The Chemistry, Pharmacology and Therapeutic Potential of the Edible Mushroom Dictyophora indusiata (Vent ex. Pers.) Fischer (Synn. Phallus indusiatus)

Abstract
:
1. Introduction
2. Overview of Chemistry
2.1. Polysaccharides
2.2. Terpenoids
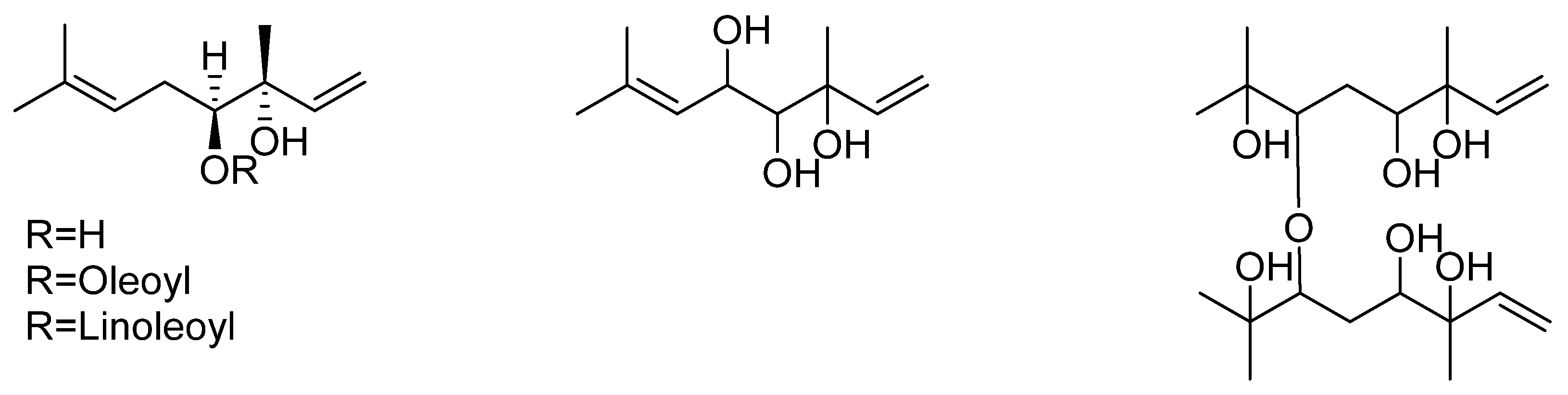
2.3. Alkaloids
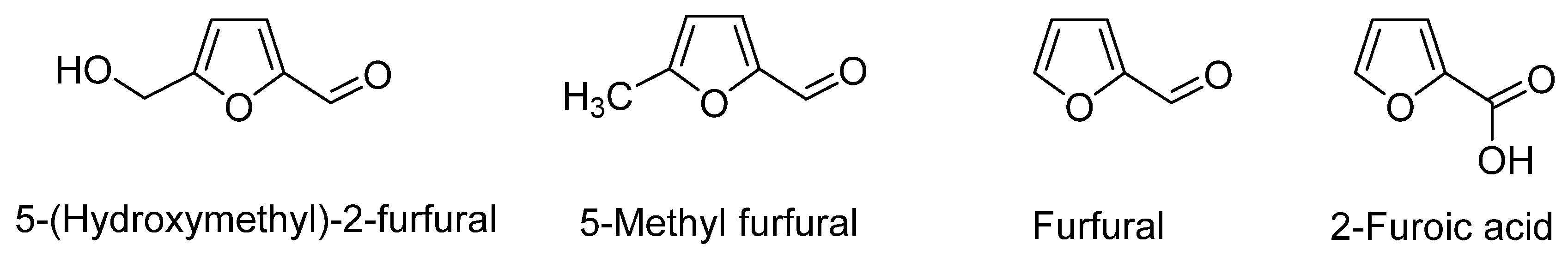
2.4. 5-(Hydroxymethyl)-2-Furfural
3. The Pharmacology of D. indusiata Polysaccharides
3.1. Antioxidant Effect
3.2. Neuroprotective Effect and Potential Application in Neurodegenerative Diseases
3.3. Anticancer Effect
3.4. Immunomodulatory Effect
3.4.1. Immunostimulation
3.4.2. General Anti-Inflammatory, Immunosuppressive and Effect on the Gut Microbiota
3.5. Antiobesity and Potential Antidiabetic Effect
3.6. Antibacterial Effects
3.7. Other Effects
3.8. Toxicity Remarks
3.9. General Summary and Conclusion
Funding
Conflicts of Interest
References
- Ker, Y.B.; Chen, K.C.; Peng, C.C.; Hsieh, C.L.; Peng, R.Y. Structural characteristics and antioxidative capability of the soluble polysaccharides present in Dictyophora indusiata (Vent. Ex Pers.) Fish Phallaceae. eCAM 2011, 2011, 396013. [Google Scholar] [PubMed] [Green Version]
- Sitinjak, R.R. The Nutritional Content of the Mushroom Phallus indusiatus Vent, which Grows in the Cocoa Plantation, Gaperta-Ujung, Medan. Pharma. Chem. 2017, 9, 44–47. [Google Scholar]
- Ouyang, S.; Luo, Y.; Liu, M.; Fan, J.; Guo, X.; Deng, F. Analysis of amino acids, vitamins and inorganic elements in Dictyophora indusiata. Hunan Yi Ke Da Xue Xue Bao 1998, 23, 535–536. [Google Scholar] [PubMed]
- Wang, Y.; Ji, X.; Yan, M.; Chen, X.; Kang, M.; Teng, L.; Wu, X.; Chen, J.; Deng, C. Protective effect and mechanism of polysaccharide from Dictyophora indusiata on dextran sodium sulfate-induced colitis in C57BL/6 mice. Int. J. Biol. Macromol. 2019, 140, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Hara, C.; Kiho, T.; Ukai, S. Polysaccharides in fungi.12. The location of the o-acetyl groups in the (1–3)-alpha-d-mannan from dictyophora-indusiata fisch. Carbohydr. Res. 1982, 111, 143–150. [Google Scholar] [CrossRef]
- Hua, Y.L.; Gao, Q.; Wen, L.R.; Yang, B.; Tang, J.; You, L.J.; Zhao, M.M. Structural characterisation of acid- and alkali-soluble polysaccharides in the fruiting body of Dictyophora indusiata and their immunomodulatory activities. Food Chem. 2012, 132, 739–743. [Google Scholar] [CrossRef]
- Kanwal, S.; Joseph, T.P.; Owusu, L.; Ren, X.M.; Li, M.Q.; Xin, Y. A Polysaccharide Isolated from Dictyophora indusiata promotes recovery from antibiotic-driven intestinal dysbiosis and improves gut epithelial barrier function in a mouse model. Nutrients 2018, 10, 1003. [Google Scholar] [CrossRef] [Green Version]
- Hara, C.; Kiho, T.; Ukai, S. A branched (1→3)-β-d-glucan from a sodium carbonate extract of Dictyophora indusiata fisch. Carbohydr. Res. 1983, 117, 201–213. [Google Scholar] [CrossRef]
- Hara, C.; Kiho, T.; Ukai, S. A branched (1→3)-β-d-glucan from a water extract of Dictyophora indusiata Fisch. Carbohydr. Res. 1986, 145, 237–246. [Google Scholar] [CrossRef]
- Hara, C.; Yokomori, Y.; Kiho, T.; Nagai, K.; Ukai, S. A (1→3)-α-D-mannan from a water extract of Dictyophora indusiata Fisch. Carbohydr. Res. 1988, 173, 332–338. [Google Scholar] [CrossRef]
- Ukai, S.; Hara, C.; Kiho, T.; Hirose, K. Polysaccharised in fungi.5. Isolation and characterisation of a mannan from aqueous ethanol extract of Dictyophora indusiata Fisch. Chem. Pharm. Bull. 1980, 28, 2647–2652. [Google Scholar] [CrossRef] [Green Version]
- Ukai, S.; Hara, C.; Kiho, T. Polysaccharised in fungi.9. A β-D-glucan from alkaline extract of Dictyophora indusiata Fisch. Chem. Pharm. Bull. 1982, 30, 2147–2154. [Google Scholar] [CrossRef] [Green Version]
- Deng, C.; Fu, H.; Xu, J.; Shang, J.; Cheng, Y. Physiochemical and biological properties of phosphorylated polysaccharides from Dictyophora indusiata. Int. J. Biol. Macromol. 2015, 72, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.S.; Song, X.L.; Zhang, J.J.; Li, H.P.; Liu, M.; Gao, Z.; Wang, X.X.; Jia, L. Antioxidation, hepatic- and renal-protection of water-extractable polysaccharides by Dictyophora indusiata on obese mice. Int. J. Biol. Macromol. 2019, 134, 290–301. [Google Scholar] [CrossRef]
- Deng, C.; Fu, H.T.; Teng, L.P.; Hu, Z.; Xu, X.F.; Chen, J.H.; Ren, T.L. Anti-tumor activity of the regenerated triple-helical polysaccharide from Dictyophora indusiata. Int. J. Biol. Macromol. 2013, 61, 453–458. [Google Scholar] [CrossRef]
- Fu, Y.; Lin, S.; Lu, M.; Wei, S.Y.; Zhou, J.; Zhao, L.; Zhang, Q.; Lin, D.R.; Liu, Y.T.; Chen, H.; et al. Quantitative evaluation of ultrasound-assisted extraction of 1,3-beta-glucans from Dictyophora indusiata using an improved fluorometric assay. Polymers 2019, 11, 864. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.H.; Gong, G.L.; Wang, Y.Y.; Li, F.; Jia, S.Y.; Qin, F.X.; Ren, H.T.; Liu, Y. Response surface optimization of enzyme-assisted extraction polysaccharides from Dictyophora indusiata. Int. J. Biol. Macromol. 2013, 61, 63–68. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Y.; Wu, L.; Wu, X.; Huang, Y.; Liu, B. Optimization of polysaccharides extraction from Dictyophora indusiata and determination of its antioxidant activity. Int. J. Biol. Macromol. 2017, 103, 175–181. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, R.N.; Li, H.F.; Xiang, Y.X.; Xiao, L.Y.; Hu, M.H.; Ma, F.L.; Ma, C.W.; Huang, Z. Antioxidant and neuroprotective effects of Dictyophora indusiata polysaccharide in Caenorhabditis elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef]
- Deng, C.; Fu, H.T.; Shang, J.Y.; Chen, J.H.; Xu, X. Dectin-1 mediates the immunoenhancement effect of the polysaccharide from Dictyophora indusiata. Int. J. Biol. Macromol. 2018, 109, 369–374. [Google Scholar] [CrossRef]
- Deng, C.; Hu, Z.; Fu, H.T.; Hu, M.H.; Xu, X.; Chen, J.H. Chemical analysis and antioxidant activity in vitro of a beta-D-glucan isolated from Dictyophora indusiata. Int. J. Biol. Macromol. 2012, 51, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Hua, Y.L.; Yang, B.; Tang, J.; Ma, Z.H.; Gao, Q.; Zhao, M.M. Structural analysis of water-soluble polysaccharides in the fruiting body of Dictyophora indusiata and their in vivo antioxidant activities. Carbohydr. Polym. 2012, 87, 343–347. [Google Scholar] [CrossRef]
- Ishiyama, D.; Fukushi, Y.; Ohnishi-Kameyama, M.; Nagata, T.; Mori, H.; Inakuma, T.; Ishiguro, Y.; Li, J.X.; Kawagishi, H. Monoterpene-alcohols from a mushroom Dictyophora indusiata. Phytochemistry 1999, 50, 1053–1056. [Google Scholar] [CrossRef]
- Kawagishi, H.; Ishiyama, D.; Mori, H.; Sakamoto, H.; Ishiguro, Y.; Furukawa, S.; Li, J.X. Dictyophorines A and B, two stimulators of NGF-synthesis from the mushroom Dictyophora indusiata. Phytochemistry 1997, 45, 1203–1205. [Google Scholar] [CrossRef]
- Huang, M.Q.; Chen, X.; Tian, H.Y.; Sun, B.G.; Chen, H.T. Isolation and identification of antibiotic albaflavenone from Dictyophora indusiata (Vent:Pers.) Fischer. J. Chem. Res. 2011, 11, 659–660. [Google Scholar] [CrossRef]
- Lee, I.K.; Yun, B.S.; Han, G.; Cho, D.H.; Kim, Y.H.; Yoo, I.D. Dictyoquinazols A, B, and C, new neuroprotective compounds from the mushroom Dictyophora indusiata. J. Nat. Prod. 2002, 65, 1769–1772. [Google Scholar] [CrossRef]
- Lizarme, Y.; Wangsahardja, J.; Marcolin, G.M.; Morris, J.C.; Jones, N.M.; Hunter, L. Synthesis and neuroprotective activity of dictyoquinazol A and analogues. Bioorg. Med. Chem. 2016, 24, 1480–1487. [Google Scholar] [CrossRef]
- Oh, C.H.; Chung Song, H. Total synthesis of neuroprotective dictyoquinazol A, B, and C. Synth. Commun. 2007, 37, 3311–3317. [Google Scholar] [CrossRef]
- Sharma, V.K.; Choi, J.; Sharma, N.; Choi, M.; Seo, S.Y. In vitro anti-tyrosinase activity of 5-(Hydroxymethyl)-2-furfural isolated from Dictyophora indusiata. Phytother. Res. 2004, 18, 841–844. [Google Scholar] [CrossRef]
- García-Villanova, B.; Guerra-Hernández, E.; Martínez-Gómez, E.; Montilla, J. Liquid chromatography for the determination of 5-(hydroxymethyl)-2-furaldehyde in breakfast cereals. J. Agric. Food Chem. 1993, 41, 1254–1255. [Google Scholar] [CrossRef]
- Meidell, E.; Filipello, F. Quantitative determination of hydroxymethylfurfural in sherries and grape concentrate. Am. J. Enol. Viticult. 1969, 20, 164–168. [Google Scholar]
- Poll, L. The influence of apple ripeness and juice storage temperature on the sensory evaluation and composition (volatile and non-volatile components) of apple juice. Lebensm. Wiss. Technol. 1985, 18, 205–211. [Google Scholar]
- Porretta, S.; Sandei, L. Determination of 5-(hydroxymethyl)-2-furfural (HMF) in tomato products: Proposal of a rapid HPLC method and its comparison with the colorimetric method. Food Chem. 1991, 39, 51–57. [Google Scholar] [CrossRef]
- Antal, M.J.; Mok, W.S.L.; Richards, G.N. Mechanism of formation of 5-(hydroxymethyl)-2-furaldehyde from d-fructose and sucrose. Carbohydr. Res. 1990, 199, 91–109. [Google Scholar] [CrossRef]
- Wang, W.; Song, X.; Gao, Z.; Zhao, H.; Wang, X.; Liu, M.; Jia, L. Anti-hyperlipidemic, antioxidant and organic protection effects of acidic-extractable polysaccharides from Dictyophora indusiata. Int. J. Biol. Macromol. 2019, 129, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lai, L.; Teng, L.; Li, Y.; Cheng, J.; Chen, J.; Deng, C. Mechanism of the anti-inflammatory activity by a polysaccharide from Dictyophora indusiata in lipopolysaccharide-stimulated macrophages. Int. J. Biol. Macromol. 2019, 126, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Han, S.Y.; Ma, C.X.; Hu, M.H.; Wang, Y.Y.; Ma, F.L.; Tao, N.; Qin, Z.H. A polysaccharide from Dictyophora indusiata inhibits the immunosuppressive function of cancer-associated fibroblasts. Cell Biochem. Funct. 2017, 35, 414–419. [Google Scholar] [CrossRef]
- Deng, C.; Shang, J.Y.; Fu, H.T.; Chen, J.X.; Liu, H.Y.; Chen, J.H. Mechanism of the immunostimulatory activity by a polysaccharide from Dictyophora indusiata. Int. J. Biol. Macromol. 2016, 91, 752–759. [Google Scholar] [CrossRef]
- Liao, W.; Chen, L.; Yu, B.; Lei, Z.; Wu, X.; Yang, J.; Ren, J. Cell-based evaluation of a novel Dictyophora indusiata polysaccharide against oxidative-induced erythrocyte hemolysis. Cell. Mol. Biol. 2016, 62, 38–44. [Google Scholar]
- Liao, W.Z.; Yu, Z.Q.; Lin, Z.H.; Lei, Z.G.; Ning, Z.X.; Regenstein, J.M.; Yang, J.G.; Ren, J.Y. Biofunctionalization of Selenium Nanoparticle with Dictyophora indusiata Polysaccharide and Its Antiproliferative Activity through Death-Receptor and Mitochondria-Mediated Apoptotic Pathways. Sci. Rep. 2015, 5, 18629. [Google Scholar] [CrossRef]
- Liao, W.Z.; Lu, Y.J.; Fu, J.N.; Ning, Z.X.; Yang, J.G.; Ren, J.Y. Preparation and Characterization of Dictyophora indusiata Polysaccharide-Zinc Complex and Its Augmented Antiproliferative Activity on Human Cancer Cells. J. Agric. Food Chem. 2015, 63, 6525–6534. [Google Scholar] [CrossRef] [PubMed]
- Liao, W.Z.; Luo, Z.; Liu, D.; Ning, Z.X.; Yang, J.G.; Ren, J.Y. Structure Characterization of a Novel Polysaccharide from Dictyophora indusiata and Its Macrophage Immunomodulatory Activities. J. Agric. Food Chem. 2015, 63, 535–544. [Google Scholar] [CrossRef]
- Deng, C.; Xu, J.J.; Fu, H.T.; Chen, J.H.; Xu, X. Characterization, antioxidant and cytotoxic activity of sulfated derivatives of a water-insoluble polysaccharides from Dictyophora indusiata. Mol. Med. Rep. 2015, 11, 2991–2998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, H.; Deng, C.; Teng, L.; Yu, L.; Su, T.; Xu, X.; Chen, J.; Yang, C. Immunomodulatory Activities on RAW 264.7 Macrophages of a Polysaccharide from Veiled Lady Mushroom, Dictyophora indusiata (Higher Basidiomycetes). Int. J. Med. Mushrooms 2015, 17, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Zhong, B.; Ma, Y.S.; Fu, D.; Zhang, C. Induction of apoptosis in osteosarcoma s180 cells by polysaccharide from Dictyophora indusiata. Cell Biochem. Funct. 2013, 31, 719–723. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Wang, Z.Y.; Wang, L.; Walid, E.; Zhang, H. In Vitro Antioxidant and Anti-Proliferation Activities of Polysaccharides from Various Extracts of Different Mushrooms. Int. J. Mol. Sci. 2012, 13, 5801–5817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, C.; Kumazawa, Y.; Inagaki, K.; Kaneko, M.; Kiho, T.; Ukai, S. Mitogenic and colony-stimulating factor-inducing activities of polysaccharide fractions from the fruit bodies of Dictyophora indusiata Fisch. Chem. Pharm. Bull. 1991, 39, 1615–1616. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, H.; Zhang, Y.; Feng, Y.; Yuan, F.; Song, X.; Gao, Z.; Zhang, J.; Song, Z. Antihyperlipidemic and hepatoprotective properties of alkali- and enzyme-extractable polysaccharides by Dictyophora indusiata. Sci. Rep. 2019, 9, 14266. [Google Scholar] [CrossRef] [Green Version]
- Kanwal, S.; Joseph, T.P.; Aliya, S.; Song, S.; Saleem, M.Z.; Nisar, M.A.; Wang, Y.; Meyiah, A.; Ma, Y.; Xin, Y. Attenuation of DSS induced colitis by Dictyophora indusiata polysaccharide (DIP) via modulation of gut microbiota and inflammatory related signaling pathways. J. Func. Foods 2019, in press. [Google Scholar] [CrossRef]
- Ukai, S.; Kiho, T.; Hara, C.; Kuruma, I.; Tanaka, Y. Polysaccharides in fungi. XIV. Anti-inflammatory effect of the polysaccharides from the fruit bodies of several fungi. J. Pharmacobiodyn. 1983, 6, 983–990. [Google Scholar] [CrossRef] [Green Version]
- Ukai, S.; Kiho, T.; Hara, C.; Morita, M.; Goto, A.; Imaizumi, N.; Hasegawa, Y. Polysaccharides in fungi. XIII. Antitumor activity of various polysaccharides isolated from Dictyophora indusiata, Ganoderma japonicum, Cordyceps cicadae, Auricularia auricula-judae, and Auricularia species. Chem. Pharm. Bull. 1983, 31, 741–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, C.; Kiho, T.; Tanaka, Y.; Ukai, S. Anti-inflammatory activity and conformational behavior of a branched (1 leads to 3)-beta-D-glucan from an alkaline extract of Dictyophora indusiata Fisch. Carbohydr. Res. 1982, 110, 77–87. [Google Scholar] [CrossRef]
- Oyetayo, V.O.; Dong, C.H.; Yao, Y.J. Antioxidant and antimicrobial properties of aqueous extract from Dictyophora indusiata. Open Mycol. J. 2009, 3, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; D’Amora, D.R.; MacNeil, L.T.; Walhout, A.J.M.; Kubiseski, T.J. The Ooxidative stress response in Caenorhabditis elegans requires the GATA transcription factor ELT-3 and SKN-1/Nrf2. Genetics 2017, 206, 1909–1922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Chen, W.-D.; Wang, Y.-D. DAF-16/FOXO Transcription factor in aging and longevity. Front. Pharmacol. 2017, 8, 548. [Google Scholar] [CrossRef]
- Tullet, J.M.A.; Green, J.W.; Au, C.; Benedetto, A.; Thompson, M.A.; Clark, E.; Gilliat, A.F.; Young, A.; Schmeisser, K.; Gems, D. The SKN-1/Nrf2 transcription factor can protect against oxidative stress and increase lifespan in C. elegans by distinct mechanisms. Aging Cell. 2017, 16, 1191–1194. [Google Scholar] [CrossRef] [Green Version]
- Habtemariam, S. The brain-derived neurotrophic factor in neuronal plasticity and neuroregeneration: New pharmacological concepts for old and new drugs. Neural Regen. Res. 2018, 13(6), 983–984. [Google Scholar] [CrossRef]
- Habtemariam, S.; Lentini, G. Plant-derived anticancer agents: Lessons from the pharmacology of geniposide and its aglycone, genipin. Biomedicines 2018, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- Neamatallah, T. Mitogen-activated protein kinase pathway: A critical regulator in tumor-associated macrophage polarization. J. Microsc. Ultrastruct. 2019, 7, 53–56. [Google Scholar] [CrossRef]
- Fang, Q.; Wang, J.F.; Zha, X.Q.; Cui, S.H.; Cao, L.; Luo, J.P. Immunomodulatory activity on macrophage of a purified polysaccharide extracted from Laminaria japonica. Carbohydr Polym. 2015, 134, 66–73. [Google Scholar] [CrossRef]
- Gürtler, H.; Pedersen, R.; Anthoni, U.; Christophersen, C.; Nielsen, P.H.; Wellington, E.M.; Pedersen, C.; Bock, K. Albaflavenone, a sesquiterpene ketone with a zizaene skeleton produced by a streptomycete with a new rope morphology. J. Antibiot. 1994, 47, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Cane, D.E. Biosynthesis of the sesquiterpene antibiotic albaflavenone in Streptomyces coelicolor. Mechanism and stereochemistry of the enzymatic formation of epi-isozizaene. J. Am. Chem. Soc. 2009, 131, 6332–6333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moody, S.C.; Zhao, B.; Lei, L.; Nelson, D.R.; Mullins, J.G.; Waterman, M.R.; Kelly, S.L.; Lamb, D.C. Investigating conservation of the albaflavenone biosynthetic pathway and CYP170 bifunctionality in streptomycetes. FEBS J. 2012, 279, 1640–1649. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lin, X.; Lei, L.; Lamb, D.C.; Kelly, S.L.; Waterman, M.R.; Cane, D.E. Biosynthesis of the sesquiterpene antibiotic albaflavenone in Streptomyces coelicolor A3. J. Biol. Chem. 2008, 283, 8183–8189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.X.; Ng, T.B. A novel ribonuclease from the veiled lady mushroom Dictyophora indusiata. Biochem. Cell Biol. 2003, 81, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Zhao, L.; Wang, H.; Ng, T.B. A novel ribonuclease with antiproliferative activity toward leukemia and lymphoma cells and HIV-1 reverse transcriptase inhibitory activity from the mushroom, Hohenbuehelia serotina. Int. J. Mol. Med. 2013, 33, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, R.; Dalvi, Y.B.; Lamrood, P.Y.; Shinde, B.P.; Nair, C.K.K. Historical and current perspectives on therapeutic potential of higher basidiomycetes: An overview. Biotechnology 2019, 9, 362. [Google Scholar] [CrossRef]
- Friedman, M. Mushroom polysaccharides: Chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [Green Version]
- Su, C.H.; Lai, M.N.; Lin, C.C.; Ng, L.T. Comparative characterization of physicochemical properties and bioactivities of polysaccharides from selected medicinal mushrooms. Appl. Microbiol. Biotechnol. 2016, 100, 4385–4393. [Google Scholar] [CrossRef]
- Wu, Y.S.; Ho, S.Y.; Nan, F.H. Ganoderma lucidum β 1,3/1,6 glucan as an immunomodulator in inflammation induced by a high-cholesterol diet. BMC Complement. Altern. Med. 2016, 16, 500. [Google Scholar] [CrossRef] [Green Version]
- Habtemariam, 2019. The Nrf2/HO-1 axis as targets for flavanones: Neuroprotection by pinocembrin, naringenin and eriodictyol. Oxid. Med. Cell. Long. 2019, 2019, 4724920. [Google Scholar]
- Habtemariam, S. Iridoids and other monoterpenes in the Alzheimer’s brain: Recent development and future prospects. Molecules 2018, 23, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habtemariam, S. Antioxidant and anti-inflammatory mechanisms of neuroprotection by ursolic acid: Addressing brain injury, cerebral ischemia, cognition deficit, anxiety, and depression. Oxid. Med. Cell Longev. 2019, 16, 8512048. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Zhang, M.; Jiang, Y.; Liu, Y.; Luo, H.; Hao, C.; Zeng, P.; Zhang, L. Preclinical and clinical studies of Coriolus versicolor polysaccharopeptide as an immunotherapeutic in China. Discov. Med. 2017, 23, 207–219. [Google Scholar] [PubMed]
- Dou, H.; Chang, Y.; Zhang, L. Coriolus versicolor polysaccharopeptide as an immunotherapeutic in China. Prog. Mol. Biol. Transl. Sci. 2019, 163, 361–381. [Google Scholar]
- Yang, S.F.; Zhuang, T.F.; Si, Y.M.; Qi, K.Y.; Zhao, J. Coriolus versicolor mushroom polysaccharides exert immunoregulatory effects on mouse B cells via membrane Ig and TLR-4 to activate the MAPK and NF-κB signaling pathways. Mol. Immunol. 2015, 64, 144–151. [Google Scholar] [CrossRef]
- Giannenasa, I.; Tsalie, E.B.; Chronisc, E.F. Consumption of Agaricus bisporus mushroom affects the performance, intestinal microbiota composition and morphology, and antioxidant status of turkey poults. Anim. Feed Sci. Technol. 2011, 165, 218–229. [Google Scholar] [CrossRef]
- Singdevsachan, S.K.; Mishra, P.A.J.; Baliyarsingh, B.; Tayung, K.; Thatoi, H. Mushroom polysaccharides as potential prebiotics with their antitumor and immunomodulating properties: A review. Bioact. Carbohydr. Diet. Fiber 2015, 7, 1–14. [Google Scholar] [CrossRef]
- Xiao, C.; Wu, Q.P.; Cai, W.; Tan, J.B.; Yang, X.B.; Zhang, J.M. Hypoglycemic effects of Ganoderma lucidum polysaccharides in type 2 diabetic mice. Arch Pharm. Res. 2012, 35, 1793–1801. [Google Scholar] [CrossRef]
- Ma, H.T.; Hsieh, J.F.; Chen, S.T. Anti-diabetic effects of Ganoderma lucidum. Phytochemistry 2015, 114, 109–113. [Google Scholar] [CrossRef]
- Pan, D.; Zhang, D.; Wu, J.; Chen, C.; Xu, Z.; Yang, H.; Zhou, P. Antidiabetic, antihyperlipidemic and antioxidant activities of a novel proteoglycan from Ganoderma lucidum fruiting bodies on db/db mice and the possible mechanism. PLoS ONE 2013, 8, e68332. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.; Young, J.D.; et al. Ganoderma lucidum reduces obesity in mice by modulating the composition of the gut microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Xiao, D.; Liu, W.; Song, Y.; Zou, B.; Li, L.; Li, P.; Cai, Y.; Liu, D.; Liao, Q.; et al. Intake of Ganoderma lucidum polysaccharides reverses the disturbed gut microbiota and metabolism in type 2 diabetic rats. Int. J. Biol. Macromol 2019. pii: S0141-8130(19)34451-4. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparation | Experimental Model | Key Findings | References |
|---|---|---|---|
| Water-extractable polysaccharides | Antioxidant assay—0.2–1.0 mg/mL | Scavenge DPPH and OH. radicals; possess reducing power. | Wang et al. [14] |
| Acid-extractable polysaccharides -average molecular weight of estimated to be 7.89 × 105 Da, 4.64 × 105 Da and 6.41 × 105 Da | Antioxidant assay—0.2–1.4 mg/mL | OH, O2− and DPPH radical scavenging effect. | Wang et al. [35] |
| Polysaccharides | LPS-stimulated RAW264.7 macrophages—25 μg/mL or 50 μg/mL | Inhibit the expression of TLR4, phosphorylated-IκBα and NF-κB nuclear translocation (p65 subunit); decrease the extracellular protein level of pro-caspase-1 and cytoplasmic pool of NLRP3; inhibit the level of activated caspase-1, and caspase-1-mediated IL-1 and IL-18 production; inhibit IL-1 and ROS production. | Wang et al. [36] |
| Purified polysaccharide—Dectin-1 | RAW264.7 cells—200 μg/mL | Induce pseudopodia formation and cell spreading; increase phagocytic uptake; enhance IL-1β and TNF-α; induce phosphorylation of ERK1/2, JNK and p38; enhance the phosphorylation of NF-κB p65 and NF-κB nuclear translocation; specifically bind to macrophages through TLR4. | Deng et al. [20] |
| Polysaccharides | Radical scavenging assay | EC50 for DPPH, OH. and O2− was 0.89 mg/mL, 0.51 mg/mL and 0.68 mg/mL respectively. | Liu et al. [18] |
| Polysaccharides | Immunosuppressive effect of prostate-cancer-associated fibroblasts—supernatant of prostate cancer fibroblasts on lymphocyte growth—0.1–0.8 mg/mL | Stimulate the proliferation of immune cells and reverse inhibition of the growth of CD4+/CD8+ T cells; down-regulate the expression of α-smooth muscle actin. | Han et al. [37] |
| Polysaccharides | RAW264.7 cells—25–200 μg/mL | Proliferative effect, up-regulate the production of NO, IL-1β, IL-6 and TNF-α (both protein and m RNA expression); effect mediated by TLR4 (inhibited by anti-TLR4 and anti-CR3 mAb); specifically bind to macrophage in TLR4-dependent manner; activate NF-κB p65. | Deng et al. [38] |
| Purified polysaccharide (DP1) | 2, 2-azobis (2-amidino-propane) dihydrochloride -induced erythrocyte hemolysis assay—20 nmol/mL | Inhibit hemolytic activity by 87.4%; inhibit ROS overproduction by 81.5%; suppress MDA level by 57.0%; inhibit the cupric chloride- induced conjugated diene formation in plasma; enhance intracellular antioxidant enzymes (SOD, GPX and CAT) activities. | Liao et al. [39] |
| Purified polysaccharide (DP1)—monodispersed selenium nanoparticles | HepG2 cells—125, 250 or 500 μg/mL | Induce cell apoptosis—nuclear condensation, DNA cleavage and accumulation of S-phase cell arrest; activate caspases 3, 8 and 9; induce ROS overproduction and mitochondrial dysfunction. | Liao et al. [40] |
| Purified polysaccharide (DP1) chelated with zinc chloride | MCF-7 cells—125 or 250 μg/mL | Induce apoptosis—DNA breakage, and S-phase cell cycle arrest, activate caspases-3, -8, and -9, mitochondrial dysfunction, and ROS overproduction. | Liao et al. [41] |
| Polysaccharide—an average molecular weight of 1132 kDa and consisted of glucose (56.2%), galactose (14.1%), and mannose (29.7%). The main linkage type of DP1 were proven to be (1→3)-linked α-L-Man, (1→2, 6)-linked α-D-Glc, (1→6)-linked β-D-Glc, (1→6)-linked β-D-Gal, and (1→6)-linked β-D-Man | RAW 264.7 cells—up to 250 μg/mL | Promote macrophage NO, TNF-α, and IL-6 secretion. | Liao et al. [42] |
| Phosphorylated polysaccharides prepared from a water-insoluble polysaccharides | Radical scavenging assay; cytotoxic effects against MCF-7 and B16 cells | Comparative study—phosphorylated derivative more potent than native polysaccharides. | Deng et al. [13] |
| Water-soluble sulfated polysaccharides—prepared from water insoluble polysaccharides | OH. and DPPH radical scavenging assay; cytotoxicity against MCF-7 and B16 cells | Comparative study—Sulfated derivative more potent than native polysaccharides. | Deng et al. [43] |
| Water-soluble polysaccharide of β-(1→3)-glucan with side branches of β-(1→6)-glucosyl unit—triple-helical structure | RAW 264.7 macrophage—50–200 µg/mL | Promote macrophage cell proliferation; increase NO and cytokines (TNF-α, IL-1, IL-6, and IL-12) production. | Fu et al. [44] |
| Crude polysaccharides | Cytotoxicity against osteosarcoma S180 cells—10–160 µg/mL | Induce cell death and DNA fragmentation; modulate the expression of apoptosis associated genes and proteins (increase bcl-2 and decrease cdk4 and p53); activate caspase-3. | Zhong et al. [45] |
| A triple helical polysaccharide (PD3) | Mouse sarcoma S180 cells—0.2, 0.5 or 1 mg/mL | No direct cytotoxicity against S-180 cells. | Deng et al. [15] |
| Water-soluble polysaccharide—purified by gel chromatography (Sephadex G-200)—β-glucan mainly consist of glucose (98.58%). | Antioxidant assay—25–1000 µg/mL | Display reducing power; OH., O2− and DPPH radical scavenging activity. | Deng et al. [21] |
| Water extract and crude polysaccharides: | Antioxidant assay; HeLa and HepG2 cells —100–600 µg/ml | Scavenge ABTS+ and OH· radicals; inhibit lipid peroxidation; inhibit cancer cell proliferation. | Li et al. [46] |
| Six polysaccharides sub fractions with molecular weight range of 801–4656 kDa. | Antioxidant assay—1–5 mg/mL | The smallest MW 801 kDa, exhibit the most potent scavenging effect against the DPPH, OH., and O2− radicals—IC50 values 0.11, 1.02 and 0.64 mg/mL respectively. | Ker et al. [1] |
| Dictyoquinazols A, B, and C | Primary cultured mouse cortical neurons—5 µM | Protect from glutamate- and NMDA-induced excitotoxicity. | Lee et al. [26] |
| Dictyophorines A and B, and a known compound, teucrenone | Astroglial cells—5 µM | Stimulate NGF synthesis and release. | Kawagishi et al. [24] |
| Homogeneous polysaccharides and a conjugated polysaccharide fraction—Fucomammogalactan (T-3-Ad), conjugated polysaccharide fraction (T-2-A), β-(1→6)-branched -(1→3)-β-D-glucans (T-4-N and T-5-N); Partially O-acetylated 1→3)-α-D-mannans (T-2-HN and T-3-M’). | C3H/He spleen cells—concentration expressed as 10 µg/well | T-3-Ad and T-2-A exhibit significant mitogenic and CSF-inducing activities; T-4-N but not T-5-N showed both mitogenic and CSF-inducing effects. T-2-HN and T-3-M did not show activity. | Hara et al. [47] |
| Preparation | Experimental Model | Key Findings | References |
|---|---|---|---|
| Water-extractable polysaccharides | High fat-induced obesity in mice—400 mg/kg, p.o. for 45 days | Reduce the serum level of TC, TG and LDL-C while enhancing HDL-C level; suppress the obesity-induced raised activities of ALT, AST, ALP, LDH and CK enzymes; improve the liver and renal antioxidant status (increase SOD, GPx, CAT and T-AOC contents/activities and suppress MDA, LPO and MPO contents; reduce hepatic lipid levels (TC, TG and NEFA); reverses the obesity associated increase in urea and creatinine or reduced albumin; ameliorate the obesity associated increased insulin and leptin and suppressed adiponectin level; restored morphological changes of the kidney and liver (histopathological study). | Wang et al. [14] |
| Acid-extractable polysaccharides -average molecular weight of estimated to be 7.89 × 105 Da, 4.64 × 105 Da and 6.41 × 105 Da | High fat-induced obesity in mice—400 mg/kg, p.o. for 45 days | Data similar as above (Wang et al., 2019a [1]) | Wang et al. [35] |
| Alkali (NaOH solution as 0.5 mol/L, 1:10, w/v at 85 °C for 5 h) or enzyme (snailase solution as 4%, 1:4, w/v) at 38 °C for 4 h) extractable polysaccharides | High fat-induced hyperlipidemia and obesity in mice—100 or 400 mg/kg, p.o. for 33 days | Reduce body weight gain; decrease the serum levels of TC, TG, LDL-C and atherogenic index; increase HDL-C in serum; improve hepatic lipid levels (TC, TG and NEFA); decrease serum enzyme activities levels of liver toxicity marker enzymes (ALT, AST, ALP, LDH, CK and TBIL; reverse the decreased antioxidant enzyme activities (SOD, GPx and CAT), reduce non-enzymatic antioxidant capacity (T-AOC), as well as increased lipid product contents (MDA and LPO); improve hepatocyte morphology (histopathological observation); reverse the hyperlipidemia-induced decrease in adiponectin level and the increases in insulin and leptin in serum; lower improve the increased blood glucose level under the OGTT. | Wang et al. [48] |
| Polysaccharides—homogeneous β-(1→3)-D-glucan with side branches of β-(1→6)-glucosyl units | DSS-induced colitis in mice—25, 50 or 100 mg/kg p.o. for 7 days | Attenuate colitis severity (colonic length and macroscopic features, tissue architecture and inflammation score); reduce splenomegaly; suppress intestinal oxidative stress (suppress MDA while increasing GSH level; enhance HO-1 protein expression level); suppress the expression of inflammatory cytokines (TNF-α, IL-6 and IL-1β, IL-18 gene expression) and MPO activity in colonic tissues; abolish the induction of protein expressions for NLRP3, phosphorylated (p)-STAT3 and p-IκBα; ameliorate the induce apoptosis and deletion (increased Bcl2 while reducing Bax protein expressions) of tight junction protein (TJP-1 protein expression) in colonic tissues; down-regulate M1 macrophage (F4/80+CD11b+ cells) polarization or increase he M2 (F4/80+CD206+ cells) subsets in splenic tissues. | Wang et al. [4] |
| Crude polysaccharides—total sugar content was 96.66% with 13.2% polysaccharide yield. Glc 59.84%, Man 23.55% and Gal 12.95% | DSS-induced colitis in mice—10 or 33 mg/kg, p.o. for two weeks before and during the DSS | Abolish clinical symptoms—recovery of body weight loss and disease markers (stool consistency and rectal bleeding, colon length, inflammatory score); improve colonic histological changes; enhance mucins and tight junction proteins (increased expression level of claudin-1, occludin, and zonula occludins (ZO-1, and ZO-2)) expression (assessed by histopathological studies); reduce the MPO and NO; enhance SOD levels in colonic mucosa; reduce the production of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, IFN-γ) and IL-17 while enhancing the anti-inflammatory cytokines (IL-4, IL-10) in colonic mucosa; decrease the phosphorylation of p65, IκB-α and ERK; suppress the expression of iNOS; enhance alpha diversity indices; reverse the dysbiosis pattern—the DSS-induced decrease in abundance of Firmicutes and increase Proteobacteria bacteria. | Kanwal et al. [49] |
| Crude polysaccharide | Dysbiosis-induced by broad-spectrum antibiotics (clindamycin metronidazole) in BALB/c mice—0.2 mg/0.2 mL) p.o. | Restore body weight loss; restore the reduced bacterial diversity—increase Lactobacillaceae, Ruminococaceae, S24-7, and Odoribacteraceae while reducing Bacteroidaceae, Enterococcaceae), and Enterobacteriaceae; ameliorate colon wall damage and inflammation; downregulate TNF-α, IL-1β, and IL-6 (protein levels); reduce endotoxins; increase the expression of tight-junction proteins (claudin, occludin, and ZO-1); restore mucus layer thickness (modulate mucin-2 protein expression). | Kanwal et al. [7] |
| Acidic polysaccharide isolated by anion-exchange chromatography | Wild-type C. elegans under parquet-induced oxidative stress and transgenic C. elegans of neuroprotection—1.0 and 2.0 mg/mL | Decrease ROS and MDA levels and increase SOD activity; restore the functional parameters of mitochondria, (membrane potential and ATP content) in parquet-stressed nematodes; effect dependent on stress response transcription factor DAF-16/FOXO; reduce ROS levels and alleviate chemosensory behavior dysfunction in transgenic C. elegans mediated by polyglutamine and amyloid-β protein. | Zhang et al. [19] |
| A triple helical polysaccharide (PD3) | Tumor cell suspension from ascitogenous sarcoma S180 mice injected subcutaneously at subaxile position in to KM mice—100 mg/kg or 200 mg/kg i.p. for 10 days. | Dose-dependent tumour suppression; enhance body weight; upregulate the level of IL-2, IL-6, and TNF-α—increase the serum cytokine levels in tumor-bearing mice. | Deng et al. [15] |
| Acid (I) and alkali (II) extractible polysaccharides; I comprised of Glc, Fru and Man, whereas II was of Glc and Fru. Glc was the dominant monosaccharide in both (molar percentage of >60%). | Mice—1–21 g/kg, i.g. for 10 days | DIPs I and II could not enhance the cell-mediated immunity and stimulate T cell formation; both enhance macrophages phagocytosis; DIP-II enhance NK cells killing activity; I increase the weight of thymus organ phagocytosis of monocyte; II could restore delayed-type hypersensitivity reaction to dinitrofluorobenzene (DNFB); II also improve natural killer cells activity and splenocytes proliferation. | Hua et al. [6] |
| Polysaccharides I and II (as above, Ref. [1]): I composed of →1)-Glc-(6→: →1)-Man-(3, 6→ with the ratio of 5.6:1.0, while II was composed of →1)-Glc-(6→: →1)-Man-(3,6→: →1)-Xyl-(5→: →1)-Gal-(3→: →1)-Gal-(6→: with the ratio of 4.9: 15.5: 7.8: 1.0: 5.7. | d-Galactose induced senescence in mice—2.7, 5.4 or 16.2 mL/kg. | Increase SOD and GPx activities | Hua et al. [22] |
| 5-(hydroxymethyl)-2-furfural from the methanol extract | Mushroom tyrosinase assay—oxidation of L-DOPA | Dose dependent inhibition with EC50 value of 0.98 mM; noncompetitive inhibitor | Sharma et al. [29] |
| Partially O-acetylated α-D-mannan—T-2-HN | Carrageenan-induced edema and scalded edematous hyperalgesia in rats’ hind paws | Display anti-inflammatory effect in both models | Ukai et al. [50] |
| T4N and T5N: two water soluble glucans: β-(1→3)-D-glucan with side branches of β-(1→6)-glucosyl units; and T2HN as a (1→3)-α-D-mannopyranosyl residue that contain O-acetyl group | Sarcoma S180 tumour bearing mice— 5, 10 or 25 mg/kg i.p. for 10 days | T4N and T5N showed antitumor activity at 5 and 10 mg/kg; T2HN showed activity at 25 mg/kg. | Ukai et al. [51] |
| 1→6)-branched (1→3)-β-d-glucan (T-5-N), isolated from a sodium hydroxide extract—triple-helical structure in neutral or slightly alkaline solution | Carrageenan-induced edema and scalded edematous hyperalgesia in rats’ hind paws—25 mg/kg i.p. | Anti-inflammatory effect | Hara et al. [52] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Habtemariam, S. The Chemistry, Pharmacology and Therapeutic Potential of the Edible Mushroom Dictyophora indusiata (Vent ex. Pers.) Fischer (Synn. Phallus indusiatus). Biomedicines 2019, 7, 98. https://doi.org/10.3390/biomedicines7040098
Habtemariam S. The Chemistry, Pharmacology and Therapeutic Potential of the Edible Mushroom Dictyophora indusiata (Vent ex. Pers.) Fischer (Synn. Phallus indusiatus). Biomedicines. 2019; 7(4):98. https://doi.org/10.3390/biomedicines7040098
Chicago/Turabian StyleHabtemariam, Solomon. 2019. "The Chemistry, Pharmacology and Therapeutic Potential of the Edible Mushroom Dictyophora indusiata (Vent ex. Pers.) Fischer (Synn. Phallus indusiatus)" Biomedicines 7, no. 4: 98. https://doi.org/10.3390/biomedicines7040098
APA StyleHabtemariam, S. (2019). The Chemistry, Pharmacology and Therapeutic Potential of the Edible Mushroom Dictyophora indusiata (Vent ex. Pers.) Fischer (Synn. Phallus indusiatus). Biomedicines, 7(4), 98. https://doi.org/10.3390/biomedicines7040098