
Associations between SNPs in Intestinal Cholesterol Absorption and Endogenous Cholesterol Synthesis Genes with Cholesterol Metabolism




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Blood Sampling and Biochemical Measurements
2.3. DNA Extraction, Genotyping, and Quality Control
2.4. Statistics
3. Results
3.1. Associations between Markers for Cholesterol Absorption and Cholesterol Synthesis, and Serum LDL-C Concentrations
3.2. The Location and Allele Frequencies of the Selected SNPs
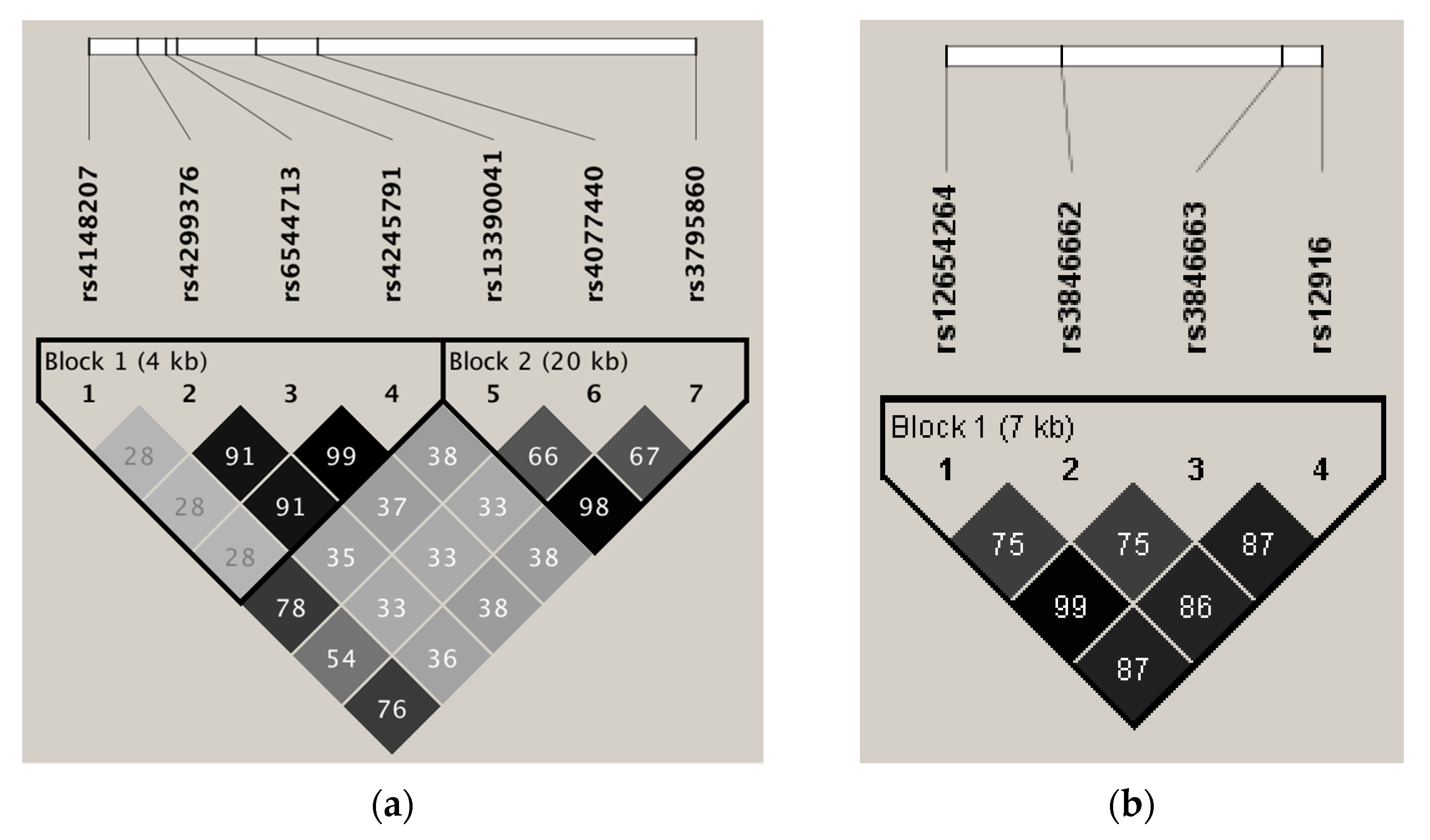
3.3. Linkage Disequilibrium and Tagging for SNPs in Genes Related to Intestinal Cholesterol Absorption
3.4. Linkage Disequilibrium and Tagging for SNPs in Genes Related to Endogenous Cholesterol Synthesis
3.5. Associations between SNPs in ABCG5, ABCG8, and NPC1L1 with TC-Standardized Serum Non-Cholesterol Sterol Levels and Serum LDL-C Concentrations
3.6. Associations between SNPs in CYP51A1, DHCR24, HMGCR, HSD17B7, LBR, and MSMO1 with TC-Standardized Serum Non-Cholesterol Sterol Levels and Serum LDL-C Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mc Auley, M.T.; Wilkinson, D.J.; Jones, J.J.L.; Kirkwood, T.B.L. A whole-body mathematical model of cholesterol metabolism and its age-associated dysregulation. BMC Syst. Biol. 2012, 6, 130. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Betters, J.L.; Yu, L. Niemann-pick C1-like 1 (NPC1L1) protein in intestinal and hepatic cholesterol transport. Annu. Rev. Physiol. 2011, 73, 239–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakulj, L.; Vissers, M.N.; Tanck, M.W.T.; Hutten, B.A.; Stellaard, F.; Kastelein, J.J.P.; Dallinga-Thie, G.M. ABCG5/G8 polymorphisms and markers of cholesterol metabolism: Systematic review and meta-analysis. J. Lipid Res. 2010, 51, 3016–3023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloch, K. The biological synthesis of cholesterol. Science 1965, 150, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Kandutsch, A.A.; Russell, A.E. Preputial gland tumor sterols. I. The occurrence of 24,25-dihydrolanosterol and a comparison with liver and the normal gland. J. Biol. Chem. 1959, 234, 2037–2042. [Google Scholar] [CrossRef]
- Kandutsch, A.A.; Russell, A.E. Preputial gland tumor sterols. 2. The identification of 4 alpha-methyl-Delta 8-cholesten-3 beta-ol. J. Biol. Chem. 1960, 235, 2253–2255. [Google Scholar] [CrossRef]
- Kandutsch, A.A.; Russell, A.E. Preputial gland tumor sterols. 3. A metabolic pathway from lanosterol to cholesterol. J. Biol. Chem. 1960, 235, 2256–2261. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Tilvis, R.S.; Kesaniemi, Y.A. Serum plant sterols and cholesterol precursors reflect cholesterol absorption and synthesis in volunteers of a randomly selected male population. Am. J. Epidemiol. 1990, 131, 20–31. [Google Scholar] [CrossRef] [Green Version]
- Santosa, S.; Varady, K.A.; AbuMweis, S.; Jones, P.J.H. Physiological and therapeutic factors affecting cholesterol metabolism: Does a reciprocal relationship between cholesterol absorption and synthesis really exist? Life Sci. 2007, 80, 505–514. [Google Scholar] [CrossRef]
- Miettinen, T.A.; Gylling, H. Synthesis and absorption markers of cholesterol in serum and lipoproteins during a large dose of statin treatment. Eur. J. Clin. investig. 2003, 33, 976–982. [Google Scholar] [CrossRef]
- Naruse, R.; Hori, K.; Terasawa, T.; Hara, K.; Suetsugu, M.; Takebayashi, K.; Morita, K.; Aso, Y.; Inukai, T. Alterations of plant sterols, lathosterol, oxidative stress and inflammatory markers after the combination therapy of ezetimibe and statin drugs in type 2 diabetic patients. Obes. Res. Clin. Pract. 2015, 9, 67–74. [Google Scholar] [CrossRef]
- Bosner, M.S.; Lange, L.G.; Stenson, W.F.; Ostlund, R.E., Jr. Percent cholesterol absorption in normal women and men quantified with dual stable isotopic tracers and negative ion mass spectrometry. J. Lipid Res. 1999, 40, 302–308. [Google Scholar] [CrossRef]
- Wu, A.H.; Ruan, W.; Todd, J.; Lynch, K.L. Biological variation of beta-sitosterol, campesterol, and lathosterol as cholesterol absorption and synthesis biomarkers. Clin. Chim. Acta 2014, 430, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Alphonse, P.A.; Jones, P.J. Revisiting Human Cholesterol Synthesis and Absorption: The Reciprocity Paradigm and its Key Regulators. Lipids 2016, 51, 519–536. [Google Scholar] [CrossRef] [PubMed]
- Renner, O.; Lütjohann, D.; Richter, D.; Strohmeyer, A.; Schimmel, S.; Müller, O.; Stange, E.F.; Harsch, S. Role of the ABCG8 19H risk allele in cholesterol absorption and gallstone disease. BMC Gastroenterol. 2013, 13, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berge, K.E.; von Bergmann, K.; Lutjohann, D.; Guerra, R.; Grundy, S.M.; Hobbs, H.H.; Cohen, J.C. Heritability of plasma noncholesterol sterols and relationship to DNA sequence polymorphism in ABCG5 and ABCG8. J. Lipid Res. 2002, 43, 486–494. [Google Scholar] [CrossRef]
- Wolff, E.; Vergnes, M.-F.; Defoort, C.; Planells, R.; Portugal, H.; Nicolay, A.; Lairon, D. Cholesterol absorption status and fasting plasma cholesterol are modulated by the microsomal triacylglycerol transfer protein -493 G/T polymorphism and the usual diet in women. Genes. Nutr. 2011, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Teupser, D.; Baber, R.; Ceglarek, U.; Scholz, M.; Illig, T.; Gieger, C.; Holdt, L.M.; Leichtle, A.; Greiser, K.H.; Huster, D.; et al. Genetic regulation of serum phytosterol levels and risk of coronary artery disease. Circ. Cardiovasc. Genet. 2010, 3, 331–339. [Google Scholar] [CrossRef] [Green Version]
- Chung, J.Y.; Cho, S.K.; Oh, E.S.; Lee, D.H.; Lim, L.A.; Jang, S.B.; Lee, Y.J.; Park, K.; Park, M.S. Effect of HMGCR variant alleles on low-density lipoprotein cholesterol-lowering response to atorvastatin in healthy Korean subjects. J. Clin. Pharmacol. 2012, 52, 339–346. [Google Scholar] [CrossRef]
- Chasman, D.I.; Posada, D.; Subrahmanyan, L.; Cook, N.R.; Stanton, V.P., Jr.; Ridker, P.M. Pharmacogenetic study of statin therapy and cholesterol reduction. JAMA 2004, 291, 2821–2827. [Google Scholar] [CrossRef] [Green Version]
- Krauss, R.M.; Mangravite, L.M.; Smith, J.D.; Medina, M.W.; Wang, D.; Guo, X.; Rieder, M.J.; Simon, J.A.; Hulley, S.B.; Waters, D.; et al. Variation in the 3-hydroxyl-3-methylglutaryl coenzyme a reductase gene is associated with racial differences in low-density lipoprotein cholesterol response to simvastatin treatment. Circulation 2008, 117, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Hegele, R.A.; Guy, J.; Ban, M.R.; Wang, J. NPC1L1 haplotype is associated with inter-individual variation in plasma low-density lipoprotein response to ezetimibe. Lipids Health Dis. 2005, 4, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, M.M.; Cyr, A.; Lepine, M.C.; Eck, P.K.; Couture, P.; Lamarche, B.; Jones, P.J. Common Variants in Cholesterol Synthesis- and Transport-Related Genes Associate with Circulating Cholesterol Responses to Intakes of Conventional Dairy Products in Healthy Individuals. J. Nutr. 2016, 146, 1008–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herron, K.L.; McGrane, M.M.; Waters, D.; Lofgren, I.E.; Clark, R.M.; Ordovas, J.M.; Fernandez, M.L. The ABCG5 polymorphism contributes to individual responses to dietary cholesterol and carotenoids in eggs. J. Nutr. 2006, 136, 1161–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Body Mass Index–BMI. Available online: https://www.euro.who.int/en/health-topics/disease-prevention/nutrition/a-healthy-lifestyle/body-mass-index-bmi (accessed on 4 October 2021).
- Kerckhoffs, D.A.; Hornstra, G.; Mensink, R.P. Cholesterol-lowering effect of β-glucan from oat bran in mildly hypercholesterolemic subjects may decrease when β-glucan is incorporated into bread and cookies. Am. J. Clin. Nutr. 2003, 78, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plat, J.; Mensink, R.P. Vegetable oil based versus wood based stanol ester mixtures: Effects on serum lipids and hemostatic factors in non-hypercholesterolemic subjects. Atherosclerosis 2000, 148, 101–112. [Google Scholar] [CrossRef]
- Plat, J.; van Onselen, E.N.M.; van Heugten, M.M.A.; Mensink, R.P. Effects on serum lipids, lipoproteins and fat soluble antioxidant concentrations of consumption frequency of margarines and shortenings enriched with plant stanol esters. Eur. J. Clin. Nutr. 2000, 54, 671–677. [Google Scholar] [CrossRef] [Green Version]
- De Smet, E.; Mensink, R.P.; Lütjohann, D.; Plat, J. Acute effects of plant stanol esters on postprandial metabolism and its relation with changes in serum lipids after chronic intake. Eur. J. Clin. Nutr. 2015, 69, 127–133. [Google Scholar] [CrossRef]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef]
- Mackay, D.S.; Jones, P.J.; Myrie, S.B.; Plat, J.; Lutjohann, D. Methodological considerations for the harmonization of non-cholesterol sterol bio-analysis. J. Chromatogr. Biomed Analyt. Technol. Biomed. Life Sci. 2014, 957, 116–122. [Google Scholar] [CrossRef]
- Axiom™ Precision Medicine Research Array (PMRA) Applied Biosystems™, Catalog Number: 902981. Available online: https://www.thermofisher.com/order/catalog/product/902981#/902981 (accessed on 20 January 2021).
- Purcell, S.; Chang, C. PLINK, Version: 1.90 Beta. Available online: www.cog-genomics.org/plink/1.9/ (accessed on 20 January 2021).
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef] [Green Version]
- Database of Single Nucleotide Polymorphisms (dbSNP). Bethesda (MD): National Center for Biotechnology Information, National Library of Medicine. dbSNP Accession:(dbSNP Build ID: 154). Available online: http://www.ncbi.nlm.nih.gov/SNP/ (accessed on 28 January 2021).
- Burkhardt, R.; Kenny, E.E.; Lowe, J.K.; Birkeland, A.; Josowitz, R.; Noel, M.; Salit, J.; Maller, J.B.; Pe’er, I.; Daly, M.J.; et al. Common SNPs in HMGCR in micronesians and whites associated with LDL-cholesterol levels affect alternative splicing of exon13. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 2078–2084. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, S.; Melander, O.; Anevski, D.; Guiducci, C.; Burtt, N.P.; Roos, C.; Hirschhorn, J.N.; Berglund, G.; Hedblad, B.; Groop, L.; et al. Polymorphisms associated with cholesterol and risk of cardiovascular events. N. Engl. J. Med. 2008, 358, 1240–1249. [Google Scholar] [CrossRef] [Green Version]
- Kathiresan, S.; Melander, O.; Guiducci, C.; Surti, A.; Burtt, N.P.; Rieder, M.J.; Cooper, G.M.; Roos, C.; Voight, B.F.; Havulinna, A.S.; et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat. Genet. 2008, 40, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Wurtz, P.; Wang, Q.; Soininen, P.; Kangas, A.J.; Fatemifar, G.; Tynkkynen, T.; Tiainen, M.; Perola, M.; Tillin, T.; Hughes, A.D.; et al. Metabolomic Profiling of Statin Use and Genetic Inhibition of HMG-CoA Reductase. J. Am. Coll. Cardiol. 2016, 67, 1200–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelini, S.; Rosticci, M.; Massimo, G.; Musti, M.; Ravegnini, G.; Consolini, N.; Sammarini, G.; D’Addato, S.; Rizzoli, E.; Botbayev, D.; et al. Relationship between Lipid Phenotypes, Overweight, Lipid Lowering Drug Response and KIF6 and HMG-CoA Genotypes in a Subset of the Brisighella Heart Study Population. Int. J. Mol. Sci 2018, 19, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Repa, J.J.; Berge, K.E.; Pomajzl, C.; Richardson, J.A.; Hobbs, H.; Mangelsdorf, D.J. Regulation of ATP-binding Cassette Sterol Transporters ABCG5 and ABCG8 by the Liver X Receptors α and β. J. Biol. Chem. 2002, 277, 18793–18800. [Google Scholar] [CrossRef] [Green Version]
- Klett, E.L.; Patel, S.B. Biomedicine. Will the real cholesterol transporter please stand up. Science 2004, 303, 1149–1150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.M.; Yu, L. Opposing Gatekeepers of Apical Sterol Transport: Niemann-Pick C1-Like 1 (NPC1L1) and ATP-Binding Cassette Transporters G5 and G8 (ABCG5/ABCG8). Immunol. Endocr. Metab. Agents. Med. Chem. 2009, 9, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Li-Hawkins, J.; Hammer, R.E.; Berge, K.E.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Overexpression of ABCG5 and ABCG8 promotes biliary cholesterol secretion and reduces fractional absorption of dietary cholesterol. J. Clin. Invest. 2002, 110, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Temel, R.E.; Tang, W.; Ma, Y.; Rudel, L.L.; Willingham, M.C.; Ioannou, Y.A.; Davies, J.P.; Nilsson, L.M.; Yu, L. Hepatic Niemann-Pick C1-like 1 regulates biliary cholesterol concentration and is a target of ezetimibe. J. Clin. Invest. 2007, 117, 1968–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Velde, A.E.; Brufau, G.; Groen, A.K. Transintestinal cholesterol efflux. Curr. Opin. Lipidol. 2010, 21, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Jo, B.S.; Choi, S.S. Introns: The Functional Benefits of Introns in Genomes. Genomics. Inform. 2015, 13, 112–118. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.S.; Cooper, T.A. Splicing in disease: Disruption of the splicing code and the decoding machinery. Nat. Rev. Genet. 2007, 8, 749–761. [Google Scholar] [CrossRef]
- Leduc, V.; Bourque, L.; Poirier, J.; Dufour, R. Role of rs3846662 and HMGCR alternative splicing in statin efficacy and baseline lipid levels in familial hypercholesterolemia. Pharm. Genom. 2016, 26, 1–11. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Gene | Tag SNP | Captured SNP | R2-Value |
|---|---|---|---|
| ABCG8 | rs4245791 | rs6544713 | 0.995 |
| rs4245791 | rs4299376 | 0.919 | |
| rs3795860 | rs13390041 | 0.982 | |
| DHCR24 | rs6676774 | rs7551288 | 0.906 |
| HMGCR | rs12916 | rs12654264 | 0.872 |
| rs12916 | rs3846662 | 0.862 | |
| rs12916 | rs3846663 | 0.879 |
| Gene | SNP | Genotype | N | Campesterol 102 × µmol/mmol TC | Sitosterol 102 × µmol/mmol TC | Lathosterol 102 × µmol/mmol TC | N | LDL-C mmol/L | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean (95% CI) | p-Value | Mean (95% CI) | p-Value | Mean (95% CI) | p-Value | Mean (95% CI) | p-Value | |||||
| ABCG5 | rs4245786 | AA | 266 | 252 (236–267) | 0.074 | 152 (142–162) | 0.041 $ | 120 (112–129) | 0.959 | 266 | 3.44 (3.30–3.57) | 0.306 |
| AG | 160 | 230 (212–249) | 136 (124–148) | 120 (110–131) | 161 | 3.34 (3.18–3.50) | ||||||
| GG | 29 | 259 (222–296) | 154 (130–178) | 123 (103–144) | 29 | 3.23 (2.90–3.55) | ||||||
| rs7599296 | AA | 15 | 261 (210–312) | 0.228 | 164 (131–197) | 0.173 | 109 (81–137) | 0.653 | 15 | 3.40 (2.95–3.85) | 0.980 | |
| AG | 141 | 255 (236–274) | 152 (140–165) | 119 (108–130) | 141 | 3.38 (3.21–3.54) | ||||||
| GG | 299 | 239 (224–254) | 143 (133–152) | 122 (113–130) | 300 | 3.39 (3.26–3.52) | ||||||
| rs4148184 | TT | 74 | 232 (207–256) | 0.297 | 142 (126–158) | 0.803 | 117 (103–130) | 0.217 | 74 | 3.30 (3.08–3.51) | 0.561 | |
| TC | 219 | 251 (235–268) | 148 (137–159) | 117 (108–126) | 219 | 3.42 (3.28–2.57) | ||||||
| CC | 161 | 242 (223–260) | 146 (134–158) | 126 (116–137) | 162 | 3.39 (3.23–3.55) | ||||||
| rs13396273 | TT | 53 | 236 (207–264) | 0.431 | 144 (126–163) | 0.819 | 116 (101–132) | 0.526 | 53 | 3.36 (3.11–3.60) | 0.922 | |
| TC | 214 | 251 (234–267) | 148 (138–159) | 119 (109–128) | 214 | 3.40 (3.26–3.55) | ||||||
| CC | 188 | 240 (222–257) | 145 (133–156) | 124 (114–134) | 189 | 3.38 (3.22–3.53) | ||||||
| ABCG8 | rs4148207 | TT | 156 | 249 (231–268) | 0.757 | 151 (139–163) | 0.364 | 121 (111–131) | 0.713 | 157 | 3.34 (3.18–3.50) | 0.530 |
| TC | 227 | 243 (226–259) | 145 (123–155) | 121 (112–130) | 227 | 3.43 (3.29–3.58) | ||||||
| CC | 72 | 241 (216–266) | 139 (123–155) | 116 (102–129) | 72 | 3.35 (3.13–3.57) | ||||||
| rs3795860 + | TT | 128 | 253 (234–273) | 154 (141–167) | 120 (109–131) | 129 | 3.32 (3.15–3.50) | |||||
| TC | 233 | 244 (228–260) | 0.342 | 146 (135–156) | 0.174 | 123 (114–131) | 0.515 | 233 | 3.46 (3.32–3.60) | 0.175 | ||
| CC | 94 | 234 (211–257) | 138 (123–152) | 115 (102–127) | 94 | 3.29 (3.09–3.49) | ||||||
| rs4077440 | TT | 92 | 256 (233–279) | 0.129 | 154 (140–169) | 0.125 | 120 (107–132) | 0.378 | 92 | 3.38 (3.18–3.58) | 0.252 | |
| TC | 217 | 249 (232–266) | 149 (138–159) | 124 (115–133) | 218 | 3.45 (3.31–1.60) | ||||||
| CC | 145 | 232 (213–251) | 138 (126–150) | 116 (105–126) | 145 | 3.30 (3.13–3.46) | ||||||
| AX_82902928 | -- | 197 | 248 (231–265) | 0.752 | 151 (140–161) | 0.334 | 120 (111–130) | 0.955 | 197 | 3.40 (3.25–3.55) | 0.145 | |
| -AC | 192 | 240 (223–258) | 141 (130–165) | 120 (110–130) | 193 | 3.43 (3.28–3.58) | ||||||
| ACAC | 66 | 246 (219–272) | 147 (130–165) | 122 (108–137) | 66 | 3.19 (2.60–3.42) | ||||||
| rs4245791 + | TT | 206 | 221 (205–237) A | 130 (120–141) A | 123 (114–132) | 206 | 3.32 (3.17–3.47) | |||||
| TC | 215 | 256 (239–272) B | <0.001 $ | 153 (143–164) B | <0.001 $ | 119 (109–128) | 0.642 | 216 | 3.46 (3.31–3.61) | 0.239 | ||
| CC | 34 | 315 (282–349) C | 180 (176–219) C | 117 (97–136) | 34 | 3.34 (3.04–3.65) | ||||||
| NPC1L1 | rs217429 | AA | 259 | 239 (223–254) | 0.190 | 142 (132–152) | 0.134 | 119 (110–128) A | 0.017 # | 259 | 3.37 (3.23–3.50) | 0.825 |
| AC | 169 | 256 (238–275) | 154 (142–166) | 117 (107–127) A | 170 | 3.42 (3.26–3.58) | ||||||
| CC | 27 | 238 (200–276) | 146 (121–170) | 149 (128–170) B | 27 | 3.39 (3.06–3.73) | ||||||
| rs217416 | TT | 239 | 240 (223–256) | 0.208 | 143 (132–153) | 0.236 | 119 (110–127) A | 0.020 # | 239 | 3.40 (3.26–3.54) | 0.922 | |
| TC | 189 | 254 (237–272) | 153 (141–164) | 118 (108–128) A | 190 | 3.38 (3.23–3.54) | ||||||
| CC | 25 | 228 (188–267) | 140 (114–165) | 149 (128–171) B | 25 | 3.33 (2.98–3.67) | ||||||
| rs11763759 | TT | 208 | 244 (227–261) | 0.961 | 145 (134–156) | 0.938 | 120 (111–130) | 0.953 | 209 | 3.42 (3.27–3.56) | 0.084 | |
| TC | 202 | 246 (229–263) | 147 (136–158) | 120 (111–129) | 202 | 3.31 (3.16–3.46) | ||||||
| CC | 43 | 242 (211–273) | 149 (128–169) | 123 (106–140) | 43 | 3.62 (3.35–3.89) | ||||||
| rs2072183 | CC | 18 | 260 (213–307) | 0.314 | 154 (123–184) | 0.361 | 121 (95–147) | 0.862 | 18 | 3.33 (2.91–3.75) | 0.930 | |
| CG | 173 | 254 (235–272) | 152 (140–164) | 122 (112–133) | 174 | 3.40 (3.24–3.57) | ||||||
| GG | 263 | 240 (225–255) | 143 (134–153) | 119 (111–128) | 263 | 3.38 (3.25–3.52) | ||||||
| Gene | SNP | Genotype | N | Campesterol 102 × µmol/mmol TC | Sitosterol 102 × µmol/mmol TC | Lathosterol 102 × µmol/mmol TC | N | LDL-C mmol/L | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean (95% CI) | p-Value | Mean (95% CI) | p-Value | Mean (95% CI) | p-Value | Mean (95% CI) | p-Value | |||||
| CYP51A1 | rs35968894 | AA | 161 | 240 (222–258) | 0.239 | 142 (131–154) | 0.334 | 115 (104–124) | 0.066 | 161 | 3.40 (3.24–3.56) | 0.976 |
| AG | 223 | 241 (224–258) | 146 (135–157) | 127 (118–136) | 224 | 3.38 (3.23–3.53) | ||||||
| GG | 71 | 262 (238–287) | 156 (140–172) | 117 (103–131) | 71 | 3.39 (3.17–3.60) | ||||||
| DHCR24 | rs6676774 + | AA | 75 | 231 (207–256) | 0.436 | 144 (128–160) | 0.887 | 120 (106–134) | 0.535 | 75 | 3.42 (3.20–3.63) | |
| AG | 208 | 246 (230–263) | 146 (135–157) | 123 (114–132) | 208 | 3.30 (3.16–3.45) | 0.122 | |||||
| GG | 172 | 249 (230–267) | 148 (136–160) | 117 (107–127) | 173 | 3.48 (3.33–3.64) | ||||||
| rs718265 | AA | 43 | 231 (200–263) | 0.292 | 143 (123–164) | 0.794 | 117 (98–134) | 0.570 | 43 | 3.35 (3.07–3.62) | 0.460 | |
| AG | 190 | 252 (235–269) | 149 (138–160) | 123 (114–133) | 190 | 3.34 (3.19–3.49) | ||||||
| GG | 222 | 240 (223–257) | 145 (134–156) | 118 (109–127) | 223 | 3.44 (3.29–3.59) | ||||||
| HMGCR | rs12916 + | TT | 151 | 240 (221–260) | 0.373 | 145 (133–158) | 0.541 | 122 (112–133) | 0.838 | 152 | 3.22 (3.05–3.39) A | 0.011 @ |
| TC | 231 | 242 (226–259) | 145 (134–155) | 119 (110–128) | 231 | 3.49 (3.35–3.63) B | ||||||
| CC | 73 | 259 (234–284) | 154 (138–170) | 122 (108–135) | 73 | 3.35 (3.13–3.56) | ||||||
| HSD17B7 | rs77482353 | AA | 156 | 241 (222–259) | 0.676 | 142 (130–154) | 0.516 | 121 (111–131) | 0.889 | 156 | 3.40 (3.24–3.56) | 0.070 |
| AG | 227 | 250 (233–266) | 150 (139–160) | 120 (111–130) | 228 | 3.32 (3.18–3.47) | ||||||
| GG | 68 | 246 (220–272) | 150 (133–167) | 117 (103–132) | 68 | 3.60 (3.72–3.83) | ||||||
| LBR | rs6678087 | TT | 141 | 247 (228–267) | 0.367 | 147 (134–160) | 0.988 | 120 (109–131) | 0.997 | 141 | 3.41 (3.24–3.58) | 0.970 |
| TC | 223 | 248 (232–265) | 147 (136–157) | 121 (112–130) | 223 | 3.39 (2.25–3.53) | ||||||
| CC | 90 | 232 (209–254) | 146 (131–161) | 120 (108–133) | 91 | 3.39 (3.19–3.59) | ||||||
| rs12141732 | TT | 226 | 241 (224–258) | 0.706 | 144 (133–155) | 0.453 | 121 (111–130) | 0.799 | 227 | 3.50 (3.35–3.65) A | 0.027 @ | |
| TC | 194 | 248 (232–265) | 147 (136–158) | 121 (112–130) | 194 | 3.28 (3.13–3.43) B | ||||||
| CC | 34 | 251 (216–286) | 159 (136–182) | 114 (95–134) | 34 | 3.50 (3.20–3.81) | ||||||
| MSMO1 | rs17046216 | AA | 53 | 237 (209–266) | 0.112 | 147 (128–165) | 0.347 | 113 (97–128) | 0.542 | 53 | 3.63 (3.38–3.88) | 0.101 |
| AG | 205 | 236 (219–253) | 142 (131–153) | 121 (112–131) | 206 | 3.35 (3.21–3.50) | ||||||
| GG | 197 | 256 (239–273) | 151 (140–162) | 122 (112–131) | 197 | 3.36 (3.21–3.51) | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schroor, M.M.; Mokhtar, F.B.A.; Plat, J.; Mensink, R.P. Associations between SNPs in Intestinal Cholesterol Absorption and Endogenous Cholesterol Synthesis Genes with Cholesterol Metabolism. Biomedicines 2021, 9, 1475. https://doi.org/10.3390/biomedicines9101475
Schroor MM, Mokhtar FBA, Plat J, Mensink RP. Associations between SNPs in Intestinal Cholesterol Absorption and Endogenous Cholesterol Synthesis Genes with Cholesterol Metabolism. Biomedicines. 2021; 9(10):1475. https://doi.org/10.3390/biomedicines9101475
Chicago/Turabian StyleSchroor, Maite M., Fatma B. A. Mokhtar, Jogchum Plat, and Ronald P. Mensink. 2021. "Associations between SNPs in Intestinal Cholesterol Absorption and Endogenous Cholesterol Synthesis Genes with Cholesterol Metabolism" Biomedicines 9, no. 10: 1475. https://doi.org/10.3390/biomedicines9101475
APA StyleSchroor, M. M., Mokhtar, F. B. A., Plat, J., & Mensink, R. P. (2021). Associations between SNPs in Intestinal Cholesterol Absorption and Endogenous Cholesterol Synthesis Genes with Cholesterol Metabolism. Biomedicines, 9(10), 1475. https://doi.org/10.3390/biomedicines9101475