
Long-Term Treatment of Azathioprine in Rats Induces Vessel Mineralization

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Blood Parameters
2.3. Histological Staining
2.4. Quantification of the Tissue Calcium Content
2.5. Ex Vivo Stimulation of Aortic Tissue
2.6. Gene Expression
2.7. Statistical Analysis
3. Results
3.1. In Vivo Model
3.2. Effect of Azathioprine on Vessel Mineralization after 24 Weeks of Treatment
3.3. Azathioprine Treatment Induces Oxidative Stress and Reduces Antioxidative Capacity in Aortic Tissue
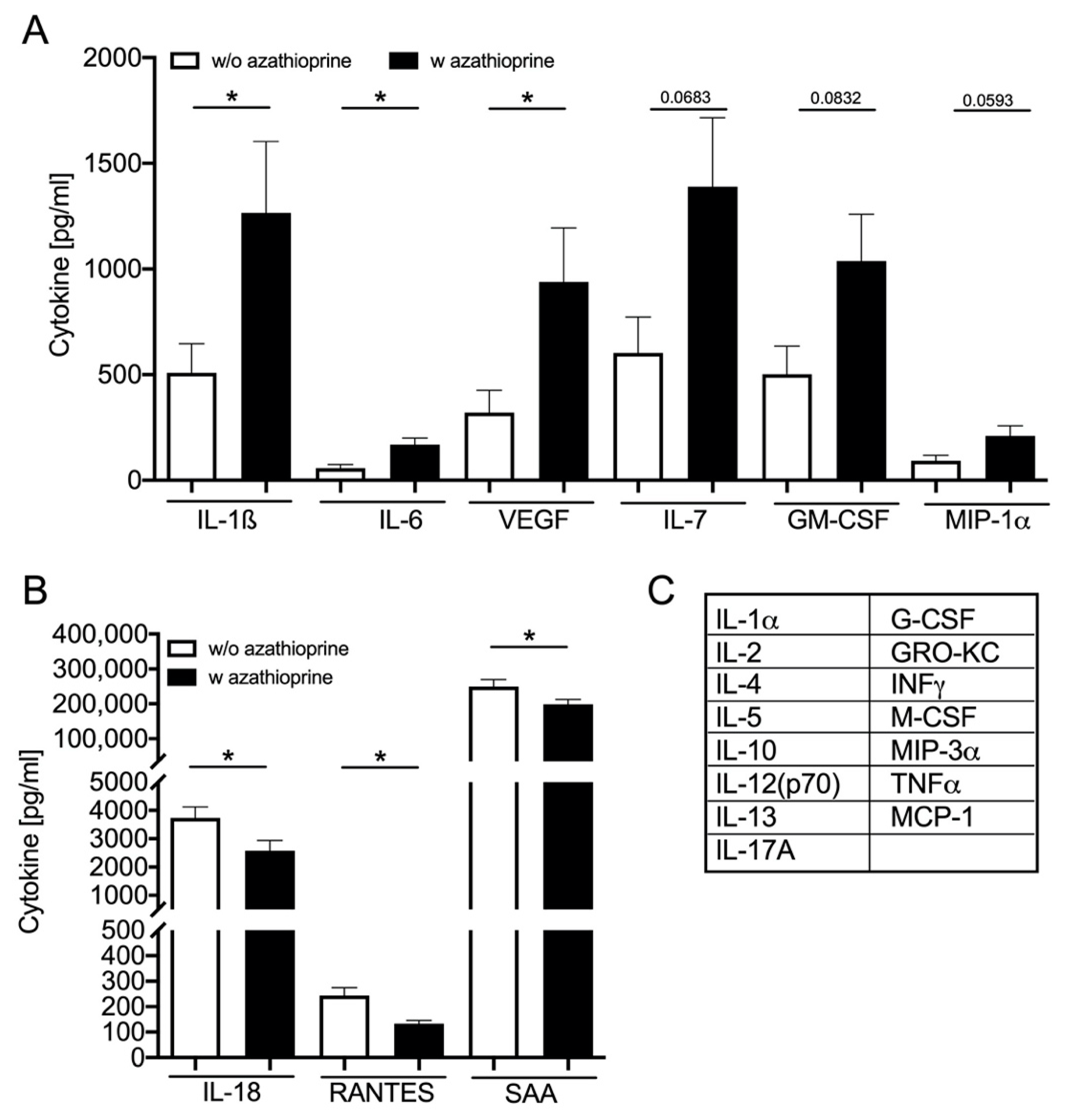
3.4. Azathioprine Treatment Induces SASP: Cytokine Plasma Level
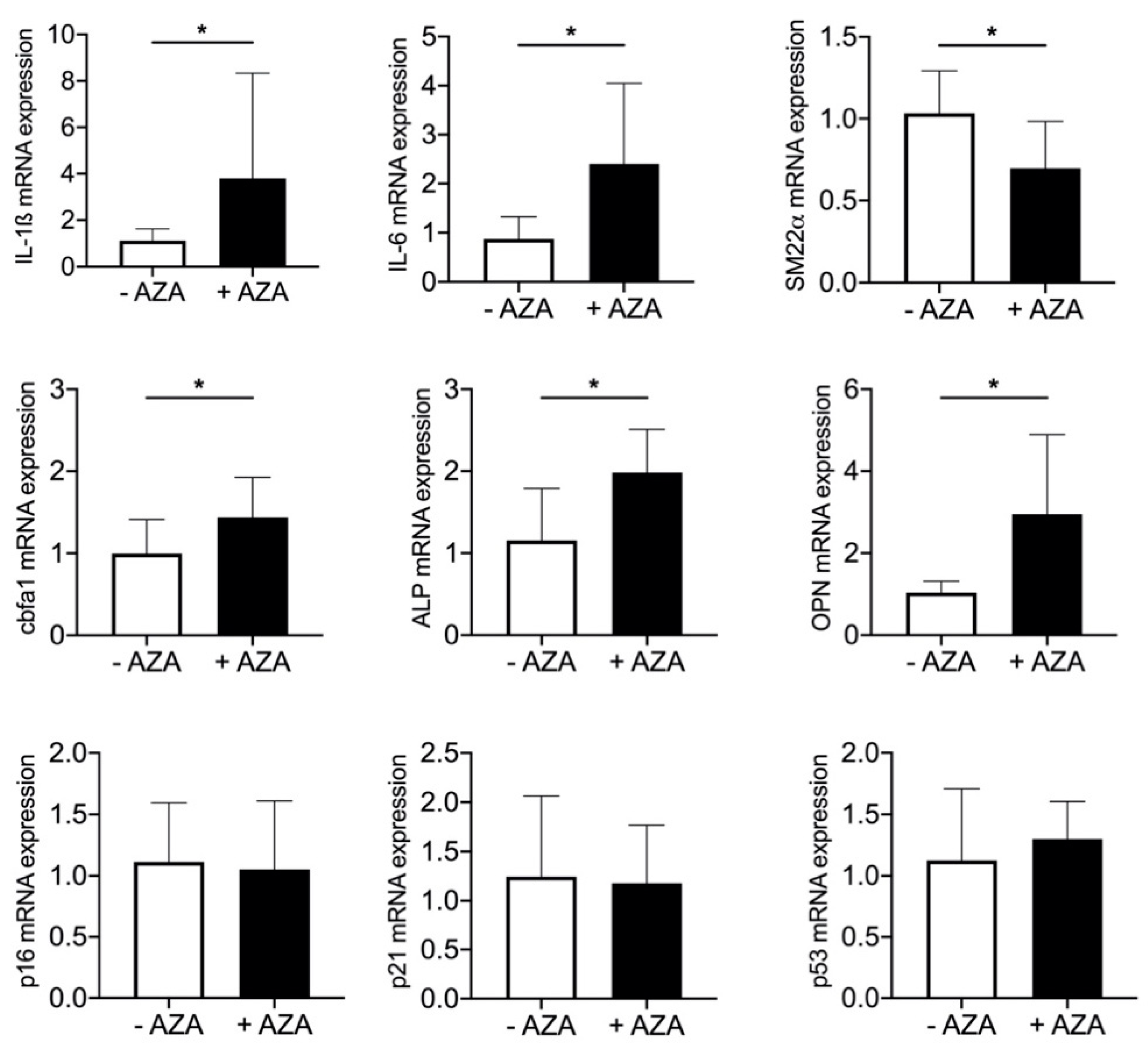
3.5. Azathioprine Treatment Induces SASP and Mineralization in Aortic Tissue
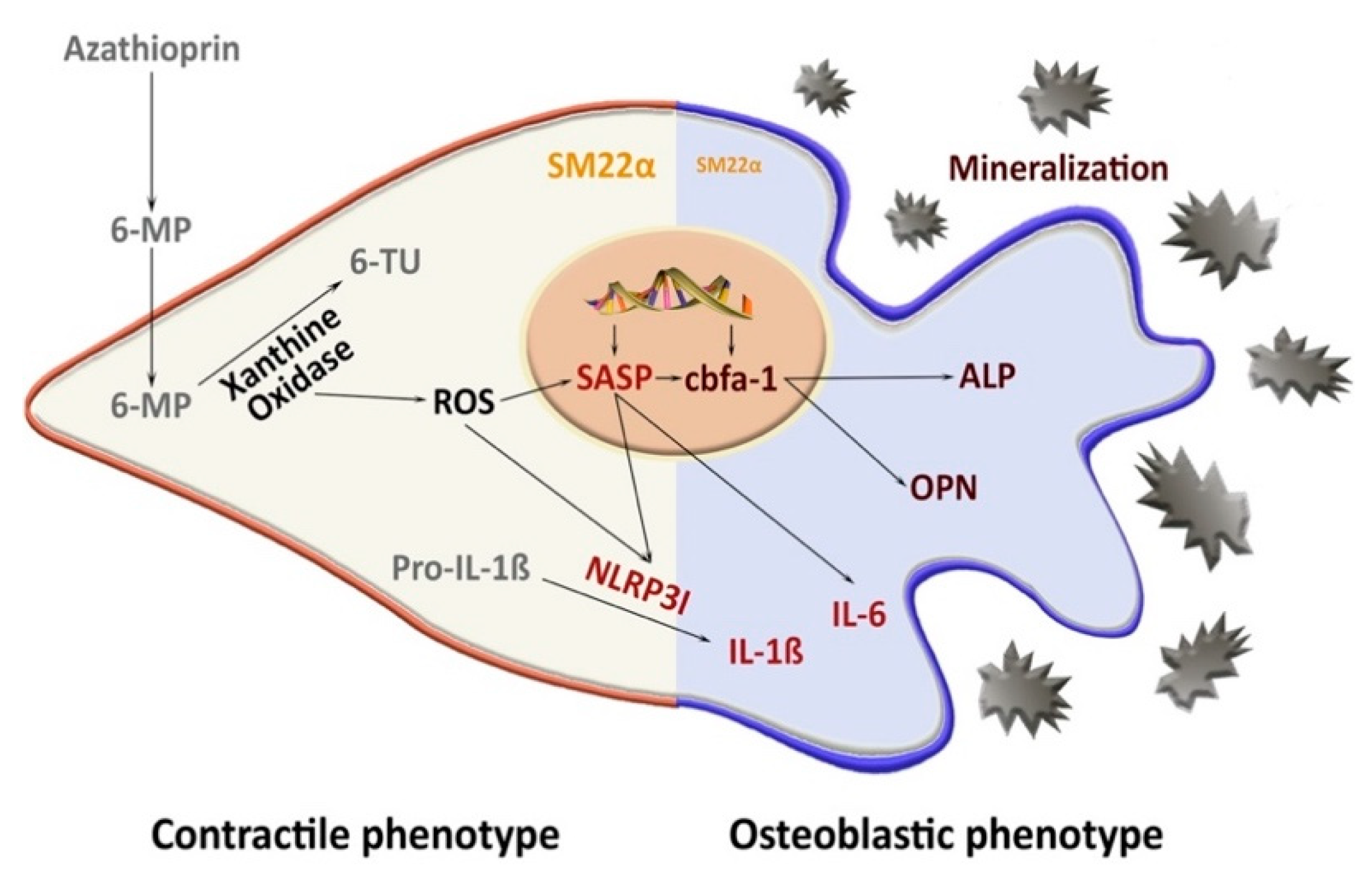
3.6. SASP Induction Is NLRP3-Dependent
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Tölle, M.; Reshetnik, A.; Schuchardt, M.; Höhne, M.; van der Giet, M. Arteriosclerosis and vascular calcification: Causes, clinical assessment and therapy. Eur. J. Clin. Investig. 2015, 45, 976–985. [Google Scholar] [CrossRef]
- Bottomley, M.J.; Harden, P.N. Update on the long-term complications of renal transplantation. Br. Med. Bull. 2013, 106, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Maréchal, C.; Coche, E.; Goffin, E.; Dragean, A.; Schlieper, G.; Nguyen, P.; Floege, J.; Kanaan, N.; Devuyst, O.; Jadoul, M. Progression of Coronary Artery Calcification and Thoracic Aorta Calcification in Kidney Transplant Recipients. Am. J. Kidney Dis. 2012, 59, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Hénaut, L.; Mary, A.; Chillon, J.-M.; Kamel, S.; Massy, Z.A. The Impact of Uremic Toxins on Vascular Smooth Muscle Cell Function. Toxins 2018, 10, 218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liefeldt, L.; Budde, K. Risk factors for cardiovascular disease in renal transplant recipients and strategies to minimize risk. Transpl. Int. 2010, 23, 1191–1204. [Google Scholar] [CrossRef] [PubMed]
- Halloran, P.F. Immunosuppressive Drugs for Kidney Transplantation. New Engl. J. Med. 2004, 351, 2715–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gearry, R.B.; Barclay, M.L. Azathioprine and 6-mercaptopurine pharmacogenetics and metabolite monitoring in inflammatory bowel disease. J. Gastroenterol. Hepatol. 2005, 20, 1149–1157. [Google Scholar] [CrossRef]
- Solomon, D.H.; Avorn, J.; Katz, J.N.; Weinblatt, M.E.; Setoguchi, S.; Levin, R.; Schneeweiss, S. Immunosuppressive medications and hospitalization for cardiovascular events in patients with rheumatoid arthritis. Arthritis Rheum. 2006, 54, 3790–3798. [Google Scholar] [CrossRef]
- Haque, S.; Gordon, C.; Isenberg, D.; Rahman, A.; Lanyon, P.; Bell, A.; Emery, P.; McHugh, N.; Teh, L.S.; Scott, D.G.; et al. Risk Factors for Clinical Coronary Heart Disease in Systemic Lupus Erythematosus: The Lupus and Atherosclerosis Evaluation of Risk (LASER) Study. J. Rheumatol. 2009, 37, 322–329. [Google Scholar] [CrossRef] [Green Version]
- Prüfer, J.; Schuchardt, M.; Tolle, M.; Prüfer, N.; Höhne, M.; Zidek, W.; van der Giet, M. Harmful Effects of the Azathioprine Metabolite 6-Mercaptopurine in Vascular Cells: Induction of Mineralization. PLoS ONE 2014, 9, e101709. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, I.-K.; Jeon, J.-H. Vascular Calcification—New Insights into Its Mechanism. Int. J. Mol. Sci. 2020, 21, 2685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voelkl, J.; Lang, F.; Eckardt, K.-U.; Amann, K.; Kuro, O.M.; Pasch, A.; Pieske, B.; Alesutan, I. Signaling pathways involved in vascular smooth muscle cell calcification during hyperphosphatemia. Cell. Mol. Life Sci. 2019, 76, 2077–2091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchis, P.; Ho, C.Y.; Liu, Y.; Beltran, L.E.; Ahmad, S.; Jacob, A.P.; Furmanik, M.; Laycock, J.; Long, D.A.; Shroff, R.; et al. Arterial “inflammaging” drives vascular calcification in children on dialysis. Kidney Int. 2019, 95, 958–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benz, K.; Varga, I.; Neureiter, D.; Campean, V.; Daniel, C.; Heim, C.; Reimann, A.; Weyand, M.; Hilgers, K.F.; Amann, K. Vascular inflammation and media calcification are already present in early stages of chronic kidney disease. Cardiovasc. Pathol. 2017, 27, 57–67. [Google Scholar] [CrossRef]
- Wen, C.; Yang, X.; Yan, Z.; Zhao, M.; Yue, X.; Cheng, X.; Zheng, Z.; Guan, K.; Dou, J.; Xu, T.; et al. Nalp3 inflammasome is activated and required for vascular smooth muscle cell calcification. Int. J. Cardiol. 2013, 168, 2242–2247. [Google Scholar] [CrossRef] [PubMed]
- Kurozumi, A.; Nakano, K.; Yamagata, K.; Okada, Y.; Nakayamada, S.; Tanaka, Y. IL-6 and sIL-6R induces STAT3-dependent differentiation of human VSMCs into osteoblast-like cells through JMJD2B-mediated histone demethylation of RUNX2. Bone 2019, 124, 53–61. [Google Scholar] [CrossRef]
- Alvarez-Leite, J.I.; dos Santos, L.C.; Costa, A.V.; Lopes, L.G.; Leonel, A.J.; Aguilar, E.C.; Noviello, M.D.L.M.; Ferrari, M.D.L. Combination of Azathioprine and Aminosalicylate Treatment Prevent Risk of Cardiovascular Disease in Women with Ulcerative Colitis by Reducing Inflammation. Med. Sci. Monit. 2015, 21, 2305–2315. [Google Scholar] [CrossRef] [Green Version]
- Schuchardt, M.; Tölle, M.; Prüfer, J.; Prüfer, N.; Huang, T.; Jankowski, V.; Jankowski, J.; Zidek, W.; van der Giet, M. Uridine adenosine tetraphosphate activation of the purinergic receptor P2Y enhances in vitro vascular calcification. Kidney Int. 2012, 81, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Bang, E.; Arulkumar, R.; Ha, S.; Chung, K.W.; Park, M.H.; Choi, Y.J.; Yu, B.P.; Chung, H.Y. Senoinflammation: A major mediator underlying age-related metabolic dysregulation. Exp. Gerontol. 2020, 134, 110891. [Google Scholar] [CrossRef]
- Polifka, J.E.; Friedman, J. Teratogen update: Azathioprine and 6-mercaptopurine. Teratology 2002, 65, 240–261. [Google Scholar] [CrossRef] [PubMed]
- Pols, T.W.; Bonta, P.I.; Pires, N.M.; Otermin, I.; Vos, M.; de Vries, M.R.; van Eijk, M.; Roelofsen, J.; Havekes, L.M.; Quax, P.H.; et al. 6-Mercaptopurine Inhibits Atherosclerosis in Apolipoprotein E*3-Leiden Transgenic Mice Through Atheroprotective Actions on Monocytes and Macrophages. Arter. Thromb. Vasc. Biol. 2010, 30, 1591–1597. [Google Scholar] [CrossRef] [Green Version]
- Ligumsky, M.; Badaan, S.; Lewis, H.; Meirow, D. Effects of 6-mercaptopurine treatment on sperm production and reproductive performance: A study in male mice. Scand. J. Gastroenterol. 2005, 40, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Kles, V.; Hyrien, O.; Poul, J.-M.; Sanders, P. Application of pharmacokinetic/pharmacodynamic and stochastic modelling to 6-mercaptopurine micronucleus induction in mouse bone marrow erythrocytes. J. Appl. Toxicol. 2003, 23, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Platzek, T.; Schwabe, R. Combined prenatal toxicity of 6-mercaptopurine riboside and hydroxyurea in mice. Teratog. Carcinog. Mutagen. 1999, 19, 223–232. [Google Scholar] [CrossRef]
- Chang, C.-Z.; Lin, C.-L.; Kassel, N.F.; Kwan, A.-L.; Howng, S.-L. 6-Mercaptopurine attenuates adhesive molecules in experimental vasospasm. Acta Neurochir. 2010, 152, 861–867. [Google Scholar] [CrossRef]
- Furukawa, S.; Hayashi, S.; Usuda, K.; Abe, M.; Ogawa, I. The relationship between fetal growth restriction and small placenta in 6-mercaptopurine exposed rat. Exp. Toxicol. Pathol. 2011, 63, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Kanemitsu, H.; Yamauchi, H.; Komatsu, M.; Yamamoto, S.; Okazaki, S.; Uchida, K.; Nakayama, H. 6-Mercaptopurine (6-MP) induces cell cycle arrest and apoptosis of neural progenitor cells in the developing fetal rat brain. Neurotoxicol. Teratol. 2009, 31, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Pires, N.M.; Pols, T.W.; de Vries, M.R.; van Tiel, C.M.; Bonta, P.I.; Vos, M.; Arkenbout, E.K.; Pannekoek, H.; Jukema, J.W.; Quax, P.H.; et al. Activation of Nuclear Receptor Nur77 by 6-Mercaptopurine Protects Against Neointima Formation. Circulation 2007, 115, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cegiela, U.; Kaczmarczyk-Sedlak, I.; Pytlik, M.; Folwarczna, J.; Nowinska, B.; Fronczek-Sokol, J. Alendronate prevents development of the skeletal changes induced by azathioprine in rats. Acta Pol. Pharm. 2013, 70, 309–315. [Google Scholar] [PubMed]
- Herrmann, J.; Babic, M.; Tölle, M.; van der Giet, M.; Schuchardt, M. Research Models for Studying Vascular Calcification. Int. J. Mol. Sci. 2020, 21, 2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiss, A.; du Chesne, A.; Denecke, B.; Grötzinger, J.; Yamamoto, K.; Renné, T.; Jahnen-Dechent, W. Structural Basis of Calcification Inhibition by α2-HS Glycoprotein/Fetuin-A. J. Biol. Chem. 2003, 278, 13333–13341. [Google Scholar] [CrossRef] [Green Version]
- Vassalle, C.; Iervasi, G. New insights for matrix Gla protein, vascular calcification and cardiovascular risk and outcome. Atherosclerosis 2014, 235, 236–238. [Google Scholar] [CrossRef]
- Byon, C.H.; Heath, J.M.; Chen, Y. Redox signaling in cardiovascular pathophysiology: A focus on hydrogen peroxide and vascular smooth muscle cells. Redox Biol. 2016, 9, 244–253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Alexander, H.D.; Ross, O.A. Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. Front. Immunol. 2018, 9, 586. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, Y.; Zhang, M.; Guo, L.; Wang, J.; Zeng, F.; Xu, D.; Yin, Z.; Xu, Y.; Wang, D.; et al. Interleukin-1β-Induced Senescence Promotes Osteoblastic Transition of Vascular Smooth Muscle Cells. Kidney Blood Press. Res. 2020, 45, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Burton, D.G.; Giles, P.J.; Sheerin, A.N.; Smith, S.K.; Lawton, J.J.; Ostler, E.L.; Rhys-Williams, W.; Kipling, D.; Faragher, R.G. Microarray analysis of senescent vascular smooth muscle cells: A link to atherosclerosis and vascular calcification. Exp. Gerontol. 2009, 44, 659–665. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Zeng, F.; Han, L.; Wang, J.; Yin, Z.; Lv, L.; Guo, L.; Wang, D.; Xu, Y.; Zhou, H. The synergistic action of phosphate and interleukin-6 enhances senescence-associated calcification in vascular smooth muscle cells depending on p53. Mech. Ageing Dev. 2019, 182, 111124. [Google Scholar] [CrossRef]
- Acosta, J.C.; Banito, A.; Wuestefeld, T.; Georgilis, A.; Janich, P.; Morton, J.P.; Athineos, D.; Kang, T.-W.; Lasitschka, F.; Andrulis, M.; et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat. Cell Biol. 2013, 15, 978–990. [Google Scholar] [CrossRef]
- Sprague, A.H.; Khalil, R.A. Inflammatory cytokines in vascular dysfunction and vascular disease. Biochem. Pharmacol. 2009, 78, 539–552. [Google Scholar] [CrossRef] [Green Version]
- Schuchardt, M.; Prüfer, N.; Tu, Y.; Herrmann, J.; Hu, X.-P.; Chebli, S.; Dahlke, K.; Zidek, W.; van der Giet, M.; Tölle, M. Dysfunctional high-density lipoprotein activates toll-like receptors via serum amyloid A in vascular smooth muscle cells. Sci. Rep. 2019, 9, 3421. [Google Scholar] [CrossRef] [Green Version]
- Hénaut, L.; Massy, Z.A. New insights into the key role of interleukin 6 in vascular calcification of chronic kidney disease. Nephrol. Dial. Transplant. 2018, 33, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Chang, Q.; Xin, M.; Wang, Q.; Li, H.; Qian, J. Endogenous bone morphogenetic protein 2 plays a role in vascular smooth muscle cell calcification induced by interleukin 6 in vitro. Int. J. Immunopathol. Pharmacol. 2017, 30, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Watson, A.D.; Ji, S.; Boströmm, K.I. Heat Shock Protein 70 Enhances Vascular Bone Morphogenetic Protein-4 Signaling by Binding Matrix Gla Protein. Circ. Res. 2009, 105, 575–584. [Google Scholar] [CrossRef]
- Callegari, A.; Coons, M.L.; Ricks, J.L.; Rosenfeld, M.E.; Scatena, M. Increased Calcification in Osteoprotegerin-Deficient Smooth Muscle Cells: Dependence on Receptor Activator of NF-κB Ligand and Interleukin 6. J. Vasc. Res. 2014, 51, 118–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyson, K.L.; Reynolds, J.L.; McNair, R.; Zhang, Q.; Weissberg, P.L.; Shanahan, C.M. Osteo/Chondrocytic Transcription Factors and Their Target Genes Exhibit Distinct Patterns of Expression in Human Arterial Calcification. Arter. Thromb. Vasc. Biol. 2003, 23, 489–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.-E.; Chen, T.; Leaf, E.M.; Speer, M.Y.; Giachelli, C.M. Runx2 Expression in Smooth Muscle Cells Is Required for Arterial Medial Calcification in Mice. Am. J. Pathol. 2015, 185, 1958–1969. [Google Scholar] [CrossRef] [PubMed]
- Scatena, M.; Liaw, L.; Giachelli, C.M. Osteopontin: A multifunctional molecule regulating chronic inflammation and vascular disease. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2302–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giachelli, C.M.; Speer, M.Y.; Li, X.; Rajachar, R.M.; Yang, H. Regulation of Vascular Calcification. Circ. Res. 2005, 96, 717–722. [Google Scholar] [CrossRef] [Green Version]
- Wada, T.; McKee, M.D.; Steitz, S.; Giachelli, C.M. Calcification of vascular smooth muscle cell cultures: Inhibition by osteo-pontin. Circ. Res. 1999, 84, 166–178. [Google Scholar] [CrossRef]
- Speer, M.Y.; McKee, M.D.; Guldberg, R.E.; Liaw, L.; Yang, H.-Y.; Tung, E.; Karsenty, G.; Giachelli, C.M. Inactivation of the Osteopontin Gene Enhances Vascular Calcification of Matrix Gla Protein–deficient Mice. J. Exp. Med. 2002, 196, 1047–1055. [Google Scholar] [CrossRef]
- Paloian, N.J.; Leaf, E.M.; Giachelli, C.M. Osteopontin protects against high phosphate-induced nephrocalcinosis and vascular calcification. Kidney Int. 2016, 89, 1027–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takemura, A.; Iijima, K.; Ota, H.; Son, B.-K.; Ito, Y.; Ogawa, S.; Eto, M.; Akishita, M.; Ouchi, Y. Sirtuin 1 Retards Hyperphosphatemia-Induced Calcification of Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2054–2062. [Google Scholar] [CrossRef] [Green Version]
- Gardner, S.E.; Humphry, M.; Bennett, M.R.; Clarke, M.C. Senescent Vascular Smooth Muscle Cells Drive Inflammation Through an Interleukin-1α–Dependent Senescence-Associated Secretory Phenotype. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1963–1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, Y.; Yang, P.; Liu, X.; Lu, L.; Chen, Y.; Zhong, X.; Li, Z.; Liu, H.; Ou, C.; et al. Trimethylamine-N-Oxide Promotes Vascular Calcification Through Activation of NLRP3 (Nucleotide-Binding Domain, Leucine-Rich-Containing Family, Pyrin Domain-Containing-3) Inflammasome and NF-κB (Nuclear Factor κB) Signals. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen & Co Ltd.: London, UK, 1959. [Google Scholar]
- Otto, G.M.; Franklin, C.L.; Clifford, C.B. Biology and Diseases of Rats. Lab. Anim. Med. 2015, 151–207. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Rat N | Body Weight [g] Initial vs. Final | Heart Weight [mg/g] | Kidney Weight [mg/g] | ALP [U/L] | IP [mmol/L] | Ca [mmol/L] | Ca x IP | PTH [pg/mL] |
|---|---|---|---|---|---|---|---|---|---|
| Control | 13 | 332.1 ± 6.2 504.2 ± 10.5 # | 2.54 ± 0.06 | 5.09 ± 0.10 | 148.50 ± 18.49 | 2.03 ± 0.12 | 2.41 ± 0.07 | 4.9 ± 0.34 | 888.0 ± 155.4 |
| AZA | 16 | 344.9 ± 5.0 511.4 ± 11.9 # | 2.60 ± 0.09 | 5.21 ± 0.13 | 204.30 1 ± 23.54 * | 2.10 ± 0.07 | 2.63 ± 0.08 * | 5.5 ± 0.25 | 1.347.0 ± 387.7 |
| Group | Rat Number | Fetuin-A [ng/mL] | MGP [ng/mL] |
|---|---|---|---|
| Control | 13 | 89.22 ± 0.28 | 92.25 ± 4.17 |
| AZA | 16 | 89.21 ± 0.17 | 84.42 ± 2.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schuchardt, M.; Herrmann, J.; Henkel, C.; Babic, M.; van der Giet, M.; Tölle, M. Long-Term Treatment of Azathioprine in Rats Induces Vessel Mineralization. Biomedicines 2021, 9, 327. https://doi.org/10.3390/biomedicines9030327
Schuchardt M, Herrmann J, Henkel C, Babic M, van der Giet M, Tölle M. Long-Term Treatment of Azathioprine in Rats Induces Vessel Mineralization. Biomedicines. 2021; 9(3):327. https://doi.org/10.3390/biomedicines9030327
Chicago/Turabian StyleSchuchardt, Mirjam, Jaqueline Herrmann, Cornelia Henkel, Milen Babic, Markus van der Giet, and Markus Tölle. 2021. "Long-Term Treatment of Azathioprine in Rats Induces Vessel Mineralization" Biomedicines 9, no. 3: 327. https://doi.org/10.3390/biomedicines9030327
APA StyleSchuchardt, M., Herrmann, J., Henkel, C., Babic, M., van der Giet, M., & Tölle, M. (2021). Long-Term Treatment of Azathioprine in Rats Induces Vessel Mineralization. Biomedicines, 9(3), 327. https://doi.org/10.3390/biomedicines9030327