Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails

 ,
,  ,
, 
Abstract
:1. Introduction
2. Global Antimicrobial Resistance and the Need for Novel Compounds
3. Cone snails as a Source of Novel Antimicrobial Peptides
3.1. Cone Snails Diversity
3.2. Cone Snail’s Venom Composition and Conotoxins Classification
3.3. Antimicrobial Activity of Conidae’s Conopeptides
4. Bioinformatics-Aided Proteotranscriptomics
4.1. Cone Snail Venom Extraction
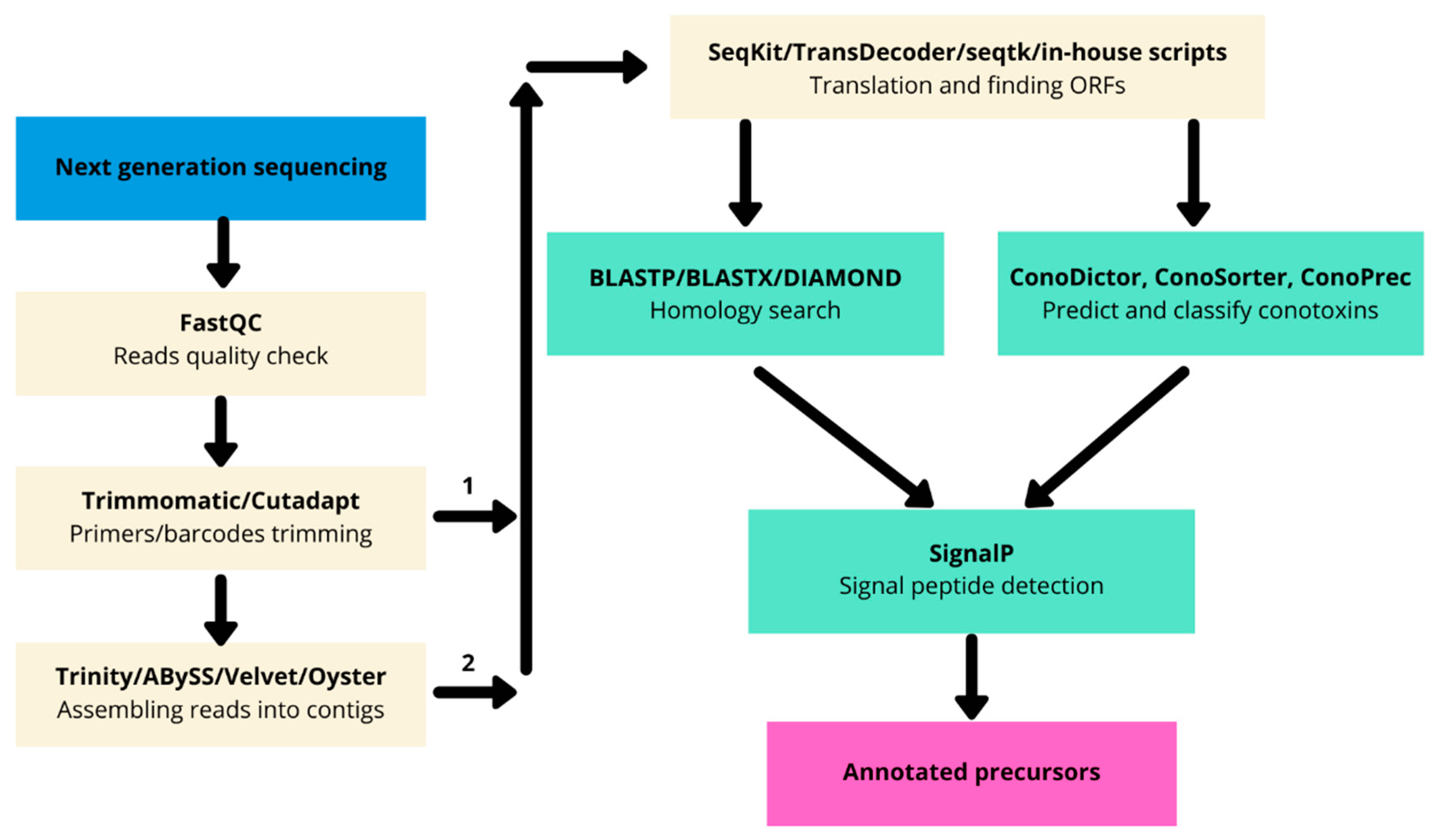
4.2. Next-Generation Transcriptomics Sequencing and Bioinformatics
4.3. Venom Gland Transcriptome Annotation Based on Profile Hidden Markov Models
4.4. Venom Proteomics
4.5. Proteotranscriptomics
4.6. In Silico AMPs Structure Determination
5. Antibacterial Activity Assays
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaynes, R. The Discovery of Penicillin—New Insights After More than 75 Years of Clinical Use. Emerg. Infect. Dis. 2017, 23, 849–853. [Google Scholar] [CrossRef]
- Woodruff, H.B.; Selman, A. Waksman, Winner of the 1952 Nobel Prize for Physiology or Medicine. Appl. Environ. Microbiol. 2014, 80, 2–8. [Google Scholar] [CrossRef] [Green Version]
- Penicillin, S. CDC Control of Infectious Diseases, 1900–1999. JAMA 1999, 282, 1029. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Gallo, R.L. Antimicrobial Peptides. Curr. Biol. 2016, 26, R14–R19. [Google Scholar] [CrossRef]
- O’Neil, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations 2016; Government of the United Kingdom: London, UK, 2016.
- Humphreys, G.; Fleck, F. United Nations Meeting on Antimicrobial Resistance. Bull. World Health Organ. 2016, 94, 638–639. [Google Scholar] [CrossRef]
- Prashanth, J.R.; Brust, A.; Jin, A.-H.; Alewood, P.F.; Dutertre, S.; Lewis, R.J. Cone Snail Venomics: From Novel Biology to Novel Therapeutics. Future Med. Chem. 2014, 6, 1659–1675. [Google Scholar] [CrossRef]
- Bordon, K.D.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Utkin, Y.N. Animal Venom Studies: Current Benefits and Future Developments. World J. Biol. Chem. 2015, 6, 28–33. [Google Scholar] [CrossRef]
- Worms, E.B. World Register of Marine Species. Available online: http://www.marinespecies.org7813 (accessed on 28 January 2021).
- Gao, B.; Peng, C.; Yang, J.; Yi, Y.; Zhang, J.; Shi, Q. Cone Snails: A Big Store of Conotoxins for Novel Drug Discovery. Toxins 2017, 9, 397. [Google Scholar] [CrossRef] [Green Version]
- Eisapoor, S.S.; Jamili, S.; Shahbazzadeh, D.; Mostafavi, P.G.; Bagheri, K.P. A New, High Yield, Rapid, and Cost-Effective Protocol to Deprotection of Cysteine-Rich Conopeptide, Omega-Conotoxin MVIIA. Chem. Biol. Drug Des. 2016, 87, 687–693. [Google Scholar] [CrossRef]
- Rigo, F.K.; Dalmolin, G.D.; Trevisan, G.; Tonello, R.; Silva, M.A.; Rossato, M.F.; Klafke, J.Z.; Cordeiro, M.D.N.; Junior, C.J.C.; Montijo, D.; et al. Effect of ω-Conotoxin MVIIA and Phα1β on Paclitaxel-Induced Acute and Chronic Pain. Pharmacol. Biochem. Behav. 2013, 114, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Herzig, V.; Cristofori-Armstrong, B.; Israel, M.R.; Nixon, S.A.; Vetter, I.; King, G.F. Animal Toxins—Nature’s Evolutionary-Refined Toolkit for Basic Research and Drug Discovery. Biochem. Pharmacol. 2020, 181, 114096. [Google Scholar] [CrossRef]
- Fratini, F.; Cilia, G.; Turchi, B.; Felicioli, A. Insects, Arachnids and Centipedes Venom: A Powerful Weapon against Bacteria. A Literature Review. Toxicon 2017, 130, 91–103. [Google Scholar] [CrossRef] [PubMed]
- Younis, S.; Taj, S.; Rashid, S. Structural Studies of Staphylococcus Aureus Sortase Inhibiton via Conus Venom Peptides. Arch. Biochem. Biophys. 2019, 671, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Bernáldez-Sarabia, J.; Figueroa-Montiel, A.; Dueñas, S.; Cervantes-Luévano, K.; Beltrán, J.; Ortiz, E.; Jiménez, S.; Possani, L.; Paniagua-Solís, J.; Gonzalez-Canudas, J.; et al. The Diversified O-Superfamily in Californiconus Californicus Presents a Conotoxin with Antimycobacterial Activity. Toxins 2019, 11, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de León-Nava, M.A.; Romero-Núñez, E.; Luna-Nophal, A.; Bernáldez-Sarabia, J.; Sánchez-Campos, L.N.; Licea-Navarro, A.F.; Morales-Montor, J.; Muñiz-Hernández, S. In Vitro Effect of the Synthetic Cal14.1a Conotoxin, Derived from Conus Californicus, on the Human Parasite Toxoplasma Gondii. Mar. Drugs 2016, 14, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, U.; Malik Mujaddad-ur-Rehman, N.K.; Fawad, S.A.; Fatima, A. Antibacterial Activity of the Venom of Heterometrus Xanthopus. Indian J. Pharmacol. 2012, 44, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, L.; Li, Z.; Zhang, R.; Wu, Y.; Li, W.; Cao, Z. StCT2, a New Antibacterial Peptide Characterized from the Venom of the Scorpion Scorpiops Tibetanus. Peptides 2012, 36, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Conde, R.; Zamudio, F.Z.; Rodríguez, M.H.; Possani, L.D. Scorpine, an Anti-Malaria and Anti-Bacterial Agent Purified from Scorpion Venom. FEBS Lett. 2000, 471, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Corzo, G.; Escoubas, P.; Villegas, E.; Barnham, K.J.; He, W.; Norton, R.S.; Nakajima, T. Characterization of Unique Amphipathic Antimicrobial Peptides from Venom of the Scorpion Pandinus Imperator. Biochem. J. 2001, 359, 35–45. [Google Scholar] [CrossRef]
- Hernández-Aponte, C.A.; Silva-Sanchez, J.; Quintero-Hernández, V.; Rodríguez-Romero, A.; Balderas, C.; Possani, L.D.; Gurrola, G.B. Vejovine, a New Antibiotic from the Scorpion Venom of Vaejovis Mexicanus. Toxicon 2011, 57, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Benoist, L.; Houyvet, B.; Henry, J.; Corre, E.; Zanuttini, B.; Zatylny-Gaudin, C. In-Depth In Silico Search for Cuttlefish (Sepia Officinalis) Antimicrobial Peptides Following Bacterial Challenge of Haemocytes. Mar. Drugs 2020, 18, 439. [Google Scholar] [CrossRef] [PubMed]
- Canelli, A.P.; Dos Santos Rodrigues, T.F.; de Goes, V.F.F.; Caetano, G.F.; Mazzi, M.V. Evaluation of the Effectiveness of Crotoxin as an Antiseptic against Candida spp. Biofilms. Toxins 2020, 12, 532. [Google Scholar] [CrossRef] [PubMed]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.; Cunha, W.R.; de Almeida, R.; Martins, C.H. Antimicrobial Activity of Apitoxin, Melittin and Phospholipase A2 of Honey Bee (Apis Mellifera) Venom against Oral Pathogens. An. Acad. Bras. Ciênc. 2015, 87, 147–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo-Braga, M.N.; Almeida, F.D.M.; Dos Santos, D.M.; de Avelar Júnior, J.T.; Dos Reis, P.V.M.; de Lima, M.E. Antimicrobial Peptides from Lycosidae (Sundevall, 1833) Spiders. Curr. Protein Pept. Sci. 2020, 21, 527–541. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Zeng, X.-C.; Yang, Y.; Luo, F.; Luo, X.; Wu, S.; Zhang, L.; Zhou, J. A Novel Class of Antimicrobial Peptides from the Scorpion Heterometrus Spinifer. Peptides 2012, 38, 389–394. [Google Scholar] [CrossRef]
- Primon-Barros, M.; Macedo, A.J. Animal Venom Peptides: Potential for New Antimicrobial Agents. Curr. Top. Med. Chem. 2017, 17, 1119–1156. [Google Scholar] [CrossRef]
- Santos, D.M.; Verly, R.M.; Piló-Veloso, D.; de Maria, M.; de Carvalho, M.A.R.; Cisalpino, P.S.; Soares, B.M.; Diniz, C.G.; Farias, L.M.; Moreira, D.F.F.; et al. LyeTx I, a Potent Antimicrobial Peptide from the Venom of the Spider Lycosa Erythrognatha. Amino Acids 2010, 39, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Schneider, R.; Primon-Barros, M.; von Borowski, R.G.; Chat, S.; Nonin-Lecomte, S.; Gillet, R.; Macedo, A.J. Pseudonajide Peptide Derived from Snake Venom Alters Cell Envelope Integrity Interfering on Biofilm Formation in Staphylococcus Epidermidis. BMC Microbiol. 2020, 20, 237. [Google Scholar] [CrossRef]
- Yan, L.; Adams, M.E. Lycotoxins, Antimicrobial Peptides from Venom of the Wolf SpiderLycosa Carolinensis. J. Biol. Chem. 1998, 273, 2059–2066. [Google Scholar] [CrossRef] [Green Version]
- Clardy, J.; Fischbach, M.; Currie, C. The Natural History of Antibiotics. Curr. Biol. 2009, 19, R437–R441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Cunha, B.R.; Fonseca, L.P.; Calado, C.R.C. Antibiotic Discovery: Where Have We Come from, Where Do We Go? Antibiotics 2019, 8, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4, 481–511. [Google Scholar] [CrossRef] [Green Version]
- Manenzhe, R.I.; Zar, H.J.; Nicol, M.P.; Kaba, M. The Spread of Carbapenemase-Producing Bacteria in Africa: A Systematic Review. J. Antimicrob. Chemother. 2015, 70, 23–40. [Google Scholar] [CrossRef] [Green Version]
- Baguy, O.M.; Nathalie, G.K.; David, C.N.G.; Daniel, S.N.G.; Julien, C.K.; Rose, K.N.; Djénéba, O.G.; Valérie, G.; Bertin, T.K.; Mireille, D. First Report of Qnr Genes in Multidrugs Resistant (ESBL) Enterobacteria Isolated from Different Ecosystems in Abidjan, Ivory Coast. Int. J. Biol. Sci. Appl. 2014, 21, 170–175. [Google Scholar]
- Carole, G.M.V.; Kouadio, G.N.; Baguy, O.M.; Djeneba, O.G.; Ayayi, A.; Bertin, T.K.; Anatole, T.A.; Innocent, K.K.; Kpoda, D.S.; Eric, T.; et al. Antimicrobial Resistance Profile and Molecular Characterization of Extended-Spectrum Beta-Lactamase Genes in Enterobacteria Isolated from Human, Animal and Environment. J. Adv. Microbiol. 2018, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Collignon, P.; Wegener, H.C.; Braam, P.; Butler, C.D. The Routine Use of Antibiotics to Promote Animal Growth Does Little to Benefit Protein Undernutrition in the Developing World. Clin. Infect. Dis. 2005, 41, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Addis, Z.; Kebede, N.; Sisay, Z.; Alemayehu, H.; Wubetie, A.; Kassa, T. Prevalence and Antimicrobial Resistance of Salmonella Isolated from Lactating Cows and in Contact Humans in Dairy Farms of Addis Ababa: A Cross Sectional Study. BMC Infect. Dis. 2011, 11, 222. [Google Scholar] [CrossRef] [Green Version]
- Fortini, D.; Fashae, K.; Garcia-Fernandez, A.; Villa, L.; Carattoli, A. Plasmid-Mediated Quinolone Resistance and -Lactamases in Escherichia Coli from Healthy Animals from Nigeria. J. Antimicrob. Chemother. 2011, 66, 1269–1272. [Google Scholar] [CrossRef]
- Kikuvi, G.M.; Ombui, J.N.; Mitema, E.S. Serotypes and Antimicrobial Resistance Profiles of Salmonella Isolates from Pigs at Slaughter in Kenya. J. Infect. Dev. Ctries. 2010, 4, 243–248. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.J.; Thottathil, S.E.; Newman, T.B. Antibiotics Overuse in Animal Agriculture: A Call to Action for Health Care Providers. Am. J. Public Health 2015, 105, 2409–2410. [Google Scholar] [CrossRef]
- Reynolds, L.; McKee, M. Factors Influencing Antibiotic Prescribing in China: An Exploratory Analysis. Health Policy 2009, 90, 32–36. [Google Scholar] [CrossRef]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The Threat of Antimicrobial Resistance in Developing Countries: Causes and Control Strategies. Antimicrob. Resist. Infect. Control 2017, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial Peptides of Multicellular Organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Petersen, A.P.; Lau, C.K.; Jing, W.; Storey, D.G.; Vogel, H.J. Mechanism of Action of Puroindoline Derived Tryptophan-Rich Antimicrobial Peptides. Biochim. Biophys. Acta Biomembr. 2013, 1828, 1802–1813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, R.N. The Mechanism of Inhibition of Protein Synthesis by the Proline-Rich Peptide Oncocin. Mol. Biol. 2015, 22, 466–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Thacker, P.A.; Watford, M.; Qiao, S. Functions of Antimicrobial Peptides in Gut Homeostasis. Curr. Protein Pept. Sci. 2015, 16, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The Value of Antimicrobial Peptides in the Age of Resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef]
- Tan, X.W.; Goh, T.W.; Saraswathi, P.; Nyein, C.L.; Setiawan, M.; Riau, A.; Lakshminarayanan, R.; Liu, S.; Tan, D.; Beuerman, R.W.; et al. Effectiveness of Antimicrobial Peptide Immobilization for Preventing Perioperative Cornea Implant-Associated Bacterial Infection. Antimicrob. Agents Chemother. 2014, 58, 5229–5238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Zhang, H.; Gao, C.; Chen, R.; Li, C. Antimicrobial Mechanism of PBD2 against Staphylococcus Aureus. Molecules 2020, 25, 3513. [Google Scholar] [CrossRef]
- Kumar, P.S. A Perspective on Toxicology of Conus Venom Peptides. Asian Pac. J. Trop. Med. 2015, 8, 337–351. [Google Scholar] [CrossRef] [Green Version]
- Eric, M.; Loïc, L.; Christophe, R.; Alain, R. A Taxonomic Iconography of Living Conidae; ConchBooks: Harxheim, Germany, 2018; Volume 1, ISBN 978-3-939767-91-6. [Google Scholar]
- Olivera, B.M.; Corneli, P.S.; Watkins, M.; Fedosov, A. Biodiversity of Cone Snails and Other Venomous Marine Gastropods: Evolutionary Success Through Neuropharmacology. Annu. Rev. Anim. Biosci. 2014, 2, 487–513. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Lewis, R. Cone Snail Biology, Bioprospecting and Conservation. In Snails: Biology, Ecology and Conservation; Nova Science Publishers, Inc.: New York, NY, USA, 2013; pp. 85–105. [Google Scholar]
- Gao, B.; Peng, C.; Lin, B.; Chen, Q.; Zhang, J.; Shi, Q. Screening and Validation of Highly-Efficient Insecticidal Conotoxins from a Transcriptome-Based Dataset of Chinese Tubular Cone Snail. Toxins 2017, 9, 214. [Google Scholar] [CrossRef]
- Schulz, J.R.; Norton, A.G.; Gilly, W.F. The Projectile Tooth of a Fish-Hunting Cone Snail: Conus Catus Injects Venom into Fish Prey Using a High-Speed Ballistic Mechanism. Biol. Bull. 2004, 207, 77–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duque, H.M.; Dias, S.C.; Franco, O.L. Structural and Functional Analyses of Cone Snail Toxins. Mar. Drugs 2019, 17, 370. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.D.; Norton, R.S. Conotoxin Gene Superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [Green Version]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, Synthesis, and Structure—Activity Relationships of Conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef]
- Kaas, Q.; Westermann, J.-C.; Craik, D.J. Conopeptide Characterization and Classifications: An Analysis Using ConoServer. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus Venom Peptide Pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef]
- Yacoub, T.; Rima, M.; Karam, M.; Sabatier, J.-M.; Fajloun, Z. Antimicrobials from Venomous Animals: An Overview. Molecules 2020, 25, 2402. [Google Scholar] [CrossRef]
- Lebbe, E.K.M.; Ghequire, M.G.K.; Peigneur, S.; Mille, B.G.; Devi, P.; Ravichandran, S.; Waelkens, E.; D’Souza, L.; de Mot, R.; Tytgat, J. Novel Conopeptides of Largely Unexplored Indo Pacific Conus sp. Mar. Drugs 2016, 14, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggs, J.S.; Rosenfeld, Y.; Shai, Y.; Olivera, B.M. Conolysin-Mt: A Conus Peptide That Disrupts Cellular Membranes. Biochemistry 2007, 46, 12586–12593. [Google Scholar] [CrossRef]
- Hemu, X.; Tam, J.P. Macrocyclic Antimicrobial Peptides Engineered from ω-Conotoxin. Curr. Pharm. Des. 2017, 23, 2131–2138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivera, B.M. Conus Peptides: Biodiversity-Based Discovery and Exogenomics. J. Biol. Chem. 2006, 281, 31173–31177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutertre, S.; Jin, A.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep Venomics Reveals the Mechanism for Expanded Peptide Diversity in Cone Snail Venom. Mol. Cell. Proteom. 2013, 12, 312–329. [Google Scholar] [CrossRef] [Green Version]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of Separate Predation- and Defence-Evoked Venoms in Carnivorous Cone Snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.D.; Safavi-Hemami, H.; Raghuraman, S.; Imperial, J.S.; Papenfuss, A.T.; Teichert, R.W.; Purcell, A.W.; Olivera, B.M.; Norton, R.S. Discovery by Proteogenomics and Characterization of an RF-Amide Neuropeptide from Cone Snail Venom. J. Proteom. 2015, 114, 38–47. [Google Scholar] [CrossRef] [Green Version]
- Bergeron, Z.L.; Chun, J.B.; Baker, M.R.; Sandall, D.W.; Peigneur, S.; Yu, P.Y.C.; Thapa, P.; Milisen, J.W.; Tytgat, J.; Livett, B.G.; et al. A ‘Conovenomic’ Analysis of the Milked Venom from the Mollusk-Hunting Cone Snail Conus Textile—The Pharmacological Importance of Post-Translational Modifications. Peptides 2013, 49, 145–158. [Google Scholar] [CrossRef]
- Jin, A.-H.; Vetter, I.; Himaya, S.W.A.; Alewood, P.F.; Lewis, R.J.; Dutertre, S. Transcriptome and Proteome of Conus Planorbis Identify the Nicotinic Receptors as Primary Target for the Defensive Venom. Proteomics 2015, 15, 4030–4040. [Google Scholar] [CrossRef]
- Prashanth, J.R.; Dutertre, S.; Jin, A.H.; Lavergne, V.; Hamilton, B.; Cardoso, F.C.; Griffin, J.; Venter, D.J.; Alewood, P.F.; Lewis, R.J. The Role of Defensive Ecological Interactions in the Evolution of Conotoxins. Mol. Ecol. 2016, 25, 598–615. [Google Scholar] [CrossRef]
- Jin, A.-H.; Dutertre, S.; Dutt, M.; Lavergne, V.; Jones, A.; Lewis, R.J.; Alewood, P.F. Transcriptomic-Proteomic Correlation in the Predation-Evoked Venom of the Cone Snail, Conus Imperialis. Mar. Drugs 2019, 17, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prator, C.A.; Murayama, K.M.; Schulz, J.R. Venom Variation during Prey Capture by the Cone Snail, Conus Textile. PLoS ONE 2014, 9, e98991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, C.; Grilley, M.; Miller, C.; Shon, K.-J.; Cruz, L.J.; Gray, W.R.; Dykert, J.; Rivier, J.; Yoshikami, D.; Olivera, B.M. A New Family of Conus Peptides Targeted to the Nicotinic Acetylcholine Receptor. J. Biol. Chem. 1995, 270, 22361–22367. [Google Scholar] [CrossRef] [Green Version]
- Bingham, J.-P.; Mitsunaga, E.; Bergeron, Z.L. Drugs from Slugs—Past, Present and Future Perspectives of ω-Conotoxin Research. Chem. Biol. Interact. 2010, 183, 1–18. [Google Scholar] [CrossRef]
- Prashanth, J.R.; Lewis, R.J.; Dutertre, S. Towards an Integrated Venomics Approach for Accelerated Conopeptide Discovery. Toxicon 2012, 60, 470–477. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Martin, M. Cutadapt Removes Adapter Sequences from High-Throughput Sequencing Reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Characterization of the Conus Bullatus Genome and Its Venom-Duct Transcriptome. BMC Genom. 2011, 12, 60. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Zerbino, D.R. Using the Velvet de Novo Assembler for Short-Read Sequencing Technologies. Curr. Protoc. Bioinform. 2010, 31, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Simpson, J.T.; Wong, K.; Jackman, S.D.; Schein, J.E.; Jones, S.J.M.; Birol, İ. ABySS: A Parallel Assembler for Short Read Sequence Data. Genome Res. 2009, 19, 1117–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacManes, M.D. The Oyster River Protocol: A Multi-Assembler and Kmer Approach for de Novo Transcriptome Assembly. PeerJ 2018, 6, e5428. [Google Scholar] [CrossRef]
- Rubio, A.; Mier, P.; Andrade-Navarro, M.A.; Garzón, A.; Jiménez, J.; Pérez-Pulido, A.J. CRISPR Sequences Are Sometimes Erroneously Translated and Can Contaminate Public Databases with Spurious Proteins Containing Spaced Repeats. Database 2020, 2020, baaa088. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Kaas, Q.; Yu, R.; Jin, A.-H.; Dutertre, S.; Craik, D.J. ConoServer: Updated Content, Knowledge, and Discovery Tools in the Conopeptide Database. Nucleic Acids Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef] [PubMed]
- Acland, A.; Agarwala, R.; Barrett, T.; Beck, J.; Benson, D.A.; Bollin, C.; Bolton, E.; Bryant, S.H.; Canese, K.; Church, D.M.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2014, 42, D7–D17. [Google Scholar] [CrossRef] [Green Version]
- Consortium, T.U. UniProt: A Worldwide Hub of Protein Knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koua, D.; Brauer, A.; Laht, S.; Kaplinski, L.; Favreau, P.; Remm, M.; Lisacek, F.; Stöcklin, R. ConoDictor: A Tool for Prediction of Conopeptide Superfamilies. Nucleic Acids Res. 2012, 40, W238–W241. [Google Scholar] [CrossRef]
- Lavergne, V.; Dutertre, S.; Jin, A.; Lewis, R.J.; Taft, R.J.; Alewood, P.F. Systematic Interrogation of the Conus Marmoreus Venom Duct Transcriptome with ConoSorter Reveals 158 Novel Conotoxins and 13 New Gene Superfamilies. BMC Genom. 2013, 14, 708. [Google Scholar] [CrossRef] [Green Version]
- Armenteros, J.J.A.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 Improves Signal Peptide Predictions Using Deep Neural Networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The Protein Families Database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A Sequence Logo Generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.M.; Dutertre, S.; Lewis, R.J.; Marí, F. Intraspecific Variations in Conus Purpurascens Injected Venom Using LC/MALDI-TOF-MS and LC-ESI-TripleTOF-MS. Anal. Bioanal. Chem. 2015, 407, 6105–6116. [Google Scholar] [CrossRef]
- Jones, A.; Bingham, J.-P.; Gehrmann, J.; Bond, T.; Loughnan, M.; Atkins, A.; Lewis, R.J.; Alewood, P.F. Isolation and Characterization of Conopeptides by High-Performance Liquid Chromatography Combined with Mass Spectrometry and Tandem Mass Spectrometry. Rapid Commun. Mass Spectrom. 1996, 10, 138–143. [Google Scholar] [CrossRef]
- Romeo, C.; Francesco, L.D.; Oliverio, M.; Palazzo, P.; Massilia, G.R.; Ascenzi, P.; Polticelli, F.; Schininà, M.E. Conus Ventricosus Venom Peptides Profiling by HPLC-MS: A New Insight in the Intraspecific Variation. J. Sep. Sci. 2008, 31, 488–498. [Google Scholar] [CrossRef]
- Abdel-Rahman, M.A.; Abdel-Nabi, I.M.; El-Naggar, M.S.; Abbas, O.A.; Strong, P.N. Intraspecific Variation in the Venom of the Vermivorous Cone Snail Conus Vexillum. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2011, 154, 318–325. [Google Scholar] [CrossRef]
- Bern, M.; Cai, Y.; Goldberg, D. Lookup Peaks: A Hybrid of de Novo Sequencing and Database Search for Protein Identification by Tandem Mass Spectrometry. Anal. Chem. 2007, 79, 1393–1400. [Google Scholar] [CrossRef] [PubMed]
- Zamdborg, L.; LeDuc, R.D.; Glowacz, K.J.; Kim, Y.-B.; Viswanathan, V.; Spaulding, I.T.; Early, B.P.; Bluhm, E.J.; Babai, S.; Kelleher, N.L. ProSight PTM 2.0: Improved Protein Identification and Characterization for Top down Mass Spectrometry. Nucleic Acids Res. 2007, 35, W701–W706. [Google Scholar] [CrossRef]
- Xu, D.; Zhang, Y. Toward Optimal Fragment Generations for Ab Initio Protein Structure Assembly: Ab Initio Fragment Generation. Proteins Struct. Funct. Bioinform. 2013, 81, 229–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Protein Structure Prediction Using Multiple Deep Neural Networks in the 13th Critical Assessment of Protein Structure Prediction (CASP13). Proteins Struct. Funct. Bioinform. 2019, 87, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Improved Protein Structure Prediction Using Potentials from Deep Learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: M07-A10; Approved Standard, 10th ed.; Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2015; ISBN 978-1-56238-987-1. [Google Scholar]
- Luna-Ramirez, K.; Tonk, M.; Rahnamaeian, M.; Vilcinskas, A. Bioactivity of Natural and Engineered Antimicrobial Peptides from Venom of the Scorpions Urodacus Yaschenkoi and U. Manicatus. Toxins 2017, 9, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumerova, N.I.; Al-Sayed, E.; Krivosudský, L.; Čipčić-Paljetak, H.; Verbanac, D.; Rompel, A. Antibacterial Activity of Polyoxometalates Against Moraxella Catarrhalis. Front. Chem. 2018, 6, 336. [Google Scholar] [CrossRef]
- Almaaytah, A.; Qaoud, M.T.; Abualhaijaa, A.; Al-Balas, Q.; Alzoubi, K.H. Hybridization and Antibiotic Synergism as a Tool for Reducing the Cytotoxicity of Antimicrobial Peptides. Infect. Drug Resist. 2018, 11, 835–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Name. | Number of Cysteines | Cysteine Pattern | Disulfide Connectivity |
|---|---|---|---|
| I | 4 | CC-C-C | I–III, II–IV |
| II | 6 | CCC-C-C-C | |
| III | 6 | CC-C-C-CC | (I–IV, II–V, III–VI), (I–VI, II–IV, III–V), (I–V, II–IV, III–VI) |
| IV | 6 | CC-C-C-C-C | I–V, II–III, IV–VI |
| V | 4 | CC-CC | I–III, II–IV |
| VI/VII | 6 | C-C-CC-C-C | I–IV, II–V, III–VI |
| VIII | 10 | C-C-C-C-C-C-C-C-C-C | |
| IX | 6 | C-C-C-C-C-C | I–IV, II–V, III–VI |
| X | 4 | CC-C-C | I–IV, II–III |
| XI | 8 | C-C-CC-CC-C-C | I–IV, II–VI, III–VII, V–VIII |
| XII | 8 | C-C-C-C-CC-C-C | |
| XIII | 8 | C-C-C-CC-C-C-C | |
| XIV | 4 | C-C-C-C | I–III, II–IV |
| XV | 8 | C-C-CC-C-C-C-C | |
| XVI | 4 | C-C-CC | |
| XVII | 8 | C-C-CC-C-CC-C | |
| XVIII | 6 | C-C-CC-CC | |
| XIX | 10 | C-C-C-CCC-C-C-C-C | |
| XX | 10 | C-CC-C-CC-C-C-C-C | |
| XXI | 10 | CC-C-C-C-CC-C-C-C | |
| XXII | 8 | C-C-C-C-C-C-C-C | |
| XXIII | 6 | C-C-C-CC-C | |
| XXIV | 4 | C-CC-C | |
| XXV | 6 | C-C-C-C-CC | |
| XXVI | 8 | C-C-C-C-CC-CC | |
| XXVII | 8 | C-C-C-CCC-C-C | |
| XXVIII | 10 | C-C-C-CC-C-C-C-C-C | |
| XXIX | 8 | CCC-C-CC-C-C | |
| XXX | 10 | C-C-CCC-C-C-C-CC | |
| XXXII | 6 | C-CC-C-C-C | |
| XXXIII | 12 | C-C-C-C-C-C-C-C-C-C-C-C |
| Pharmacological Family | Definition | Conotoxin Representative |
|---|---|---|
| α (alpha) | Nicotinic acetylcholine receptors | Alpha-conotoxin GIA (P01519) |
| γ (gamma) | Neuronal pacemaker cation currents (inward cation current) | Gamma-conotoxin PnVIIA (P56711) |
| δ (delta) | Voltage-gated Na channels (agonist, delay inactivation) | Delta-conotoxin TxVIA (Q9U655) |
| ε (epsilon) | Presynaptic Ca channels or G protein-coupled presynaptic receptors | Epsilon-conotoxin TxVA (P81755) |
| ι (iota) | Voltage-gated Na channels (agonist, no delayed inactivation) | Iota-conotoxin RXIA (Q7Z094) |
| κ (kappa) | Voltage-gated K channels (blocker) | Kappa-conotoxin PVIIA (P56633) |
| µ (mu) | Voltage-gated Na channels (antagonist, blocker) | Mu-conotoxin GIIIA (P01523) |
| ρ (rho) | Alpha1-adrenoceptors (GPCR) | Rho-conotoxin TIA (P58811) |
| ς (sigma) | Serotonin-gated ion channels 5-HT3 | Sigma-conotoxin GVIIIA (P58924) |
| τ (tau) | Somatostatin receptor | Tau-conotoxin CnVA (P0DJL6) |
| χ (chi) | Neuronal noradrenaline transporter | Chi-conotoxin MrIA (P58808) |
| ω (omega) | Voltage-gated Ca channels (blocker) | Omega-conotoxin GVIA (P01522) |
| Accession | ID | Description |
|---|---|---|
| PF16981 | Chi-conotoxin | chi-Conotoxin or t superfamily |
| PF02950 | Conotoxin | Conotoxin |
| PF17557 | Conotoxin_I2 | I2-superfamily conotoxins |
| PF05374 | Mu-conotoxin | Mu-Conotoxin |
| PF07473 | Toxin_11 | Spasmodic peptide gm9a; conotoxin from Conus species |
| PF07829 | Toxin_14 | Alpha-A conotoxin PIVA-like protein |
| PF08087 | Toxin_18 | Conotoxin O-superfamily |
| PF08088 | Toxin_19 | Conotoxin I-superfamily |
| PF08094 | Toxin_24 | Conotoxin TVIIA/GS family |
| PF08097 | Toxin_26 | Conotoxin T-superfamily |
| PF07365 | Toxin_8 | Alpha conotoxin precursor |
| Conotoxin Superfamily | pHMM | Cysteine Framework | Sequence Logo |
|---|---|---|---|
| A | CN_A | I, II, IV, VI/VII, XIV, XXII |  |
| B | CN_B | (conantokins, disulfide-poor) |  |
| D | CN_D | XXVIII, IV, XIV, XV, XX, XXIV |  |
| H | CN_H | VI/VII |  |
| I1 | CN_I1 | VI/VII, XI, XXII |  |
| I2 | CN_I2 | VI/VII, XI, XII, XIII, XIV |  |
| I3 | CN_I3 | VI/VII, XI |  |
| J | CN_J | XIV |  |
| K | CN_K | XXIII |  |
| L | CN_L | XIV, XXIV |  |
| M | CN_M | XXXII, I, II, III, IV, VI/VII, IX, XIV, XVI |  |
| N | CN_N | XV |  |
| O1 | CN_O1 | XXIX, I, VI/VII, IX, XII, XIV, XVI |  |
| O2 | CN_O2 | VI/VII, XIV, XV, XVI |  |
| O3 | CN_O3 | VI/VII, XVI |  |
| P | CN_P | IX, XIV |  |
| R | CN_R | XIV |  |
| S | CN_S | XXXIII, VIII |  |
| T | CN_T | I, V, X, XVI |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ebou, A.; Koua, D.; Addablah, A.; Kakou-Ngazoa, S.; Dutertre, S. Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails. Biomedicines 2021, 9, 344. https://doi.org/10.3390/biomedicines9040344
Ebou A, Koua D, Addablah A, Kakou-Ngazoa S, Dutertre S. Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails. Biomedicines. 2021; 9(4):344. https://doi.org/10.3390/biomedicines9040344
Chicago/Turabian StyleEbou, Anicet, Dominique Koua, Audrey Addablah, Solange Kakou-Ngazoa, and Sébastien Dutertre. 2021. "Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails" Biomedicines 9, no. 4: 344. https://doi.org/10.3390/biomedicines9040344
APA StyleEbou, A., Koua, D., Addablah, A., Kakou-Ngazoa, S., & Dutertre, S. (2021). Combined Proteotranscriptomic-Based Strategy to Discover Novel Antimicrobial Peptides from Cone Snails. Biomedicines, 9(4), 344. https://doi.org/10.3390/biomedicines9040344