Asthma and Obesity in Children



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Asthma Endotypes
3.2. Obesity and Asthma
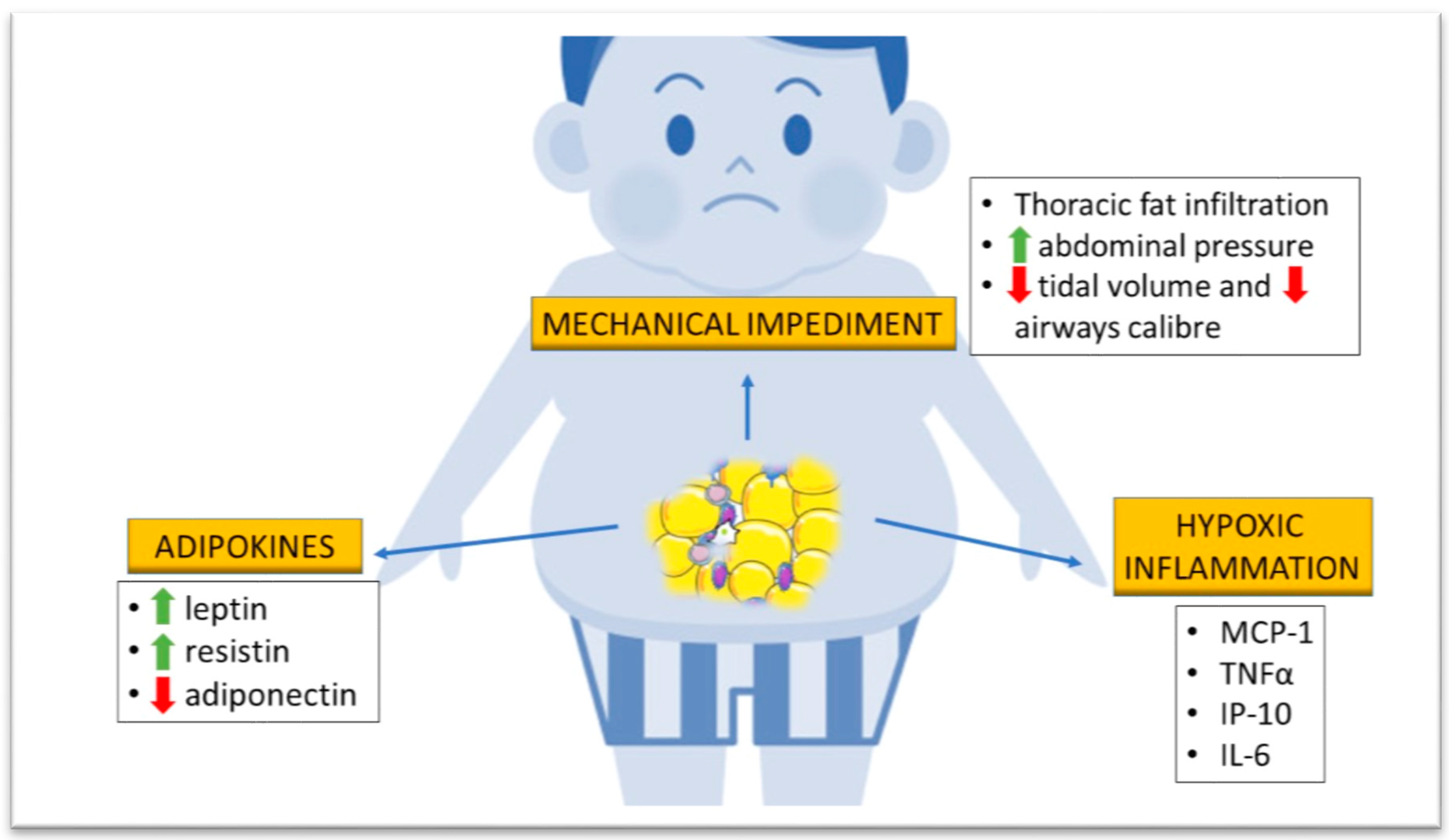
3.2.1. Mechanical Impediment
3.2.2. Adipokine Production
3.2.3. Hypoxic Inflammation
3.2.4. Gut Microbiota
4. Common Obesity-Asthma Predisposition
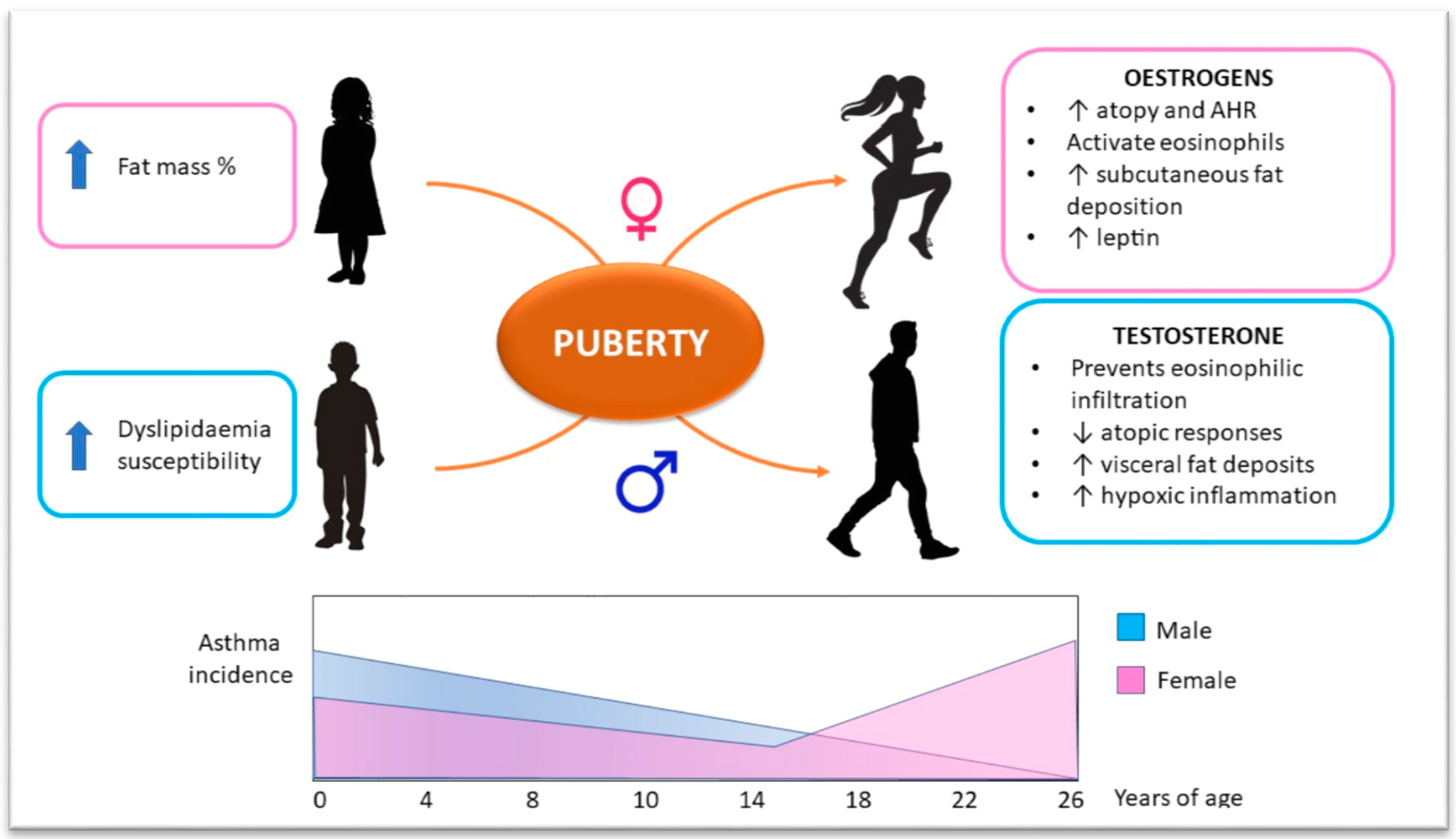
5. Sex Hormone Influence on Obesity–Asthma Link
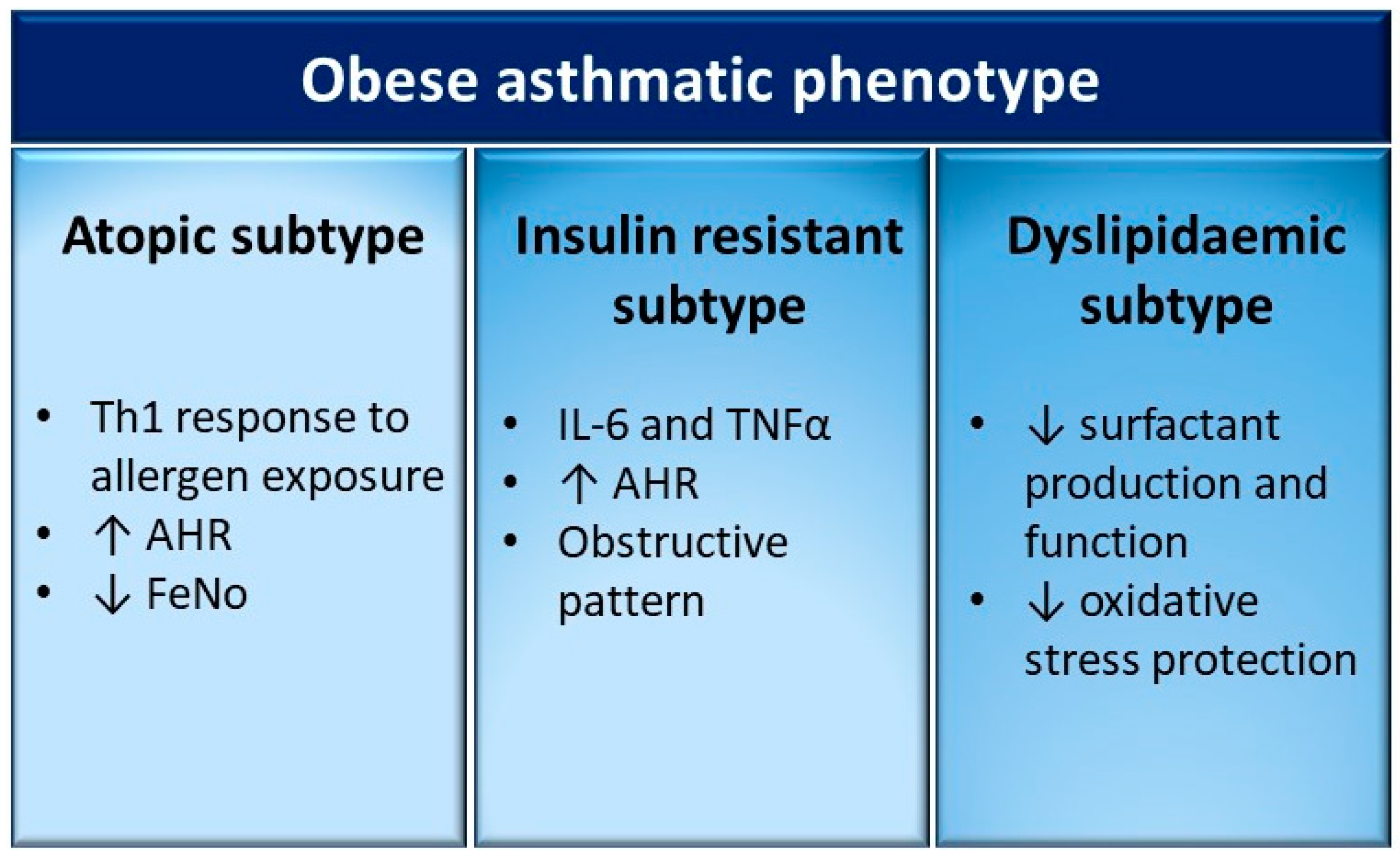
6. Different Clinical Subtypes of the Obese-Asthmatic Phenotype in Children
6.1. Obese-Asthmatic Phenotype and Atopy
6.2. Obese-Asthmatic Phenotype and Insulin Resistance
6.3. Obese-Asthmatic Phenotype and Dyslipidaemia
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Pearce, N.; Aït-Khaled, N.; Beasley, R.; Mallol, J.; Keil, U.; Mitchell, E.A.; Robertson, C.; Anderson, H.R.; Asher, M.I.; Björkstén, B.; et al. Worldwide trends in the prevalence of asthma symptoms: Phase III of the International Study of Asthma and Allergies in Childhood (ISAAC). Thorax 2007, 62, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Di Cesare, M.; Sorić, M.; Bovet, P.; Miranda, J.J.; Bhutta, Z.; Stevens, G.A.; Laxmaiah, A.; Kengne, A.P.; Bentham, J. The epidemiological burden of obesity in childhood: A worldwide epidemic requiring urgent action. BMC Med. 2019, 17, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, W.J.; Phipatanakul, W. Difficult-to-control asthma: Epidemiology and its link with environmental factors. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshi, M.; Cardinal, J.; Hill, R.J.; Davies, P.S.W.; Wainwright, C. Asthma and insulin resistance in children. Respirology 2010, 15, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Cottrell, L.; Neal, W.A.; Ice, C.; Perez, M.K.; Piedimonte, G. Metabolic abnormalities in children with asthma. Am. J. Respir. Crit. Care Med. 2011, 183, 441–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuruvilla, M.E.; Lee, F.E.H.; Lee, G.B. Understanding Asthma Phenotypes, Endotypes, and Mechanisms of Disease. Clin. Rev. Allergy Immunol. 2019, 56, 219–233. [Google Scholar] [CrossRef]
- Moffatt, M.F.; Gut, I.G.; Demenais, F.; Strachan, D.P.; Bouzigon, E.; Heath, S.; Mutius, E.; Farrall, M.; Large-Scale, L.M.A. A large-scale, consortium-based genomewide association study of asthma. N. Engl. J. Med. 2010, 363, 1211–1221. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Ge, M.Q.; Kokalari, B.; Redai, I.G.; Wang, X.; Kemeny, D.M.; Bhandoola, A.H.A. Group 2 innate lymphoid cells mediate ozone-induced airway inflammation and hyperresponsiveness in mice. J. Allergy Clin. Immunol. 2016, 137, 571–578. [Google Scholar] [CrossRef] [Green Version]
- Durrani, S.R.; Viswanathan, R.K.; Busse, W.W. What effect does asthma treatment have on airway remodeling? Current perspectives. J. Allergy Clin. Immunol. 2011, 128, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Chung, K. Airway smooth muscle cells: Contributing to and regulating airway mucosal inflammation? Eur. Respir. J. 2000, 15, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Pillai, S.; Abbas, A.; Lichtman, A. Cellular and Molecular Immunology, 6th ed.; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Chen, Y.C.; Dong, G.H.; Lin, K.C.; Lee, Y.L. Gender difference of childhood overweight and obesity in predicting the risk of incident asthma: A systematic review and meta-analysis. Obes. Rev. 2013, 14, 222–231. [Google Scholar] [CrossRef]
- Deng, X.; Ma, J.; Yuan, Y.; Zhang, Z.; Niu, W. Association between overweight or obesity and the risk for childhood asthma and wheeze: An updated meta-analysis on 18 articles and 73 252 children. Pediatr. Obes. 2019, 14. [Google Scholar] [CrossRef]
- Hammer, L.D.; Kraemer, H.C.; Wilson, D.M.; Ritter, P.L.; Dornbusch, S.M. Standardized percentile curves of body-mass index for children and adolescents. Am. J. Dis. Child. 1991, 145, 259–263. [Google Scholar] [CrossRef]
- Guibas, G.V.; Manios, Y.; Xepapadaki, P.; Moschonis, G.; Douladiris, N.; Mavrogianni, C.; Papadopoulos, N.G. The obesity-asthma link in different ages and the role of Body Mass Index in its investigation: Findings from the Genesis and Healthy Growth Studies. Allergy Eur. J. Allergy Clin. Immunol. 2013, 68, 1298–1305. [Google Scholar] [CrossRef]
- Ashwell, M.; Hsieh, S.D. Six reasons why the waist-to-height ratio is a rapid and effective global indicator for health risks of obesity and how its use could simplify the international public health message on obesity. Int. J. Food Sci. Nutr. 2005, 56, 303–307. [Google Scholar] [CrossRef]
- Sánchez Jiménez, J.; Herrero Espinet, F.J.; Mengibar Garrido, J.M.; Roca Antonio, J.; Peños Mayor, S.; Peñas Boira, M.d.M.; Roca Comas, A.; Ballester Martínez, A. Asthma and insulin resistance in obese children and adolescents. Pediatr. Allergy Immunol. 2014, 25, 699–705. [Google Scholar] [CrossRef]
- Kuschnir, F.C.; Felix, M.M.R.; Kuschnir, M.C.; Bloch, K.V.; Azevedo de Oliveira Costa Jordão, E.; Solé, D.; Ledo Alves da Cunha, A.J.; Szklo, M. Severe asthma is associated with metabolic syndrome in Brazilian adolescents. J. Allergy Clin. Immunol. 2018, 141, 1947–1949.e4. [Google Scholar] [CrossRef] [PubMed]
- Lang, J.E.; Bunnell, H.T.; Lima, J.J.; Hossain, M.J.; Wysocki, T.; Bacharier, L.; Dempsey, A.; Ulrich, L.; Test, M.R.; Forrest, C.B. Effects of age, sex, race/ethnicity, and allergy status in obesity-related pediatric asthma. Pediatr. Pulmonol. 2019, 54, 1684–1693. [Google Scholar] [CrossRef] [PubMed]
- Almqvist, C.; Worm, M.; Leynaert, B. Impact of gender on asthma in childhood and adolescence: A GA 2LEN review. Allergy Eur. J. Allergy Clin. Immunol. 2008, 63, 47–57. [Google Scholar] [CrossRef]
- Moore, W.C.; Meyers, D.A.; Wenzel, S.E.; Teague, W.G.; Li, H.; Li, X.; D’Agostino, R.; Castro, M.; Curran-Everett, D.; Fitzpatrick, A.M.; et al. Identification of asthma phenotypes using cluster analysis in the severe asthma research program. Am. J. Respir. Crit. Care Med. 2010, 181, 315–323. [Google Scholar] [CrossRef]
- Beuther, D.A.; Weiss, S.T.; Sutherland, E.R. Obesity and asthma. Am. J. Respir. Crit. Care Med. 2006, 174, 112–119. [Google Scholar] [CrossRef]
- Shore, S.A. Obesity and asthma: Lessons from animal models. J. Appl. Physiol. 2007, 102, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Davidson, W.J.; Mackenzie-Rife, K.A.; Witmans, M.B.; Montgomery, M.D.; Ball, G.D.C.; Egbogah, S.; Eves, N.D. Obesity negatively impacts lung function in children and adolescents. Pediatr. Pulmonol. 2014, 49, 1003–1010. [Google Scholar] [CrossRef]
- Gomez-Llorente, M.A.; Romero, R.; Chueca, N.; Martinez-Cañavate, A.; Gomez-Llorente, C. Obesity and asthma: A missing link. Int. J. Mol. Sci. 2017, 18, 1490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sood, A.; Shore, S.A. Adiponectin, Leptin and Resistin in Asthma: Basic Mechanisms through Population Studies. J. Allergy 2013, 2013, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Guler, N.; Kirerleri, E.; Ones, U.; Tamay, Z.; Salmayenli, N.; Darendeliler, F. Leptin: Does it have any role in childhood asthma? J. Allergy Clin. Immunol. 2004, 114, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, D.; Canfield, S.M.; Andrade, A.; Isasi, C.R.; Hall, C.B.; Rubinstein, A.; Arens, R. Obesity-associated asthma in children a distinct entity. Chest 2012, 141, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Shore, S.A.; Terry, R.D.; Flynt, L.; Xu, A.; Hug, C. Adiponectin attenuates allergen-induced airway inflammation and hyperresponsiveness in mice. J. Allergy Clin. Immunol. 2006, 118, 389–395. [Google Scholar] [CrossRef]
- Ionescu, L.I.; Alphonse, R.S.; Arizmendi, N.; Morgan, B.; Abel, M.; Eaton, F.; Duszyk, M.; Vliagoftis, H.; Aprahamian, T.R.; Walsh, K.; et al. Airway delivery of soluble factors from plastic-adherent bone marrow cells prevents murine asthma. Am. J. Respir. Cell Mol. Biol. 2012, 46, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Shin, Y.H.; Lee, K.E.; Kim, E.S.; Sohn, M.H.; Kim, K.E. Relationship between adipokines and manifestations of childhood asthma. Pediatr. Allergy Immunol. 2008, 19, 535–540. [Google Scholar] [CrossRef]
- Ziora, D.; Machura, E.; Ziora, K.T.; Swietochowska, E.; Oswiecimska, J.M.; Kasperska-Zajac, A. Serum resistin levels are elevated in schoolchildren with atopic asthma. Neuro Endocrinol. Lett. 2013, 34, 212–216. [Google Scholar]
- Rastogi, D.; Fraser, S.; Oh, J.; Huber, A.M.; Schulman, Y.; Bhagtani, R.H.; Khan, Z.S.; Tesfa, L.; Hall, C.B.; Macian, F. Inflammation, metabolic dysregulation, and pulmonary function among obese urban adolescents with asthma. Am. J. Respir. Crit. Care Med. 2015, 191, 149–160. [Google Scholar] [CrossRef] [Green Version]
- Scott, H.A.; Gibson, P.G.; Garg, M.L.; Wood, L.G. Airway inflammation is augmented by obesity and fatty acids in asthma. Eur. Respir. J. 2011, 38, 594–602. [Google Scholar] [CrossRef] [Green Version]
- Ravussin, Y.; Koren, O.; Spor, A.; LeDuc, C.; Gutman, R.; Stombaugh, J.; Knight, R.; Ley, R.E.; Leibel, R.L. Responses of gut microbiota to diet composition and weight loss in lean and obese mice. Obesity 2012, 20, 738–747. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Bäckhed, F.; Fulton, L.; Gordon, J.I. Diet-induced obesity is linked to marked but reversible alterations in the mouse distal gut microbiome. Cell Host Microbe 2008, 3, 213–223. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Zhu, Z.; Anttila, V.; Smoller, J.W.; Lee, P.H. Statistical power and utility of metaanalysis methods for cross-phenotype genome-wide association studies. PLoS ONE 2018, 13, e0193256. [Google Scholar] [CrossRef] [Green Version]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor. Science (80-) 2010, 328, 228–231. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. MetaHIT Consortium. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the gut microbiota on host adiposity are modulated by the shortchain fatty-acid binding G protein–coupled receptor, Gpr. Proc. Natl. Acad. Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nature 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Shore, S.A.; Cho, Y. Obesity and asthma: Microbiome-metabolome interactions. Am. J. Respir. Cell Mol. Biol. 2016, 54, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Comhair, S.A.; McDunn, J.; Bennett, C.; Fettig, J.; Erzurum, S.C.; Kalhan, S.C. Metabolomic endotype of asthma. J. Immunol. 2015, 195, 643–650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcik, W.; Boutin, R.C.T.; Sokolowska, M.; Finlay, B.B. The Role of Lung and Gut Microbiota in the Pathology of Asthma. Immunity 2020, 52, 241–255. [Google Scholar] [CrossRef] [Green Version]
- Hallstrand, T.S.; Fischer, M.E.; Wurfel, M.M.; Afari, N.; Buchwald, D.; Goldberg, J. Genetic pleiotropy between asthma and obesity in a community-based sample of twins. J. Allergy Clin. Immunol. 2005, 116, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Guo, Y.; Shi, H.; Liu, C.L.; Panganiban, R.A.; Chung, W.; O’Connor, L.J.; Himes, B.E.; Gazal, S.; Hasegawa, K.; et al. Shared genetic and experimental links between obesity-related traits and asthma subtypes in UK Biobank. J. Allergy Clin. Immunol. 2020, 145, 537–549. [Google Scholar] [CrossRef] [Green Version]
- McGillick, E.V.; Lock, M.C.; Orgeig, S.; Morrison, J.L. Maternal obesity mediated predisposition to respiratory complications at birth and in later life: Understanding the implications of the obesogenic intrauterine environment. Paediatr. Respir. Rev. 2017, 21, 11–18. [Google Scholar] [CrossRef] [PubMed]
- WHO. Report of the Commission of Ending Childhood Obesity; WHO Press: Geneva, Switzerland, 2016. [Google Scholar]
- Rastogi, D.; Suzuki, M.; Greally, J.M. Differential epigenome-wide DNA methylation patterns in childhood obesity-associated asthma. Sci. Rep. 2013, 3, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaheen, S.O.; Sterne, J.A.C.; Montgomery, S.M.; Azima, H. Birth weight, body mass index and asthma in young adults. Thorax 1999, 54, 396–402. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Dales, R.; Krewski, D.; Breithaupt, K. Increased effects of smoking and obesity on asthma among female Canadians: The National Population Health Survey, 1994–1995. Am. J. Epidemiol. 1999, 150, 255–262. [Google Scholar] [CrossRef]
- Castro-Rodríguez, J.A.; Holberg, C.J.; Morgan, W.J.; Wright, A.L.; Martinez, F.D. Increased incidence of asthmalike symptoms in girls who become overweight or obese during the school years. Am. J. Respir. Crit. Care Med. 2001, 163, 1344–1349. [Google Scholar] [CrossRef]
- Hamano, N.; Terada, N.; Maesako, K.; Hohki, G.; Ito, T.; Yamashita, T.; Konno, A. Effect of Female Hormones on the Production of IL-4 and IL-13 From Peripheral Blood Mononuclear Cells. Acta Otolaryngol. Suppl. 1998, 537, 27–31. [Google Scholar]
- Hamano, N.; Terada, N.; Maesako, K.; Numata, T.; Konno, A. Effect of sex hormones on eosinophilic inflammation in nasal mucosa. Allergy Asthma Proc. 1998, 19, 263–269. [Google Scholar] [CrossRef]
- Hellings, P.W.; Vandekerckhove, P.; Claeys, R.; Billen, J.; Kasran, A.; Ceuppens, J.L. Progesterone increases airway eosinophilia and hyper-responsiveness in a murine model of allergic asthma. Clin. Exp. Allergy 2003, 33, 1457–1463. [Google Scholar] [CrossRef]
- Murray, C.S.; Canoy, D.; Buchan, I.; Woodcock, A.; Simpson, A.; Custovic, A. Body mass index in young children and allergic disease: Gender differences in a longitudinal study. Clin. Exp. Allergy 2011, 41, 78–85. [Google Scholar] [CrossRef]
- Gilliland, F.D.; Berhane, K.; Islam, T.; McConnell, R.; Gauderman, W.J.; Gilliland, S.S.; Avol, E.; Peters, J.M. Obesity and the risk of newly diagnosed asthma in school-age children. Am. J. Epidemiol. 2003, 158, 406–415. [Google Scholar] [CrossRef] [Green Version]
- Ahmadizar, F.; Vijverberg, S.J.H.; Arets, H.G.M.; De Boer, A.; Lang, J.E.; Kattan, M.; Palmer, C.N.A.; Mukhopadhyay, S.; Turner, S.; Der Zee, A.H.M.V. Childhood obesity in relation to poor asthma control and exacerbation: A meta-analysis. Eur. Respir. J. 2016, 48, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Tung, K.Y.; Tsai, C.H.; Su, M.W.; Wang, P.C.; Chen, C.H.; Lee, Y.L. Lipid profiles in children with and without asthma: Interaction of asthma and obesity on hyperlipidemia. Diabetes Metab. Syndr. Clin. Res. Rev. 2013, 7, 20–25. [Google Scholar] [CrossRef]
- Visness, C.M.; London, S.J.; Daniels, J.L.; Kaufman, J.S.; Yeatts, K.B.; Siega-Riz, A.M.; Liu, A.H.; Calatroni, A.; Zeldin, D.C. Association of obesity with IgE levels and allergy symptoms in children and adolescents: Results from the National Health and Nutrition Examination Survey 2005–2006. J. Allergy Clin. Immunol. 2009, 123, 1163–1169.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, R.A.; Zhu, M.; Rivera-Sanchez, Y.M.; Lu, F.L.; Theman, T.A.; Flynt, L. Allergic airway responses in obese mice. Am. J. Respir. Crit. Care Med. 2007, 176, 650–658. [Google Scholar] [CrossRef] [Green Version]
- Von Mutius, E.; Schwartz, J.; Neas, L.M.; Dockery, D.; Weiss, S.T. Relation of body mass index to asthma and atopy in children: The National Health and Nutrition Examination Study III. Thorax 2001, 56, 835–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, D.; Chinn, S.; Potts, J.; Burney, P. Association of body mass index with respiratory symptoms and atopy: Results from the European Community Respiratory Health Survey. Clin. Exp. Allergy. 2002, 32, 831–837. [Google Scholar] [CrossRef]
- Taveras, E.M.; Rifas-Shiman, S.L.; Camargo, C.A., Jr.; Gold, D.R.; Litonjua, A.A.; Oken, E.; Weiss, S.T.; Gillman, M.W. Higher adiposity in infancy associated with recurrent wheeze in a prospective cohort of children. J. Allergy Clin. Immunol. 2008, 121, 1161–1166. [Google Scholar] [CrossRef] [Green Version]
- Di Filippo, P.; Scaparrotta, A.; Rapino, D.; De Giorgis, T.; Petrosino, M.I.; Attanasi, M.; Di Pillo, S.; Chiarelli, F.; Mohn, A. Insulin resistance and lung function in obese asthmatic pre-pubertal children. J. Pediatr. Endocrinol. Metab. 2018, 31, 45–51. [Google Scholar] [CrossRef]
- Han, Y.Y.; Forno, E.; Celedón, J.C. Adiposity, fractional exhaled nitric oxide, and asthma in U.S. Children. Am. J. Respir. Crit. Care Med. 2014, 190, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Santamaria, F.; Montella, S.; De Stefano, S.; Sperlì, F.; Barbarano, F.; Spadaro, R.; Franzese, A. Asthma, atopy, and airway inflammation in obese children. J. Allergy Clin. Immunol. 2007, 120, 965–967. [Google Scholar] [CrossRef]
- Consilvio, N.P.; Di Pillo, S.; Verini, M.; De Giorgis, T.; Cingolani, A.; Chiavaroli, V.; Chiarelli, F.; Mohn, A. The reciprocal influences of asthma and obesity on lung function testing, ahr, and airway inflammation in prepubertal children. Pediatr. Pulmonol. 2010, 45, 1103–1110. [Google Scholar] [CrossRef] [Green Version]
- Kazaks, A.; Uriu-Adams, J.Y.; Stern, J.S.; Albertson, T.E.; De Winter-De Groot, K.M.; Uiterwaal, C.S.P.M.; Van Der Ent, C.K. No significant relationship between exhaled nitric oxide and body mass index in people with asthma [4] (multiple letters). J. Allergy Clin. Immunol. 2005, 116, 929–930. [Google Scholar] [CrossRef]
- Erkoçoʇlu, M.; Kaya, A.; Özcan, C.; Akan, A.; Vezir, E.; Azkur, D.; Kara, Ö.; Demirel, F.; Ginis, T.; Civelek, E.; et al. The effect of obesity on the level of fractional exhaled nitric oxide in children with asthma. Int. Arch. Allergy Immunol. 2013, 162, 156–162. [Google Scholar] [CrossRef]
- Yiallouros, P.K.; Savva, S.C.; Kolokotroni, O.; Dima, K.; Zerva, A.; Kouis, P.; Bousquet, J.; Middleton, N. Asthma: The role of low high-density-lipoprotein cholesterol in childhood and adolescence. Int. Arch. Allergy Immunol. 2014, 165, 91–99. [Google Scholar] [CrossRef]
- Cavalher-Machado, S.C.; de Lima, W.T.; Damazo, A.S.; de Frias Carvalho, V.; Martins, M.A.; Silva, P.M.R.; Sannomiya, P. Down-regulation of mast cell activation and airway reactivity in diabetic rats: Role of insulin. Eur. Respir. J. 2004, 24, 552–558. [Google Scholar] [CrossRef]
- Schaafsma, D.; McNeill, K.D.; Stelmack, G.L.; Gosens, R.; Baarsma, H.A.; Dekkers, B.G.J.; Frohwerk, E.; Penninks, J.M.; Sharma, P.; Ens, K.M.; et al. Insulin increases the expression of contractile phenotypic markers in airway smooth muscle. Am. J. Physiol. Cell Physiol. 2007, 293, 429–439. [Google Scholar] [CrossRef]
- Dekkers, B.G.J.; Schaafsma, D.; Tran, T.; Zaagsma, J.; Meurs, H. Insulin-induced laminin expression promotes a hypercontractile airway smooth muscle phenotype. Am. J. Respir. Cell Mol. Biol. 2009, 41, 494–504. [Google Scholar] [CrossRef]
- Al-Shawwa, B.A.; Al-Huniti, N.H.; DeMattia, L.; Gershan, W. Asthma and insulin resistance in morbidly obese children and adolescents. J. Asthma 2007, 44, 469–473. [Google Scholar] [CrossRef]
- Karampatakis, N.; Karampatakis, T.; Galli-Tsinopoulou, A.; Kotanidou, E.P.; Tsergouli, K.; Eboriadou-Petikopoulou, M.; Haidopoulou, K. Impaired glucose metabolism and bronchial hyperresponsiveness in obese prepubertal asthmatic children. Pediatr. Pulmonol. 2017, 52, 160–166. [Google Scholar] [CrossRef]
- Perez, M.K.; Piedimonte, G. Metabolic asthma: Is there a link between obesity, diabetes, and asthma? Immunol. Allergy Clin. N. Am. 2014, 34, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Forno, E.; Han, Y.Y.; Muzumdar, R.H.; Celedón, J.C. Insulin resistance, metabolic syndrome, and lung function in U.S. adolescents with and without asthma. J. Allergy Clin. Immunol. 2016, 136, 304–311. [Google Scholar] [CrossRef] [Green Version]
- Fang, L.J.; Huang, C.S.; Liu, Y.C.; Su, Y.M.; Wan, K.S. The lipid profile in obese asthmatic children compared to non-obese asthmatic children. Allergol. Immunopathol. 2016, 44, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Fessler, M.B.; Massing, M.W.; Spruell, B.; Jaramillo, R.; Draper, D.W.; Madenspacher, J.H.; Arbes, S.J.; Calatroni, A.; Zeldin, D.C. Novel Relationship of Serum Cholesterol with Asthma and Wheeze in the United States. J. Allergy Clin. Immunol. 2009, 124, 967–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nistor, A.; Simionescu, M. Uptake of low density lipoproteins by the hamster lung. Interactions with capillary endothelium. Am. Rev. Respir. Dis. 1986, 134, 1266–1272. [Google Scholar] [PubMed]
- Chen, C.L.; Liu, I.H.; Fliesler, S.J.; Han, X.; Huang, S.S.; San Huang, J. Cholesterol suppresses cellular TGFbeta responsiveness: Implications in atherogenesis. J. Cell Sci. 2007, 120, 3509–3521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulman, E.S.; Quinn, T.J.; Post, T.J.; O’Donnell, P.; Rodriguez, A.; Gonen, B. Low density lipoprotein (LDL) inhibits histamine release from human mast cells. Biochem. Biophys. Res. Commun. 1987, 148, 553–559. [Google Scholar] [CrossRef]
- Harrison, G.J.; Jordan, L.R.; Selley, M.L.; Willis, R.J. Low-density lipoproteins inhibit histamine and NaNO2 relaxations of the coronary vasculature and reduce contractile function in isolated rat hearts. Heart Vessel. 1995, 10, 249–257. [Google Scholar] [CrossRef]
- Alkassabany, Y.M.; Farghaly, A.G.; El-Ghitany, E.M. Prevalence, risk factors, and predictors of nonalcoholic fatty liver disease among schoolchildren: A hospital-based study in Alexandria, Egypt. Arab. J. Gastroenterol. 2014, 15, 76–81. [Google Scholar] [CrossRef]
- Mantovani, A.; Lonardo, A.; Vinco, G.; Zoppini, G.; Lippi, G.; Bonora, E.; Loomba, R.; Tilg, H.; Byrne, C.D.; Fabbri, L.; et al. Association between non-alcoholic fatty liver disease and decreased lung function in adults: A systematic review and meta-analysis. Diabetes Metab. 2019, 45, 536–544. [Google Scholar] [CrossRef]
- Peng, T.C.; Kao, T.W.; Wu, L.W.; Chen, Y.J.; Chang, Y.W.; Wang, C.C.; Tsao, Y.T.; Chen, W.L. Association between Pulmonary Function and Nonalcoholic Fatty Liver Disease in the NHANES III Study. Medicine 2015, 94, 1–6. [Google Scholar] [CrossRef]
- Hong, H.C.; Hwang, S.Y.; Choi, H.Y.; Yoo, H.J.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Baik, S.H.; Choi, D.S.; Choi, K.M. Relationship between sarcopenia and nonalcoholic fatty liver disease: The Korean Sarcopenic Obesity Study. Hepatology 2014, 59, 1772–1778. [Google Scholar] [CrossRef] [PubMed]
- Takamura, T.; Misu, H.; Ota, T.; Kaneko, S. Fatty liver as a consequence and cause of insulin resistance. Endocr. J. 2012, 59, 745–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Targher, G.; Bertolini, L.; Scala, L.; Zoppini, G.; Zenari, L.; Falezza, G. Non-alcoholic hepatic steatosis and its relation to increased plasma biomarkers of inflammation and endothelial dysfunction in non-diabetic men. Role of visceral adipose tissue. Diabet. Med. 2005, 22, 1354–1358. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cytokine | Source | Role |
|---|---|---|
| Alarmins (TSLP, IL-25, IL-33) | Epithelial cells | ILC2s and APCs activation after allergen exposure |
| IL-5 | Activated ILC2s, mast-cells and Th2 lymphocytes | Promotes eosinophil proliferation and differentiation |
| IL-4 | Th2 lymphocytes | IgE production, Th2 lymphocytes differentiation |
| IL-13 | Mainly Th2 lymphocytes | promotes fibrosis, mucus secretion, IgE production, chemokines and adhesion molecules production |
| MCP-1 (CCL2) | Activated monocytes | Leukocytes migration to inflammation site |
| IP-10 (CXCL-11) | Activated monocytes | T-CD8+ migration to inflammation site |
| TNF | Activated monocytes, others | Neutrophils migration and activation, systemic inflammatory response |
| IL-1 | Activated monocytes, others | Systemic inflammatory response |
| IFN-γ | NK cells, T-CD8+ cells, Th1 lymphocytes | Macrophages activation, Th1 differentiation, IgG production, MHC class I and II exposure |
| TGF-β | T lymphocytes, activated mononuclear monocytes | Inflammatory regulator, T-reg differentiation, IL-17 differentiation, IgA production, tissue repair |
| IL-17 | Th17 lymphocytes | Neutrophils proliferation, differentiation and migration, induces IL-1, TNF and chemokines production |
| IL-6 | Activated monocytes and T-lymphocytes | Promotes cellular-mediated response (↑ IL-17) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sansone, F.; Attanasi, M.; Di Pillo, S.; Chiarelli, F. Asthma and Obesity in Children. Biomedicines 2020, 8, 231. https://doi.org/10.3390/biomedicines8070231
Sansone F, Attanasi M, Di Pillo S, Chiarelli F. Asthma and Obesity in Children. Biomedicines. 2020; 8(7):231. https://doi.org/10.3390/biomedicines8070231
Chicago/Turabian StyleSansone, Francesco, Marina Attanasi, Sabrina Di Pillo, and Francesco Chiarelli. 2020. "Asthma and Obesity in Children" Biomedicines 8, no. 7: 231. https://doi.org/10.3390/biomedicines8070231
APA StyleSansone, F., Attanasi, M., Di Pillo, S., & Chiarelli, F. (2020). Asthma and Obesity in Children. Biomedicines, 8(7), 231. https://doi.org/10.3390/biomedicines8070231