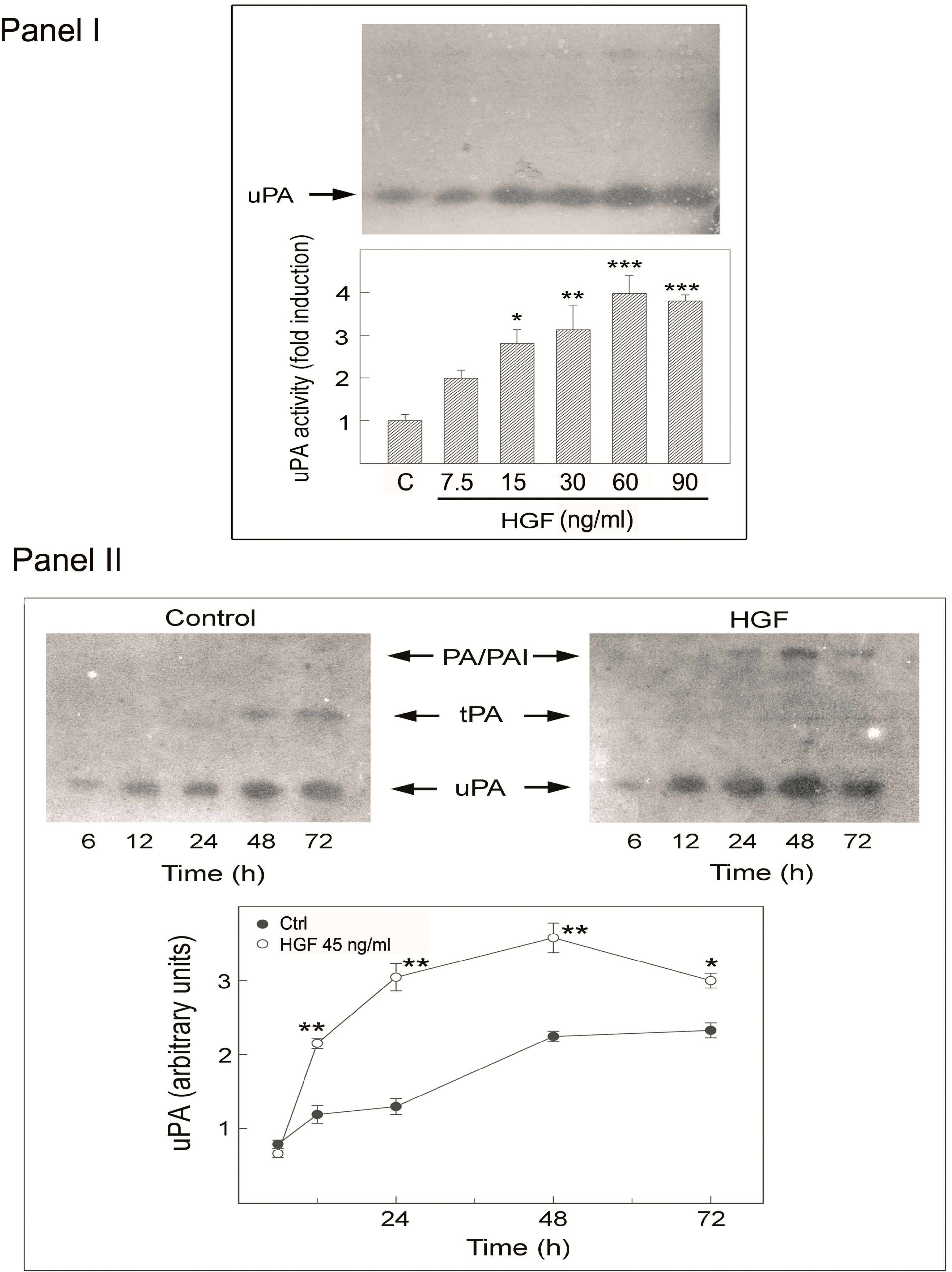
2.5. Effects of HGF on PA Activity in Cultured Rat Myoid Cells


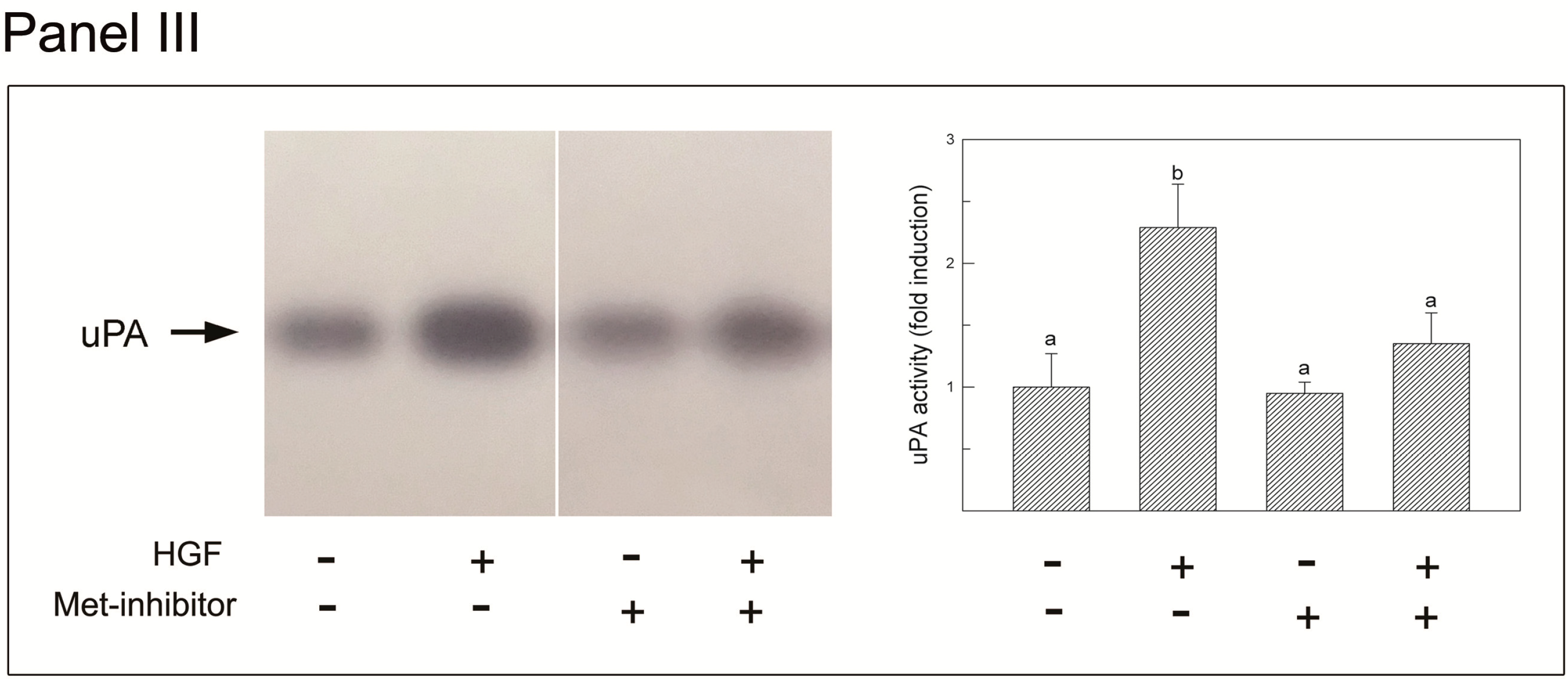
2.6. Effects of Met Inhibition on PA Activity in Cultured Adult Seminiferous Tubule Fragments
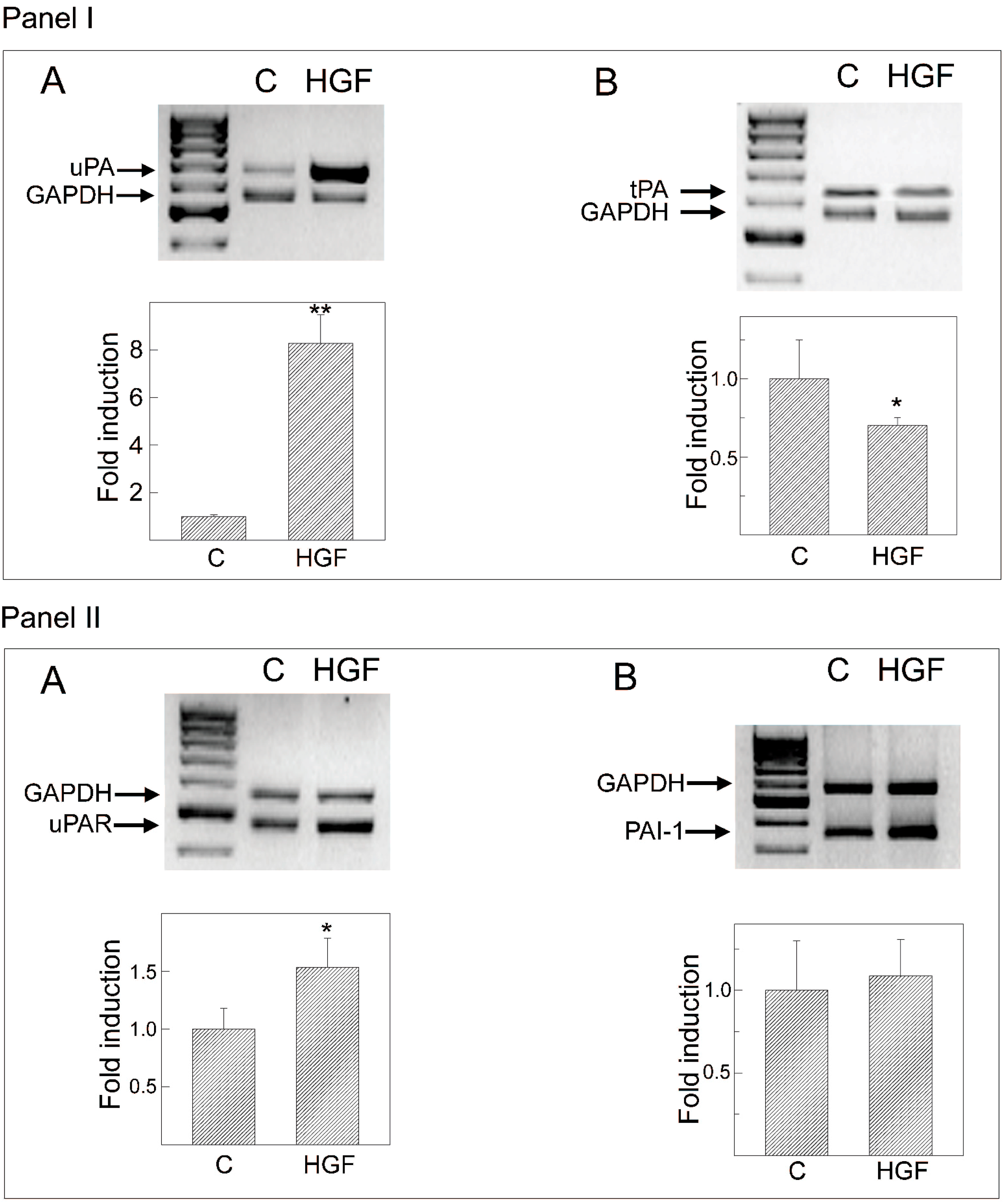
2.7. Hormonal Regulation of tPA, uPA, PAI-1 and uPAR mRNA Levels
2.8. Transforming Growth Factor-β (TGF-β) Production


3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Preparation and Culture
4.3. Isolation of Seminiferous Tubule Fragments
4.4. Met Immunofluorescence
4.5. F-Actin Detection
4.6. Confocal Microscopy
4.7. RNA Extraction and RT
4.8. Multiplex PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers | Product Length | Reaction Conditions | Gene Bank Accession Number |
|---|---|---|---|---|
| GAPDH | Fw 5'-TTCAACGGCACAGTCAAGGCT-3' | 552 bp | NM_017008.4 | |
| Rv 5'-ATTGGGGGTAGGAACACGGAA-3' | ||||
| uPA | Fw 5'-CCCTGCCTGGCCTGGAATTC-3' | 664 bp | Ann: 59 °C, 30" | NM_013085.3 |
| Rv 5'-CCAAACGGAGCATCACCAAACC-3' | Elong: 72 °C, 30" | |||
| uPAR | Fw 5'-GAAGACCAACAGGACCATGAGC-3' | 462 bp | Ann: 55 °C, 30" | NM_134352.1 |
| Rv 5'-CATCCAAAGGTGCTGTTCCC-3' | Elong: 72 °C, 30" | |||
| tPA | Fw 5'-GGGGGTACGTGTCAGCAGGCC-3' | 621 bp | Ann: 62 °C, 30" | NM_013151.2 |
| Rv 5'-CGCAGGTGGAGCATGGGGAC-3' | Elong: 72 °C, 30" | |||
| PAI-1 | Fw 5'-CTGGTGCTGGTGAACGCCCTC-3' | 338 bp | Ann: 59 °C, 30" | NM_012620.1 |
| Rv 5'-CAGGATGAGGAGGCGGGGC-3' | Elong: 72 °C, 30" |
4.9. Bioassay for TGF-β
4.10. Gel Electrophoresis and Zymography
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Nakamura, T.; Nishizawa, T.; Hagiya, M.; Seki, T.; Shimonishi, M.; Sugimura, A.; Tashiro, K.; Shimizu, S. Molecular cloning and expression of human hepatocyte growth factor. Nature 1989, 342, 440–443. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Nawa, K.; Ichihara, A. Partial purification and characterization of hepatocyte growth factor from serum of hepatectomized rats. Biochem. Biophys. Res. Commun. 1984, 122, 1450–1459. [Google Scholar] [CrossRef] [PubMed]
- Stoker, M.; Gherardi, E.; Perryman, M.; Gray, J. Scatter factor is a fibroblast-derived modulator of epithelial cell mobility. Nature 1987, 327, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Weidner, K.M.; Hartmann, G.; Sachs, M.; Birchmeier, W. Properties and functions of scatter factor/hepatocyte growth factor and its receptor c-Met. Am. J. Respir. Cell Mol. Biol. 1993, 8, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, F.F.; Pennington, C.J.; Kassiri, Z.; Rubin, J.S.; Soloway, P.D.; Ruther, U.; Edwards, D.R.; Khokha, R. Metalloproteinase inhibitor TIMP-1 affects hepatocyte cell cycle via HGF activation in murine liver regeneration. Hepatology 2005, 41, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Dean, M.; Kaul, K.; Braun, M.J.; Gonda, M.A.; Vande, W.G. Sequence of MET protooncogene cDNA has features characteristic of the tyrosine kinase family of growth-factor receptors. Proc. Natl. Acad. Sci. USA 1987, 84, 6379–6383. [Google Scholar] [CrossRef]
- Ricci, G.; Catizone, A. Pleiotropic activities of HGF/c-Met system in testicular physiology: Paracrine and endocrine implications. Front. Endocrinol. 2014, 5, 38. [Google Scholar] [CrossRef]
- Catizone, A.; Ricci, G.; Arista, V.; Innocenzi, A.; Galdieri, M. Hepatocyte growth factor and c-MET are expressed in rat prepuberal testis. Endocrinology 1999, 140, 3106–3113. [Google Scholar] [PubMed]
- Ricci, G.; Catizone, A.; Galdieri, M. Pleiotropic activity of hepatocyte growth factor during embryonic mouse testis development. Mech. Dev. 2002, 118, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Catizone, A.; Ricci, G.; Galdieri, M. Expression and functional role of hepatocyte growth factor receptor (C-MET) during postnatal rat testis development. Endocrinology 2001, 142, 1828–1834. [Google Scholar] [PubMed]
- Catizone, A.; Ricci, G.; Caruso, M.; Ferranti, F.; Canipari, R.; Galdieri, M. Hepatocyte growth factor (HGF) regulates blood-testis barrier (BTB) in adult rats. Mol. Cell Endocrinol. 2012, 348, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, I.; Kallajoki, M.; Narvanen, O.; Paranko, J.; Thornell, L.E.; Miettinen, M.; Lehto, V.P. Peritubular myoid cells of human and rat testis are smooth muscle cells that contain desmin-type intermediate filaments. Anat. Rec. 1986, 215, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Tung, P.S.; Fritz, I.B. Characterization of rat testicular peritubular myoid cells in culture: Alpha-smooth muscle isoactin is a specific differentiation marker. Biol. Reprod. 1990, 42, 351–365. [Google Scholar] [CrossRef] [PubMed]
- Worley, R.T.; Nicholson, H.D.; Pickering, B.T. Testicular oxytocin: An initiator of seminiferous tubule movement? 1985, 123, 205–212. [Google Scholar]
- Pickering, B.T.; Birkett, S.D.; Guldenaar, S.E.; Nicholson, H.D.; Worley, R.T.; Yavachev, L. Oxytocin in the testis: What, where, and why? Ann. N. Y. Acad. Sci. 1989, 564, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Buhrley, L.E.; Ellis, L.C. Contractility of rat testicular seminiferous tubules in vitro: Prostaglandin f 1 alpha and indomethacin-1,2. Prostaglandins 1975, 10, 151–162. [Google Scholar] [PubMed]
- Tripiciano, A.; Filippini, A.; Giustiniani, Q.; Palombi, F. Direct visualization of rat peritubular myoid cell contraction in response to endothelin. Biol. Reprod. 1996, 55, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Filippini, A.; Tripiciano, A.; Palombi, F.; Teti, A.; Paniccia, R.; Stefanini, M.; Ziparo, E. Rat testicular myoid cells respond to endothelin: Characterization of binding and signal transduction pathway. Endocrinology 1993, 133, 1789–1796. [Google Scholar] [PubMed]
- Catizone, A.; Ricci, G.; Galdieri, M. HGF and postnatal testis development. Mol. Cell Endocrinol. 2005, 241, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Catizone, A.; Ricci, G.; Galdieri, M. Hepatocyte growth factor modulates Sertoli–Sertoli tight junction dynamics. J. Cell Physiol. 2008, 216, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Catizone, A.; Ricci, G.; Tufano, M.A.; Perfetto, B.; Canipari, R.; Galdieri, M. Hepatocyte growth factor (HGF) modulates Leydig cell extracellular matrix components. J. Androl. 2010, 31, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, A.; Gangopadhyay, S.S.; Krishnan, R.; Ranpura, S.A.; Rajendran, K.; Ram-Mohan, S.; Mulone, M.; Gong, E.M.; Adam, R.M. JunB mediates basal- and TGFbeta1-induced smooth muscle cell contractility. PLoS One 2013, 8, e53430. [Google Scholar] [CrossRef] [PubMed]
- Nunes, I.; Shapiro, R.L.; Rifkin, D.B. Characterization of latent TGF-beta activation by murine peritoneal macrophages. J. Immunol. 1995, 155, 1450–1459. [Google Scholar] [PubMed]
- Dano, K.; Andreasen, A.; Grondahl-Hansen, J.; Kristensen, P.; Nielsen, L.S.; Skriver, L. Plasminogen activators, tissue degradation and cancer. Adv. Cancer Res. 1985, 44, 139–266. [Google Scholar] [PubMed]
- Vassalli, J.D.; Belin, D. Amiloride selectively inhibits the urokinase-type plasminogen activator. FEBS Lett. 1987, 214, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Anthony, C.T.; Rosselli, M.; Skinner, M.K. Actions of the testicular paracrine factor (P-Mod-S) on Sertoli cell transferrin secretion throughout pubertal development. Endocrinology 1991, 129, 353–360. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, S.; Blasi, F. The urokinase receptor as an entertainer of signal transduction. Front. Biosci. 2009, 14, 4575–4587. [Google Scholar] [CrossRef]
- Kjoller, L. The urokinase plasminogen activator receptor in the regulation of the actin cytoskeleton and cell motility. Biol. Chem. 2002, 383, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Lugano, R.; Pena, E.; Badimon, L.; Padro, T. Aggregated low-density lipoprotein induce impairment of the cytoskeleton dynamics through urokinase-type plasminogen activator/urokinase-type plasminogen activator receptor in human vascular smooth muscle cell. J. Thromb. Haemost. 2012, 10, 2158–2167. [Google Scholar] [CrossRef] [PubMed]
- Harris, G.C.; Nicholson, H.D. Stage-related differences in rat seminiferous tubule contractility in vitro and their response to oxytocin. J. Endocrinol. 1998, 157, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Vihko, K.K.; Suominen, J.J.; Parvinen, M. Cellular regulation of plasminogen activator secretion during spermatogenesis. Biol. Reprod. 1984, 31, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Parvinen, M. Regulation of the seminiferous epithelium. Endocr. Rev. 1982, 3, 404–417. [Google Scholar] [CrossRef] [PubMed]
- Vihko, K.K.; Penttila, T.L.; Parvinen, M.; Belin, D. Regulation of urokinase- and tissue-type plasminogen activator gene expression in the rat seminiferous epithelium. Mol. Endocrinol. 1989, 3, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Penttila, T.L.; Kaipia, A.; Toppari, J.; Parvinen, M.; Mali, P. Localization of urokinase- and tissue-type plasminogen activator mRNAs in rat testes. Mol. Cell Endocrinol. 1994, 105, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Russel, L.D. Movement of spermatocytes from the basal to the adluminal compartment of the rat testis. Am. J. Anat. 1977, 148, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Leblond, C.P.; Clermont, Y. Definition of the stages of the cycle of the seminiferous epithelium of the rat. Ann. N. Y. Acad. Sci. 1952, 55, 548–573. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.; Clermont, Y.; Nadler, N.J. Cyclic endocytic activity and kinetics of lysosomes in Sertoli cells of the rat: A morphometric analysis. Biol. Reprod. 1986, 34, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Odekon, L.E.; Blasi, F.; Rifkin, D.B. Requirement for receptor-bound urokinase in plasmin-dependent cellular conversion of latent TGF-beta to TGF-beta. J. Cell Physiol. 1994, 158, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Ailenberg, M.; Tung, P.S.; Fritz, I.B. Transforming growth factor-beta elicits shape changes and increases contractility of testicular peritubular cells. Biol. Reprod. 1990, 42, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Palombi, F.; di Carlo, C. Alkaline phosphatase is a marker for myoid cells in cultures of rat peritubular and tubular tissue. Biol. Reprod. 1988, 39, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Parvinen, M.; Vanha-Perttula, T. Identification and enzyme quantitation of the stages of the seminiferous epithelial wave in the rat. Anat. Rec. 1972, 174, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Gokhale, V.M.; Szabo, L.; Munoz, R.M.; Baek, H.; Bashyam, S.; Hurley, L.H.; von Hoff, D.D.; Han, H. Identification of a novel inhibitor of urokinase-type plasminogen activator. Mol. Cancer Ther. 2007, 6, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Harpel, J.G.; Metz, C.N.; Nunes, I.; Loskutoff, D.J.; Rifkin, D.B. An assay for transforming growth factor-beta using cells transfected with a plasminogen activator inhibitor-1 promoter-luciferase construct. Anal. Biochem. 1994, 216, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Belin, D.; Godeau, F.; Vassalli, J.D. Tumor promoter PMA stimulates the synthesis and secretion of mouse pro-urokinase in MSV-transformed 3T3 cells: This is mediated by an increase in urokinase mRNA content. EMBO J. 1984, 3, 1901–1906. [Google Scholar] [PubMed]
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catizone, A.; Ricci, G.; Caruso, M.; Galdieri, M.; Scheri, K.C.; Di Paolo, V.; Canipari, R. HGF Modulates Actin Cytoskeleton Remodeling and Contraction in Testicular Myoid Cells. Biomedicines 2015, 3, 89-109. https://doi.org/10.3390/biomedicines3010089
Catizone A, Ricci G, Caruso M, Galdieri M, Scheri KC, Di Paolo V, Canipari R. HGF Modulates Actin Cytoskeleton Remodeling and Contraction in Testicular Myoid Cells. Biomedicines. 2015; 3(1):89-109. https://doi.org/10.3390/biomedicines3010089
Chicago/Turabian StyleCatizone, Angela, Giulia Ricci, Maria Caruso, Michela Galdieri, Katia Corano Scheri, Virginia Di Paolo, and Rita Canipari. 2015. "HGF Modulates Actin Cytoskeleton Remodeling and Contraction in Testicular Myoid Cells" Biomedicines 3, no. 1: 89-109. https://doi.org/10.3390/biomedicines3010089
APA StyleCatizone, A., Ricci, G., Caruso, M., Galdieri, M., Scheri, K. C., Di Paolo, V., & Canipari, R. (2015). HGF Modulates Actin Cytoskeleton Remodeling and Contraction in Testicular Myoid Cells. Biomedicines, 3(1), 89-109. https://doi.org/10.3390/biomedicines3010089