CRP Is Transported by Monocytes and Monocyte-Derived Exosomes in the Blood of Patients with Coronary Artery Disease

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Experimental Section
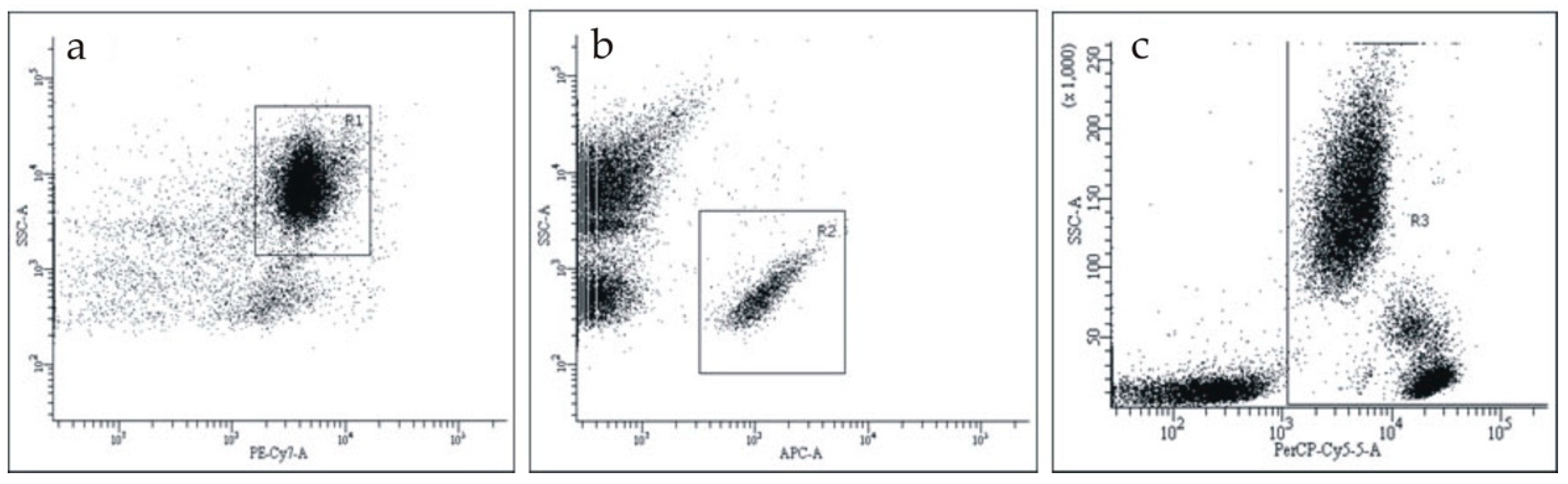
2.1. Blood Samples Collection
2.2. The Antibody Panel for Detection of mCRP and pCRP Forms
2.3. The Antibody Panels for Flow Cytometry
2.4. Staining, Detection, and Counting of Cells and Microparticles in Blood Samples
2.5. Monocyte Culture Preparation
2.6. HepG2 Culture Preparation
2.7. Real-Time Polymerase Chain Reaction (RT-PCR)
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eisenhardt, S.U.; Thiele, J.R.; Bannasch, H.; Stark, G.B.; Peter, K. C-reactive protein: How conformational changes influence inflammatory properties. Cell Cycle 2009, 8, 3885–3892. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Potempa, L.A.; El Kebir, D.; Filep, J.G. C-reactive protein and inflammation: Conformational changes affect function. Biol. Chem. 2015, 396, 1181–1197. [Google Scholar] [CrossRef] [PubMed]
- Thiele, J.R.; Zeller, J.; Bannasch, H.; Stark, G.B.; Peter, K.; Eisenhardt, S.U. Targeting C-Reactive Protein in Inflammatory Disease by Preventing Conformational Changes. Mediat. Inflamm. 2015, 2015, 372432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.R.; Wu, Y.; Zhu, L.; Potempa, L.A.; Sheng, F.L.; Lu, W.; Zhao, J. Cell membranes and liposomes dissociate C-reactive protein (CRP) to form a new, biologically active structural intermediate: mCRP(m). FASEB J. 2007, 21, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Khreiss, T.; József, L.; Potempa, L.A.; Filep, J.G. Opposing effects of C-reactive protein forms on shear-induced neutrophil-platelet adhesion and neutrophil aggregation in whole blood. Circulation 2004, 110, 2713–2720. [Google Scholar] [CrossRef]
- Ji, S.R.; Ma, L.; Bai, C.J.; Shi, J.M.; Li, H.Y.; Potempa, L.A.; Filep, J.G.; Zhao, J.; Wu, Y. Monomeric C-reactive protein activates endothelial cells via interaction with lipid raft microdomains. FASEB J. 2009, 23, 1806–1816. [Google Scholar] [CrossRef]
- Khreiss, T.; József, L.; Potempa, L.A.; Filep, J.G. Loss of pentameric symmetry in C-reactive protein induces interleukin-8 secretion through peroxynitrite signaling in human neutrophils. Circ. Res. 2005, 97, 690–697. [Google Scholar] [CrossRef] [Green Version]
- Slevin, M.; Krupinski, J. A role for monomeric C-reactive protein in regulation of angiogenesis, endothelial cell inflammation and thrombus formation in cardiovascular/cerebrovascular disease? Histol. Histopathol. 2009, 24, 1473–1478. [Google Scholar] [CrossRef]
- Badimon, L.; Peña, E.; Arderiu, G.; Padró, T.; Slevin, M.; Vilahur, G.; Chiva-Blanch, G. C-Reactive Protein in Atherothrombosis and Angiogenesis. Front. Immunol. 2018, 9, 430. [Google Scholar] [CrossRef]
- Peña, E.; de la Torre, R.; Arderiu, G.; Slevin, M.; Badimon, L. mCRP triggers angiogenesis by inducing F3 transcription and TF signalling in microvascular endothelial cells. Thromb. Haemost. 2017, 117, 357–370. [Google Scholar] [CrossRef]
- Molins, B.; Peña, E.; Vilahur, G.; Mendieta, C.; Slevin, M.; Badimon, L. C-reactive protein forms differ in their effects on thrombus growth. Arter. Thromb. Vasc. Biol. 2008, 28, 2239–2246. [Google Scholar] [CrossRef] [Green Version]
- Eisenhardt, S.U.; Habersberger, J.; Murphy, A.; Chen, Y.C.; Woollard, K.J.; Bassler, N.; Qian, H.; von Zur Muhlen, C.; Hagemeyer, C.E.; Ahrens, I.; et al. Dissociation of pentameric to monomeric C-reactive protein on activated platelets localizes inflammation to atherosclerotic plaques. Circ. Res. 2009, 105, 128–137. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, M.; Hamoud, S.; Tendler, Y.; Meilin, E.; Lazarovitch, A.; Nitecki, S.; Hayek, T. A significant correlation between C-Reactive protein levels in blood monocytes derived macrophages versus content in carotid atherosclerotic lesions. J. Inflamm. 2014, 11, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Q.; Wright, J.R. Expression of C-reactive protein by alveolar macrophages. J. Immunol. 1996, 156, 4815–4820. [Google Scholar] [PubMed]
- Kolb-Bachofen, V.; Puchta-Teudt, N.; Egenhofer, C. Expression of membrane-associated C-reactive protein by human monocytes: Indications for a selectin-like activity participating in adhesion. Glycoconj. J. 1995, 12, 122–127. [Google Scholar] [CrossRef]
- Ciubotaru, I.; Potempa, L.A.; Wander, R.C. Production of monomeric C-reactive protein in U937-derived macrophages. Exp. Biol. Med. 2005, 230, 762–770. [Google Scholar] [CrossRef] [Green Version]
- Habersberger, J.; Strang, F.; Scheichl, A.; Htun, N.; Bassler, N.; Merivirta, R.M.; Diehl, P.; Krippner, G.; Meikle, P.; Eisenhardt, S.U.; et al. Circulating microparticles generate and transport monomeric C-reactive protein in patients with myocardial infarction. Cardiovasc. Res. 2012, 96, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Crawford, J.R.; Trial, J.; Nambi, V.; Hoogeveen, R.C.; Taffet, G.E.; Entman, M.L. Plasma Levels of Endothelial Microparticles Bearing Monomeric C-reactive Protein are Increased in Peripheral Artery Disease. J. Cardiovasc. Transl. Res. 2016, 9, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Basso, M.; Bonetto, V. Extracellular Vesicles and a Novel Form of Communication in the Brain. Front. Neurosci. 2016, 10, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Tang, B.; Liu, X.; Wu, X.; Wang, H.; Xu, D.; Guo, Y. Increased monomeric CRP levels in acute myocardial infarction: A possible new and specific biomarker for diagnosis and severity assessment of disease. Atherosclerosis 2015, 239, 343–349. [Google Scholar] [CrossRef]
- Kiefer, C.R.; Stock, R.E.; Flanagan, S.S.; Darling, C.E.; Smith, C.S.; Snyder, L.M. Early verification of myocardial ischemia with a novel biomarker of acute tissue damage: C-reactive protein fractional forms. Clin. Chim. Acta 2012, 413, 1536–1541. [Google Scholar] [CrossRef] [PubMed]
- Sadallah, S.; Eken, C.; Schifferli, J.A. Ectosomes as immunomodulators. Semin. Immunopathol. 2011, 33, 487–495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, Z.L.; Combes, V.; Grau, G.E.; King, N.J. Microparticles as immune regulators in infectious disease—An opinion. Front. Immunol. 2011, 2, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, A.P., 3rd; Mackman, N. Microparticles in hemostasis and thrombosis. Circ. Res. 2011, 108, 1284–1297. [Google Scholar] [CrossRef] [PubMed]
- Pepys, M.B.; Hirschfield, G.M.; Tennent, G.A.; Gallimore, J.R.; Kahan, M.C.; Bellotti, V.; Hawkins, P.N.; Myers, R.M.; Smith, M.D.; Polara, A.; et al. Targeting C-reactive protein for the treatment of cardiovascular disease. Nature 2006, 440, 1217–1221. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, L.C.P.; Janiszewski, M.; Pontieri, V.; de Almeida Pedro, M.; Bassi, E.; Tucci, P.J.F.; Laurindo, F.R.M. Platelet-derived exosomes from septic shock patients induce myocardial dysfunction. Crit. Care. 2007, 11, R120. [Google Scholar] [CrossRef] [Green Version]
- Paganelli, F.; Gaudry, M.; Ruf, J.; Guieu, R. Recent Advances in the Role of the Adenosinergic System in Coronary Artery Disease. Cardiovasc. Res. 2020, cvaa275. [Google Scholar] [CrossRef]
- Ruf, J.; Vairo, D.; Paganelli, F.; Guieu, R. Extracellular vesicles with ubiquitinated adenosine A 2A receptor in plasma of patients with coronary artery disease. J. Cell Mol. Med. 2019, 23, 6805–6811. [Google Scholar] [CrossRef] [Green Version]
- Deharo, P.; Marlinge, M.; Guiol, C.; Vairo, D.; Fromonot, J.; Mace, P.; Chefrour, M.; Gastaldi, M.; Bruzzese, L.; Gaubert, M.; et al. Homocysteine concentration and adenosine A 2A receptor production by peripheral blood mononuclear cells in coronary artery disease patients. J. Cell Mol. Med. 2020, 24, 8942–8949. [Google Scholar] [CrossRef]
- Melnikov, I.; Chumachenko, P.; Kozlov, S.; Majorova, A.; Saburova, O.; Portnaya, T.; Osidak, M.; Domogatsky, S.; Buryachkovskaya, L.; Gabbasov, Z. Monomeric C-reactive protein and local inflammatory response in patients with stable coronary artery disease. Atherosclerosis 2018, 275, e105. [Google Scholar] [CrossRef]
- Haider, D.G.; Leuchten, N.; Schaller, G.; Gouya, G.; Kolodjaschna, J.; Schmetterer, L.; Kapiotis, S.; Wolzt, M. C-reactive protein is expressed and secreted by peripheral blood mononuclear cells. Clin. Exp. Immunol. 2006, 146, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Kuta, A.E.; Baum, L.L. C-reactive protein is produced by a small number of normal human peripheral blood lymphocytes. J. Exp. Med. 1986, 164, 321–326. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CRP-Positive Cells, % | |
|---|---|
| CD235a-positive erythrocytes | 4.3 ± 1.6 |
| CD41-positive platelets | 3.5 ± 1.3 |
| CD45-positive leukocytes | 26.5 ± 9.1 |
| mCRP-Positive MPs, % | pCRP-Positive MPs, % | p * | |
|---|---|---|---|
| Erythrocyte-derived MPs | 21.1 ± 9.8 | 64.1 ± 20.4 | p > 0.05 |
| Platelet-derived MPs | 0.4 ± 0.3 | 0.5 ± 0.2 | p > 0.05 |
| Leukocyte-derived MPs | 47.9 ± 6.2 | 12.9 ± 2.3 | p < 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melnikov, I.; Kozlov, S.; Saburova, O.; Zubkova, E.; Guseva, O.; Domogatsky, S.; Arefieva, T.; Radyukhina, N.; Zvereva, M.; Avtaeva, Y.; et al. CRP Is Transported by Monocytes and Monocyte-Derived Exosomes in the Blood of Patients with Coronary Artery Disease. Biomedicines 2020, 8, 435. https://doi.org/10.3390/biomedicines8100435
Melnikov I, Kozlov S, Saburova O, Zubkova E, Guseva O, Domogatsky S, Arefieva T, Radyukhina N, Zvereva M, Avtaeva Y, et al. CRP Is Transported by Monocytes and Monocyte-Derived Exosomes in the Blood of Patients with Coronary Artery Disease. Biomedicines. 2020; 8(10):435. https://doi.org/10.3390/biomedicines8100435
Chicago/Turabian StyleMelnikov, Ivan, Sergey Kozlov, Olga Saburova, Ekaterina Zubkova, Olga Guseva, Sergey Domogatsky, Tatiana Arefieva, Natalia Radyukhina, Maria Zvereva, Yuliya Avtaeva, and et al. 2020. "CRP Is Transported by Monocytes and Monocyte-Derived Exosomes in the Blood of Patients with Coronary Artery Disease" Biomedicines 8, no. 10: 435. https://doi.org/10.3390/biomedicines8100435
APA StyleMelnikov, I., Kozlov, S., Saburova, O., Zubkova, E., Guseva, O., Domogatsky, S., Arefieva, T., Radyukhina, N., Zvereva, M., Avtaeva, Y., Buryachkovskaya, L., & Gabbasov, Z. (2020). CRP Is Transported by Monocytes and Monocyte-Derived Exosomes in the Blood of Patients with Coronary Artery Disease. Biomedicines, 8(10), 435. https://doi.org/10.3390/biomedicines8100435