Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains

 ,
,  ,
,  ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Kefir Grains Isolation
2.2. Isolation of LAB Strains
2.3. Bacterial Strains and Culture Conditions
2.4. Resistance to Low pH, Pepsin, Pancreatin and Tolerance to Bile Salts
2.5. Antibiotic Susceptibility
2.6. Molecular Characterisation
2.7. Human Cancer Cell Lines
2.8. Quantitative Adhesion Assay
2.9. Cell Proliferation Assay
2.10. Evaluation of Pro-Apoptotic Activity by Flow Cytometry
2.11. Assessment of Cell Cycle Progression by Flow Cytometry
2.12. Statistical Analysis
3. Results and Discussion
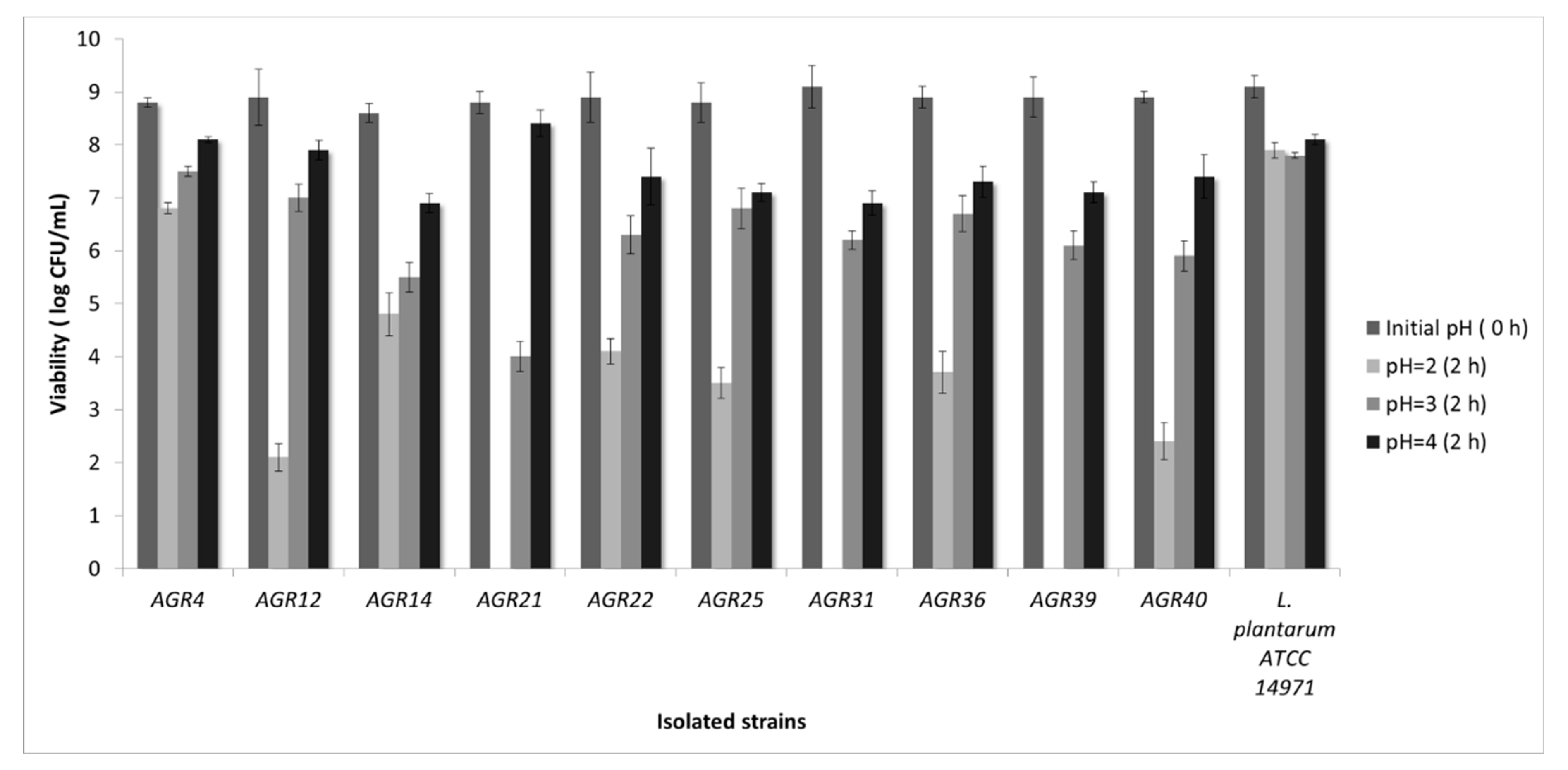
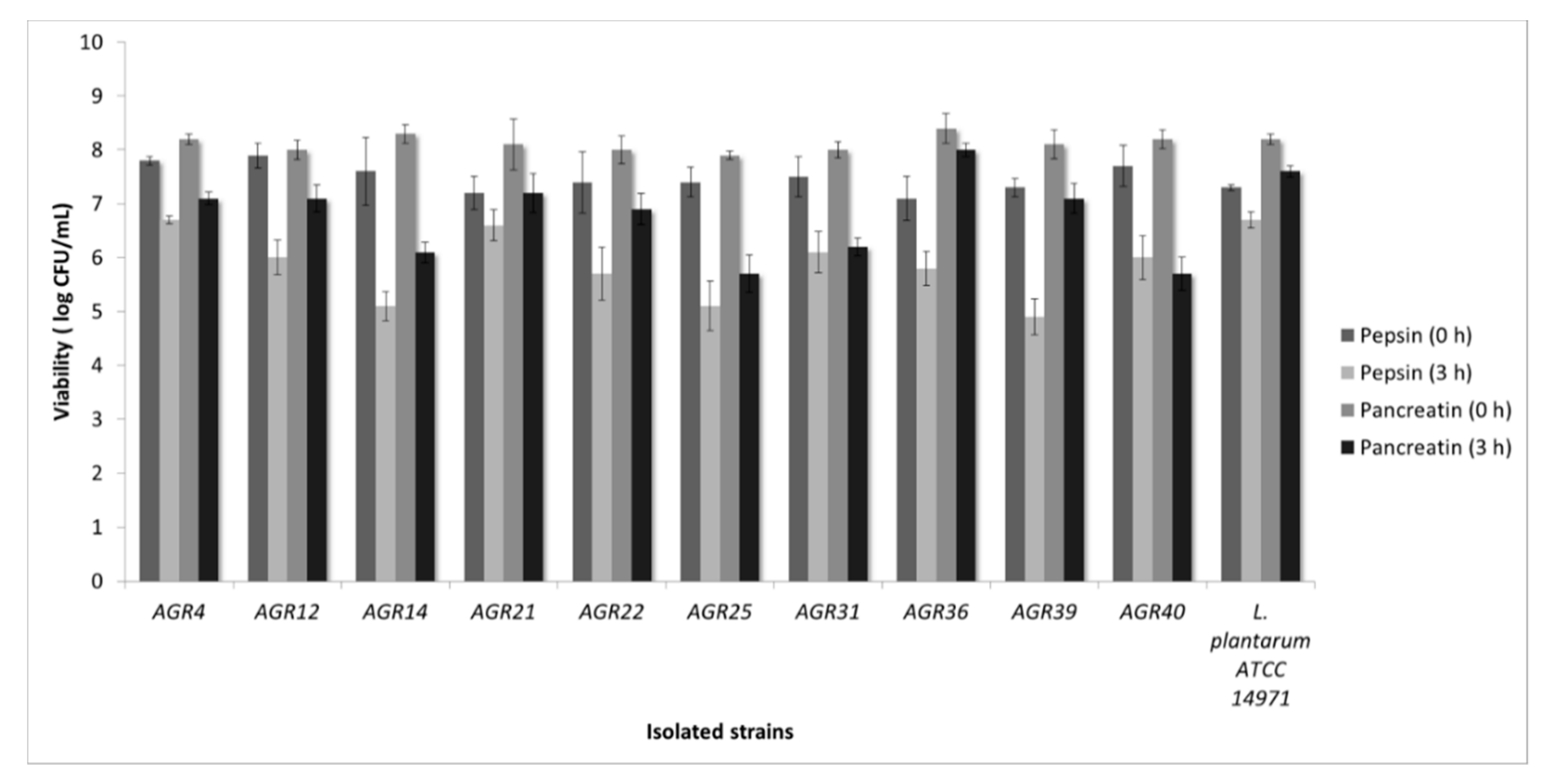
3.1. Isolation of LAB Strains from Kefir Grains and In Vitro Screening for Probiotic Properties
3.2. Safety Profile—Antibiotic Susceptibility
3.3. Molecular Characterization of Lactobacillus Strain AGR 4
3.4. Αdhesion of L. paracasei AGR 4 on Epithelial Colon Cancer Cells
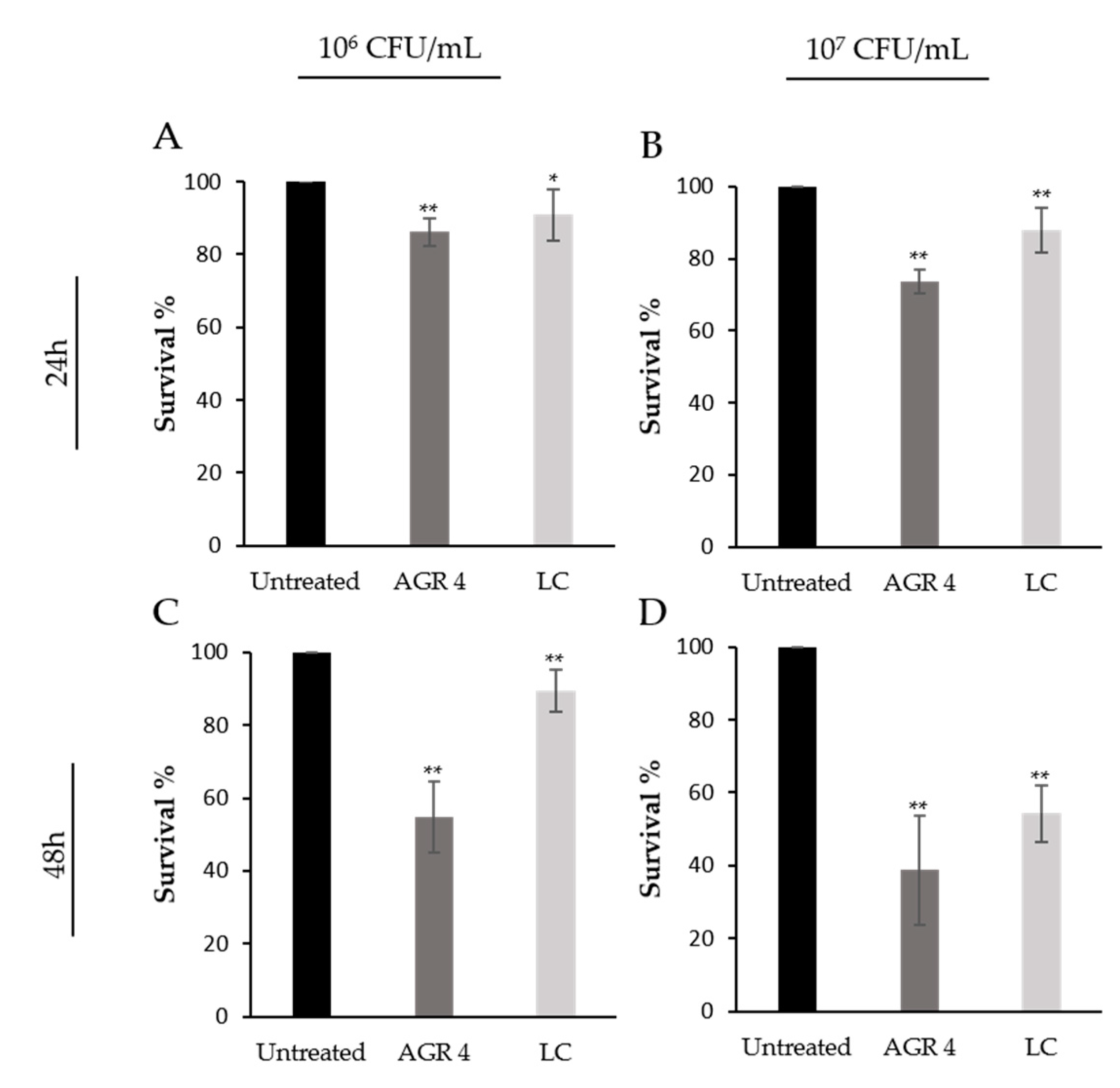
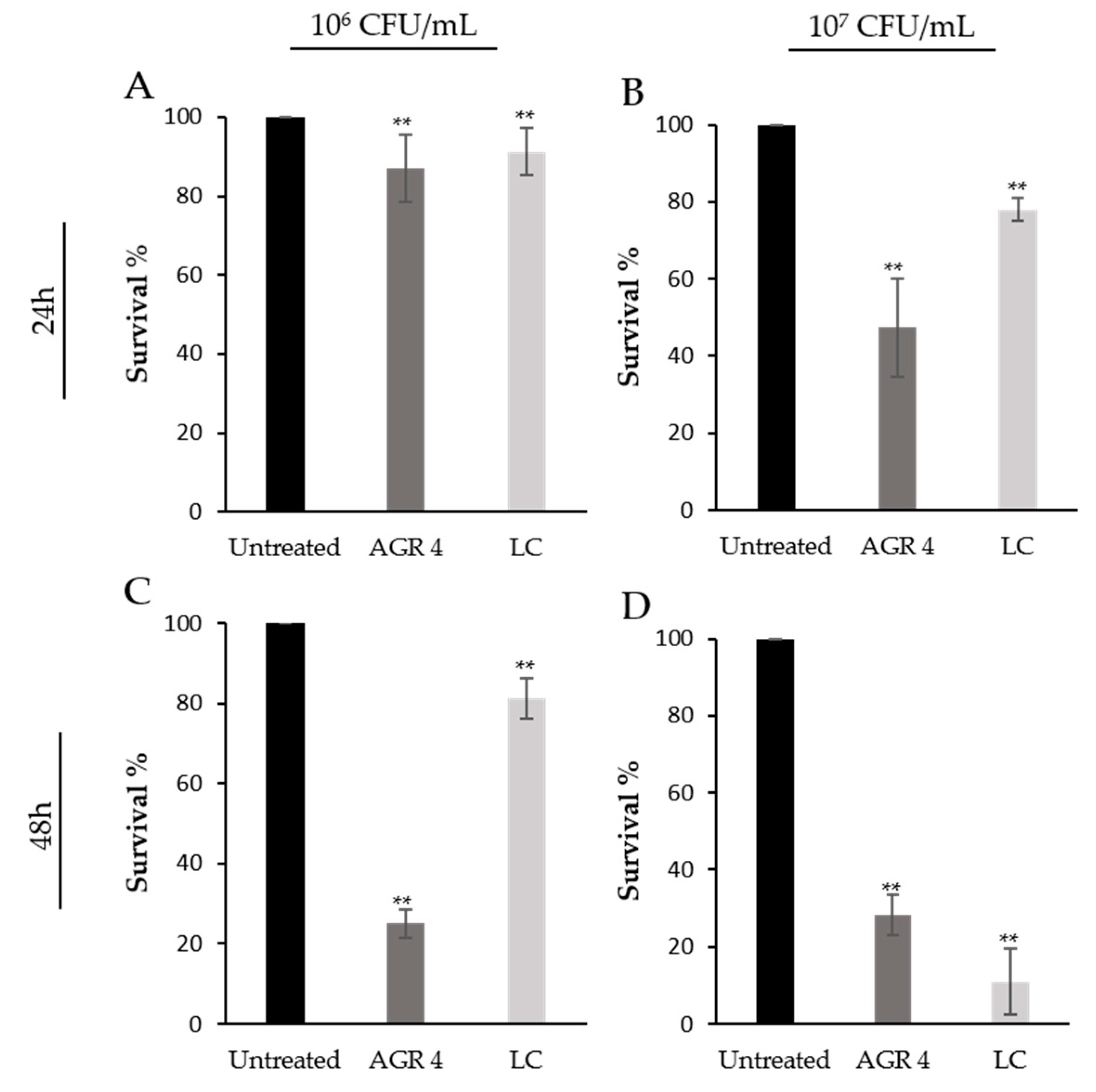
3.5. L. paracasei AGR 4 Induces Cytotoxic Effects against HT-29 and A375 Cancer Cells
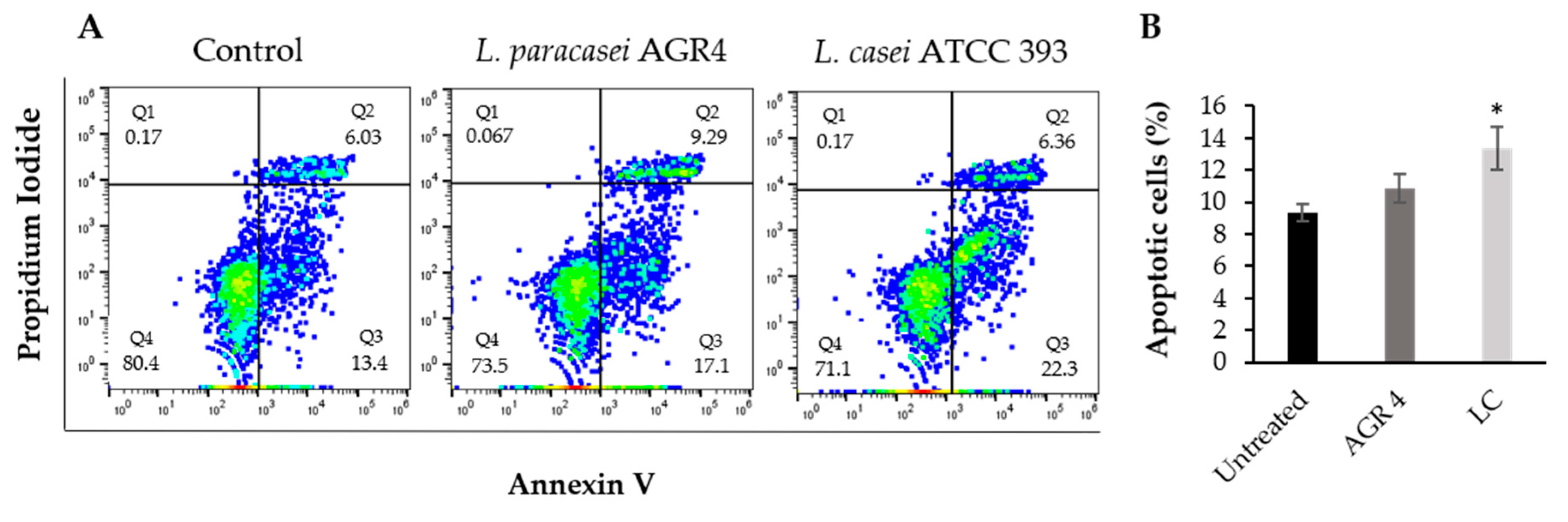
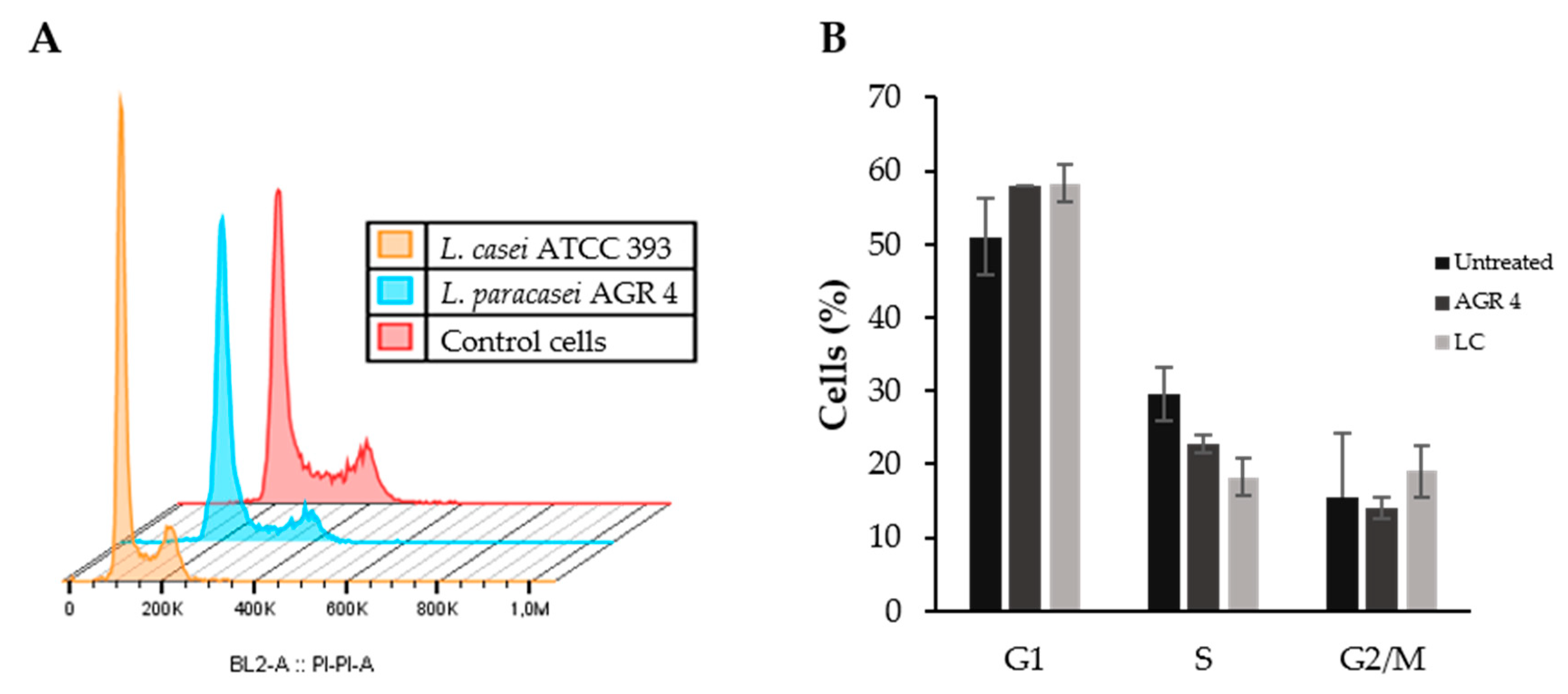
3.6. Lactobacillus Paracasei AGR 4 Does Not Induce Apoptotic Cells Death or Cell Cycle Arrest on HT-29 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations/World Health Organization (FAO/WHO). Evaluation of Health and Nutritional Properties of Powder Milk and Live Lactic Acid Bacteria. 2002. Available online: www.fao.org/3/a-a0512e.pdf (accessed on 5 December 2020).
- Islam, S.U. Clinical Uses of Probiotics. Medicine 2016, 95, 2658. [Google Scholar] [CrossRef] [PubMed]
- Kiousi, D.E.; Karapetsas, A.; Karolidou, K.; Panayiotidis, M.I.; Pappa, A.; Galanis, A. Probiotics in Extraintestinal Diseases: Current Trends and New Directions. Nutrients 2019, 11, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zmora, N.; Zilberman-Schapira, G.; Suez, J.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Kotler, E.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.; et al. Personalized Gut Mucosal Colonization Resistance to Empiric Probiotics Is Associated with Unique Host and Microbiome Features. Cell 2018, 174, 1388–1405.e21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bermudez-Brito, M.; Plaza-Díaz, J.; Muñoz-Quezada, S.; Gómez-Llorente, C.; Gil, A. Probiotic mechanisms of action. Ann. Nutr. Metab. 2012, 61, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Tuo, Y.; Song, X.; Song, Y.; Liu, W.; Tang, Y.; Gao, Y.; Jiang, S.; Qian, F.; Mu, G. Screening probiotics from Lactobacillus strains according to their abilities to inhibit pathogen adhesion and induction of pro-inflammatory cytokine IL-8. J. Dairy Sci. 2018, 101, 4822–4829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsham, A.D.S.; MacKenzie, D.A.; Cook, V.; Wemyss-Holden, S.; Hews, C.L.; Juge, N.; Schüller, S. Lactobacillus reuteri Inhibition of Enteropathogenic Escherichia coli Adherence to Human Intestinal Epithelium. Front. Microbiol. 2016, 7, 244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thirabunyanon, M.; Hongwittayakorn, P. Potential probiotic lactic acid bacteria of human origin induce antiproliferation of colon cancer cells via synergic actions in adhesion to cancer cells and short-chain fatty acid bioproduction. Appl. Biochem. Biotechnol. 2013, 169, 511–525. [Google Scholar] [CrossRef]
- Saxami, G.; Ypsilantis, P.; Sidira, M.; Simopoulos, C.; Kourkoutas, Y.; Galanis, A. Distinct adhesion of probiotic strain Lactobacillus casei ATCC 393 to rat intestinal mucosa. Anaerobe 2012, 18, 417–420. [Google Scholar] [CrossRef]
- Faghfoori, Z.; Gargari, B.P.; Gharamaleki, A.S.; Bagherpoure, H.; Khosroushahi, A.Y. Cellular and molecular mechanisms of probiotics effects on colorectal cancer. J. Funct. Foods 2015, 18, 463–472. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, X.; Zhang, L.; Xu, S.; Zhang, Q.; Lu, R. A surface-layer protein from Lactobacillus acidophilus NCFM induces autophagic death in HCT116 cells requiring ROS-mediated modulation of mTOR and JNK signaling pathways. Food Funct. 2019, 10, 4102–4112. [Google Scholar] [CrossRef]
- Tian, P.J.; Li, B.L.; Shan, Y.J.; Zhang, J.N.; Chen, J.Y.; Yu, M.; Zhang, L.W. Extraction of Peptidoglycan from L. paracasei subp. Paracasei X12 and Its Preliminary Mechanisms of Inducing Immunogenic Cell Death in HT-29 Cells. Int. J. Mol. Sci. 2015, 16, 20033–20049. [Google Scholar] [PubMed]
- Fontana, L.; Bermudez-Brito, M.; Plaza-Diaz, J.; Muñoz-Quezada, S.; Gil, A. Sources, isolation, characterisation and evaluation of probiotics. Br. J. Nutr. 2013, 2, S35–S50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Castillo, V.; Marín-Vega, A.M.; Ilabaca, A.; Albarracín, L.; Marcial, G.; Kitazawa, H.; Garcia-Cancino, A.; Villena, J. Characterization of the immunomodulatory and anti-Helicobacter pylori properties of the human gastric isolate Lactobacillus rhamnosus UCO-25A. Biofouling 2019, 35, 922–937. [Google Scholar] [CrossRef]
- Poinsot, P.; Penhoat, A.; Mitchell, M.; Sauvinet, V.; Meiller, L.; Louche-Pélissier, C.; Meugnier, E.; Ruiz, M.; Schwarzer, M.; Michalski, M.C.; et al. Probiotic from human breast milk, Lactobacillus fermentum, promotes growth in animal model of chronic malnutrition. Pediatr. Res. 2020, 88, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Rosa, D.D.; Dias, M.M.; Grześkowiak, L.M.; Reis, S.A.; Conceição, L.L.; Maria do Carmo, G.P. Milk kefir: Nutritional, microbiological and health benefits. Nutr. Res. Rev. 2017, 30, 82–96. [Google Scholar] [CrossRef] [PubMed]
- Bengoa, A.A.; Iraporda, C.; Garrote, G.L.; Abraham, A.G. Kefir micro-organisms: Their role in grain assembly and health properties of fermented milk. J. Appl. Microbiol. 2019, 126, 686–700. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; Bancalari, E.; Milanović, V.; Cardinali, F.; Osimani, A.; Sardaro, M.L.S.; Bottari, B.; Bernini, V.; Aquilanti, L.; Clementi, F. Study of the bacterial diversity of foods: PCR-DGGE versus LH-PCR. Int. J. Food Microbiol. 2017, 242, 24–36. [Google Scholar] [CrossRef]
- Vardjan, T.; Lorbeg, P.M.; Rogelj, I.; Majhenič, A.Č. Characterization and stability of lactobacilli and yeast microbiota in kefir grains. J. Dairy Sci. 2013, 96, 2729–2736. [Google Scholar] [CrossRef] [Green Version]
- Plessas, S.; Nouska, C.; Karapetsas, A.; Kazakos, S.; Alexopoulos, A.; Mantzourani, I.; Chondrou, P.; Fournomiti, M.; Galanis, A.; Bezirtzoglou, E. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from Feta-type cheese. Food Chem. 2017, 226, 102–108. [Google Scholar] [CrossRef]
- Mantzourani, I.; Terpou, A.; Alexopoulos, A.; Chondrou, P.; Galanis, A.; Bekatorou, A.; Bezirtzoglou, E.; Koutinas, A.A.; Plessas, S. Application of A Novel Potential Probiotic Lactobacillus paracasei Strain Isolated from Kefir Grains in the Production of Feta-Type Cheese. Microorganisms 2018, 6, 121. [Google Scholar] [CrossRef] [Green Version]
- Mantzourani, I.; Chondrou, P.; Bontsidis, C.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Galanis, A.; Plessas, S. Assessment of the probiotic potential of lactic acid bacteria isolated from kefir grains: Evaluation of adhesion and antiproliferative properties in in vitro experimental systems. Ann. Microbiol. 2019, 69, 751–763. [Google Scholar]
- Charteris, W.P.; Kelly, P.M.; Morelli, L.; Collins, J.K. Antibiotic susceptibility of potentially probiotic Lactobacillus species. J. Food Protect. 1998, 61, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Klijn, N.; Weerkamp, A.H.; de Vos, W.M. Identification of mesophilic lactic acid bacteria by using polymerase chain reaction-amplified variable regions of 16S rRNA and specific DNA probes. Appl. Env. Microbiol. 1991, 57, 3390–3393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventura, M.; Canchaya, C.; Meylan, V.; Klaenhammer, T.R.; Zink, R. Analysis, characterization, and loci of the tuf genes in Lactobacillus and Bifidobacterium species and their direct application for species identification. Appl. Env. Microbiol. 2003, 69, 6908–6922. [Google Scholar] [CrossRef] [Green Version]
- Argyri, A.A.; Zoumpopoulou, G.; Karatzas, K.A.; Tsakalidou, E.; Nychas, G.J.; Panagou, E.Z.; Tassou, C.C. Selection of potential probiotic lactic acid bacteria from fermented olives by in vitro tests. Food Microbiol. 2013, 33, 282–291. [Google Scholar] [CrossRef]
- Chondrou, P.; Karapetsas, A.; Kiousi, D.E.; Tsela, D.; Tiptiri-Kourpeti, A.; Anestopoulos, I.; Kotsianidis, I.; Bezirtzoglou, E.; Pappa, A.; Galanis, A. Lactobacillus paracasei K5 displays adhesion, anti-proliferative activity and apoptotic effects in human colon cancer cells. Benef. Microbes 2018, 9, 975–983. [Google Scholar] [CrossRef]
- Sabokbar, N.; Khodaiyan, F. Characterization of pomegranate juice and whey based novel beverage fermented by kefir grains. J. Food Sci. Technol. 2015, 52, 3711–3718. [Google Scholar] [CrossRef] [Green Version]
- Bengoa, A.A.; Llamas, M.G.; Iraporda, C.; Dueñas, M.T.; Abraham, A.G.; Garrote, G.L. Impact of growth temperature on exopolysaccharide production and probiotic properties of Lactobacillus paracasei strains isolated from kefir grains. Food Microbiol. 2018, 69, 212–218. [Google Scholar] [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP). Guidance on the assessment of bacterial susceptibility to antimicrobials of human and veterinary importance. EFSA J. 2012, 10, 2740. [Google Scholar]
- Danielsen, M.; Wind, A. Susceptibility of Lactobacillus spp. to antimicrobial agents. Int. J. Food Microbiol. 2003, 82, 1–11. [Google Scholar] [CrossRef]
- Kastner, S.; Perreten, V.; Bleuler, H.; Hugenschmidt, G.; Lacroix, C.; Meile, L. Antibiotic susceptibility patterns and resistance genes of starter cultures and probiotic bacteria used in food. Syst. Appl. Microbiol. 2006, 29, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Drago, L.; Mattina, R.; De Vecchi, E.; Toscano, M. Phenotypic and genotypic antibiotic resistance in some probiotics proposed for medical use. Int. J. Antimicrob. Agents. 2013, 41, 396–397. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sidira, M.; Kourkoutas, Y.; Kanellaki, M.; Charalampopoulos, D. In vitro study on the cell adhesion ability of immobilized lactobacilli on natural supports. Food Res. Int. 2015, 76, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.J.; Kim, K.T.; Kim, T.Y.; Paik, H.D. Probiotic Properties and Antioxidant Activities of Pediococcus pentosaceus SC28 and Levilactobacillus brevis KU15151 in Fermented Black Gamju. Foods 2020, 9, 1154. [Google Scholar] [CrossRef]
- Vidhyasagar, V.; Jeevaratnam, K. Evaluation of Pediococcus pentosaceus strains isolated from Idly batter for probiotic properties in vitro. J. Funct. Foods 2013, 5, 235–243. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Ben-Zeev Brik, R.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e16. [Google Scholar] [CrossRef] [Green Version]
- Tiptiri-Kourpeti, A.; Spyridopoulou, K.; Santarmaki, V.; Aindelis, G.; Tompoulidou, E.; Lamprianidou, E.E.; Saxami, G.; Ypsilantis, P.; Lampri, E.S.; Simopoulos, C.; et al. Lactobacillus casei Exerts Anti-Proliferative Effects Accompanied by Apoptotic Cell Death and Up-Regulation of TRAIL in Colon Carcinoma Cells. PLoS ONE 2016, 11, e0147960. [Google Scholar] [CrossRef]
- Saxami, G.; Karapetsas, A.; Lamprianidou, E.; Kotsianidis, I.; Chlichlia, A.; Tassou, C.; Zoumpourlis, V.; Galanis, A. Two potential probiotic lactobacillus strains isolated from olive microbiota exhibit adhesion and anti-proliferative effects in cancer cell lines. J. Funct. Foods 2016, 24, 461–471. [Google Scholar] [CrossRef]
- Aindelis, G.; Tiptiri-Kourpeti, A.; Lampri, E.; Spyridopoulou, K.; Lamprianidou, E.; Kotsianidis, I.; Ypsilantis, P.; Pappa, A.; Chlichlia, K. Immune Responses Raised in an Experimental Colon Carcinoma Model Following Oral Administration of Lactobacillus casei. Cancers 2020, 12, 368. [Google Scholar] [CrossRef] [Green Version]
- Gianotti, L.; Morelli, L.; Galbiati, F.; Rocchetti, S.; Coppola, S.; Beneduce, A.; Gilardini, C.; Zonenschain, D.; Nespoli, A.; Braga, M. A randomized double-blind trial on perioperative administration of probiotics in colorectal cancer patients. World J. Gastroenterol. 2010, 16, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Mego, M.; Chovanec, J.; Vochyanova-Andrezalova, I.; Konkolovsky, P.; Mikulova, M.; Reckova, M.; Miskovska, V.; Bystricky, B.; Beniak, J.; Medvecova, L.; et al. Prevention of irinotecan induced diarrhea by probiotics: A randomized double blind, placebo controlled pilot study. Complement. Ther. Med. 2015, 23, 356–362. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota-host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Patra, V.; Gallais Sérézal, I.; Wolf, P. Potential of Skin Microbiome, Pro- and/or Pre-Biotics to Affect Local Cutaneous Responses to UV Exposure. Nutrients 2020, 12, 1795. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Kwon, M.; Lee, J.; Park, S.; Seo, J.; Roh, S. Anti-Cancer Effects of Lactobacillus plantarum L-14 Cell-Free Extract on Human Malignant Melanoma A375 Cells. Molecules 2020, 25, 3895. [Google Scholar] [CrossRef]
- Wang, T.; Zheng, N.; Luo, Q.; Jiang, L.; He, B.; Yuan, X.; Shen, L. Probiotics Lactobacillus reuteri Abrogates Immune Checkpoint Blockade-Associated Colitis by Inhibiting Group 3 Innate Lymphoid Cells. Front. Immunol. 2019, 10, 1235. [Google Scholar] [CrossRef]
- Shi, L.; Sheng, J.; Chen, G.; Zhu, P.; Shi, C.; Li, B.; Park, C.; Wang, J.; Zhang, B.; Liu, Z.; et al. Combining IL-2-based immunotherapy with commensal probiotics produces enhanced antitumor immune response and tumor clearance. J. Immunother. Cancer 2020, 8, e000973. [Google Scholar]
- Alizadeh, S.; Esmaeili, A.; Omidi, Y. Anti-cancer properties of Escherichia coli Nissle 1917 against HT-29 colon cancer cells through regulation of Bax/Bcl-xL and AKT/PTEN signaling pathways. Iran. J. Basic Med. Sci. 2020, 23, 886–893. [Google Scholar]
- Ghanavati, R.; Asadollahi, P.; Shapourabadi, M.B.; Razavi, S.; Talebi, M.; Rohani, M. Inhibitory effects of Lactobacilli cocktail on HT-29 colon carcinoma cells growth and modulation of the Notch and Wnt/β-catenin signaling pathways. Microb. Pathog. 2020, 139, 103829. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Oh, S.; Yun, H.S.; Oh, S.; Kim, S.H. Cell-bound exopolysaccharide from probiotic bacteria induces autophagic cell death of tumour cells. Lett. Appl. Microbiol. 2010, 51, 123–130. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agent | AGR 4 | AGR 12 | AGR 14 | AGR 21 | AGR 22 | AGR 25 | AGR 31 | AGR 36 | AGR 39 | AGR 40 | L. plantarum ATCC 14917 | Cut-Off † |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Minimum Inhibitory Concentration μg/mL) | ||||||||||||
| AMX | 3.48 ± 0.43 a | 4.20 ± 0.26 b | 5.05 ± 0.25 de | 4.98 ± 0.21 cde | 4.08 ± 0.31 b | 4.85 ± 0.29 de | 3.42 ± 0.21 a | 4.41 ± 0.55 bc | 4.56 ± 0.41 bcd | 5.11 ± 0.47 e | 4.55 ± 0.21 bcde | n.r. ‡* |
| AMC | 1.61 ± 0.31 de | 1.05 ± 0.3 ab | 1.18 ± 0.13 bc | 1.62 ± 0.22 de | 1.22 ± 0.11 bc | 1.65 ± 0.19 de | 1.38 ± 0.21 cd | 0.76 ± 0.07 a | 1.81 ± 0.09 ef | 0.95 ± 0.08 ab | 2.08 ± 0.21 f | n.r. ‡* |
| AMP | 1.04 ± 0.21 ab | 1.47 ± 0.18 d | 2.13 ± 0.12 ef | 2.38 ± 0.19 f | 1.13 ± 0.21 abc | 2.08 ± 0.11 e | 1.28 ± 0.09 bcd | 1.38 ± 0.09 c d | 1.52 ± 0.08 d | 0.95 ± 0.11 a | 1.21 ± 0.12 abc | 4 ‡ |
| CLI | 0.49 ± 0.05 a | 1.08 ± 0.14 bc | 0.79 ± 0.08 ab | 1.27 ± 0.09 cd | 0.89 ± 0.08 b | 1.13 ± 0.09 bc | 0.91 ± 0.11 b | 1.08 ± 0.19 bc | 0.85 ± 0.14 b | 1.49 ± 0.08 d | 0.85 ± 0.09 b | 1 ‡ |
| ERY | 0.59 ± 0.12 a | 1.08 ± 0.08 bcd | 1.06 ± 0.13 bc | 1.89 ± 0.07 e | 0.72 ± 0.09 a | 0.59 ± 0.07 a | 1.04 ± 0.08 bc | 1.29 ± 0.19 cd | 1.11 ± 0.09 bcd | 1.49 ± 0.29 de | 1.15 ± 0.21 cd | 1 ‡ |
| GEN | 4.08 ± 2.12 a | 4.74 ± 0.21 a | 5.19 ± 0.08 a | 5.27 ± 0.11 a | 4.32 ± 0.28 a | 4.09 ± 0.14 a | 4.59 ± 0.37 a | 5.29 ± 0.07 a | 4.81 ± 0.31 a | 4.98 ± 0.23 a | 4.29 ± 0.09 a | 32 ‡ |
| MDL | 189.7 ± 47.4 a | 148.0 ± 21.8 a | 195.3 ± 21.0 a | 201.2 ± 18.7 a | 195.2 ± 11.4 a | 200.9 ± 24.5 a | 195.1 ± 17.9 a | 198.2 ± 18.7 a | 200.8 ± 21.8 a | 199.8 ± 12.1 a | 199.1 ± 11.0 a | n.r. ‡* |
| TET | 1.24 ± 0.35 a | 3.41 ± 0.31 cd | 3.08 ± 0.28 bcd | 3.11 ± 0.33 bd | 4.08 ± 0.31 cde | 3.08 ± 0.43 bcd | 1.99 ± 0.18 ab | 2.48 ± 0.13 bc | 3.15 ± 0.23 cd | 4.85 ± 0.31 e | 4.15 ± 0.95 de | 4‡ |
| TGC | 0.48 ± 0.13 a | 0.58 ± 0.11 a | 0.52 ± 0.04 a | 0.52 ± 0.17 a | 0.58 ± 0.11 a | 0.53 ± 0.05 a | 0.57 ± 0.14 a | 0.61 ± 0.08 a | 0.42 ± 0.09 a | 0.65 ± 0.08 a | 0.71 ± 0.08 a | n.r. ‡* |
| VAN | >256 | >256 | >256 | >256 | >256 | >256 | >256 | >256 | >256 | >256 | >256 | n.r. ‡* |
| LAB Strains | Adhesion Ability (%) |
|---|---|
| Lactobacillus AGR 4 | 6.1 ± 0.14 |
| L. casei ATCC 393 | 4.8 ± 0.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plessas, S.; Kiousi, D.E.; Rathosi, M.; Alexopoulos, A.; Kourkoutas, Y.; Mantzourani, I.; Galanis, A.; Bezirtzoglou, E. Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains. Biomedicines 2020, 8, 594. https://doi.org/10.3390/biomedicines8120594
Plessas S, Kiousi DE, Rathosi M, Alexopoulos A, Kourkoutas Y, Mantzourani I, Galanis A, Bezirtzoglou E. Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains. Biomedicines. 2020; 8(12):594. https://doi.org/10.3390/biomedicines8120594
Chicago/Turabian StylePlessas, Stavros, Despoina Eugenia Kiousi, Marina Rathosi, Athanasios Alexopoulos, Yiannis Kourkoutas, Ioanna Mantzourani, Alex Galanis, and Eugenia Bezirtzoglou. 2020. "Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains" Biomedicines 8, no. 12: 594. https://doi.org/10.3390/biomedicines8120594
APA StylePlessas, S., Kiousi, D. E., Rathosi, M., Alexopoulos, A., Kourkoutas, Y., Mantzourani, I., Galanis, A., & Bezirtzoglou, E. (2020). Isolation of a Lactobacillus paracasei Strain with Probiotic Attributes from Kefir Grains. Biomedicines, 8(12), 594. https://doi.org/10.3390/biomedicines8120594