
Generation of Stilbene Glycoside with Promising Cell Rejuvenation Activity through Biotransformation by the Entomopathogenic Fungus Beauveria bassiana

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Metabolites of RSV Manufactured by B. bassiana
2.3. HPLC Analysis of RSV Metabolites
2.4. Isolation of RSV Metabolites
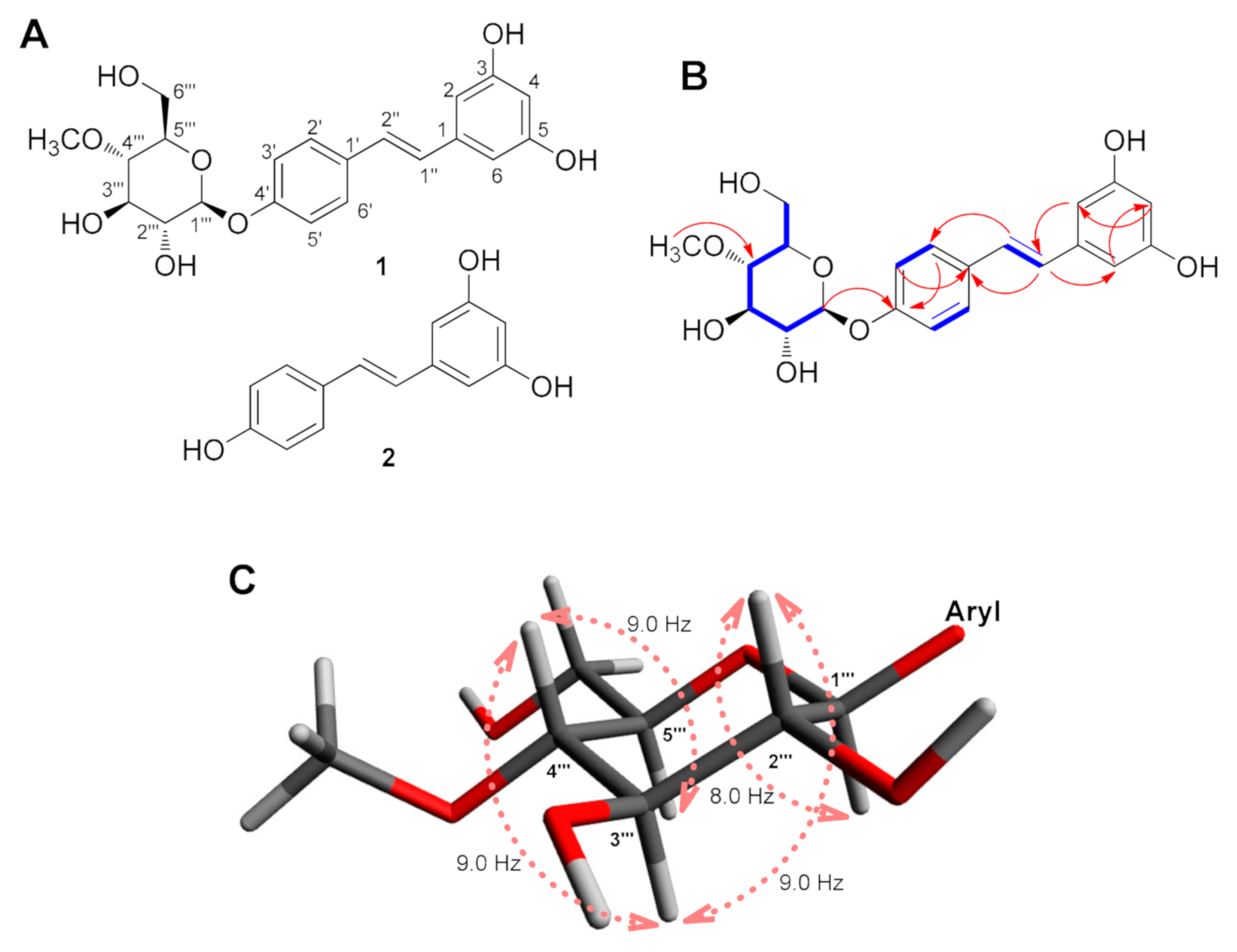
Resvebassianol A (1)
2.5. Acid Hydrolysis of 1
2.6. Microarray
2.7. Cell Culture
2.8. Cell Viability
2.9. Enzyme-Linked Immunosorbent Assay (ELISA)
2.10. Cell Proliferation and Migration Assay
2.11. Statistical Analysis
3. Results and Discussion
3.1. Identification of RSV Metabolite with 4-O-Methyl-D-Glucopyranose through Biotransformation by Beauveria bassiana
3.2. Structural Elucidation of Resvebassianol A (1)
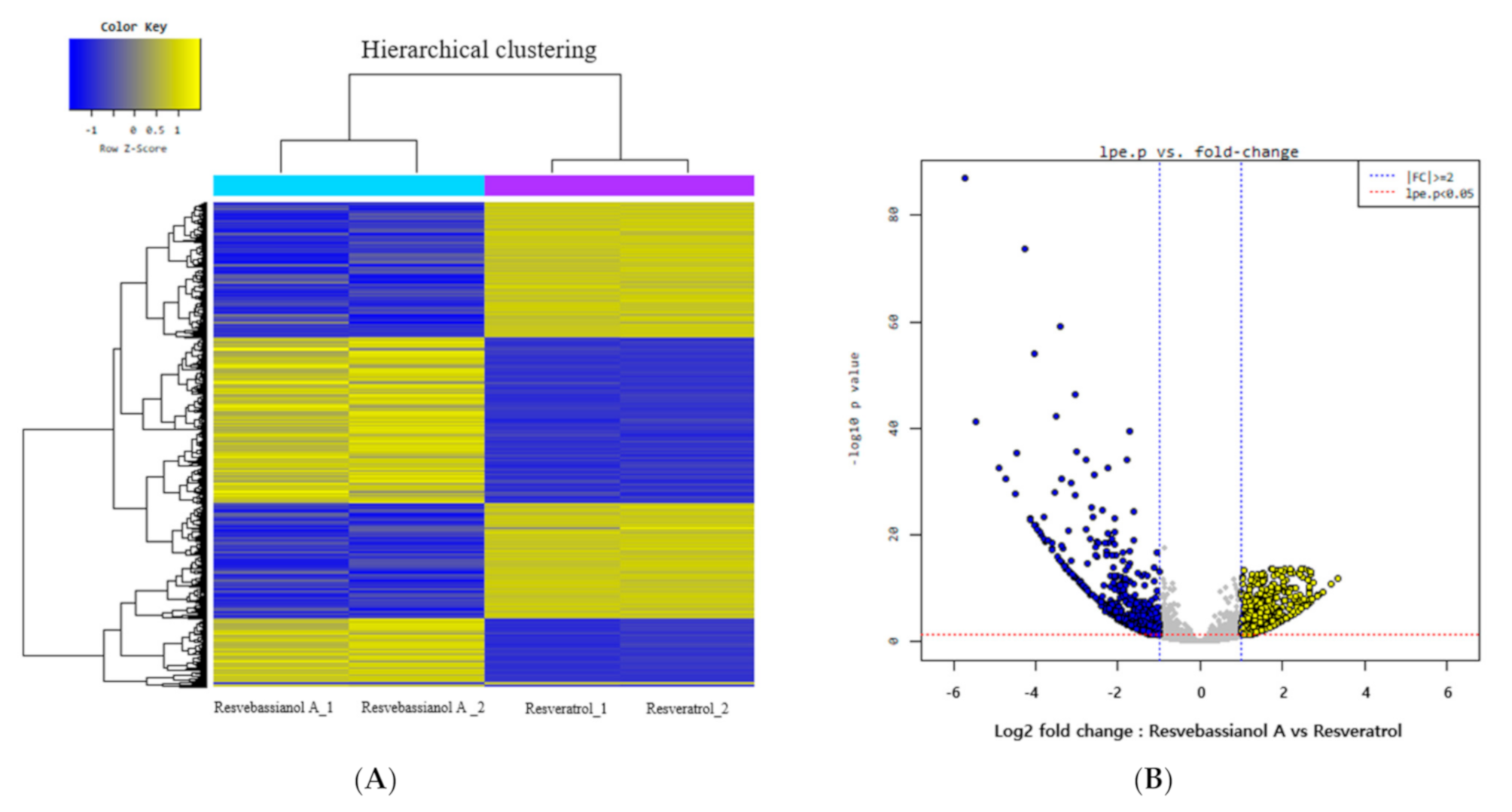
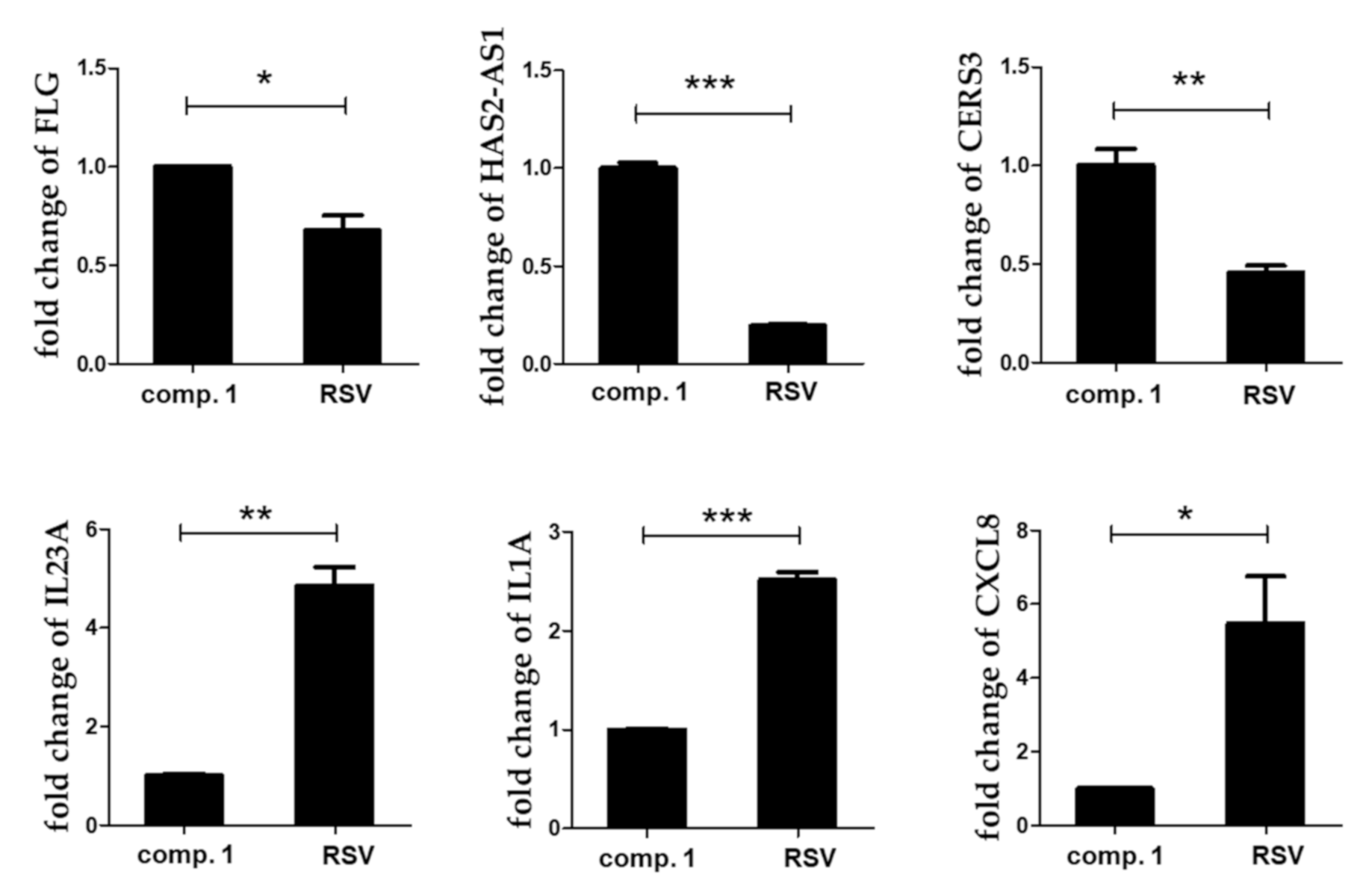
3.3. Microarray Analysis to Determine the Functional Differences between Resvebassianol A and RSV
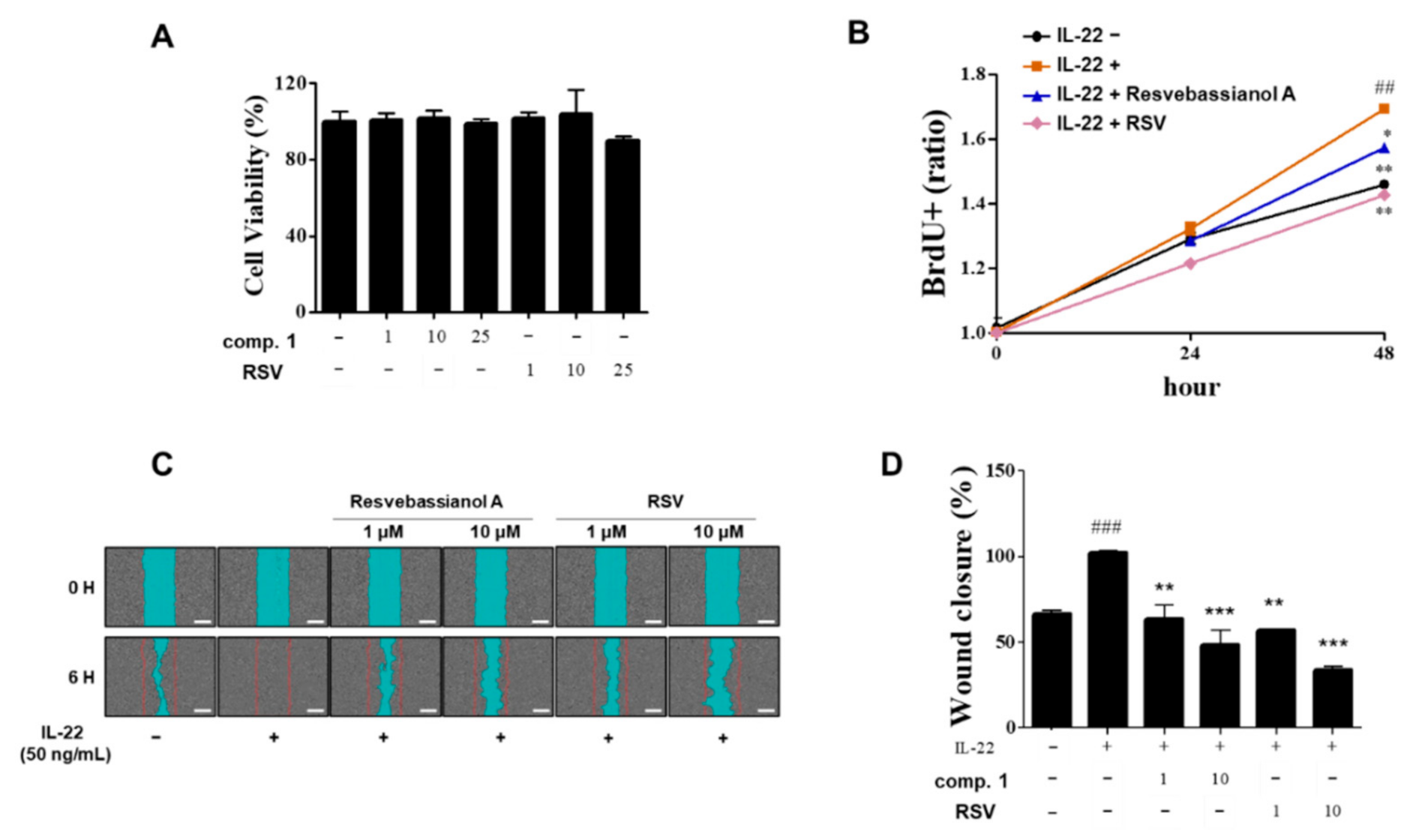
3.4. Effects of Resvebassianol A on the Proliferation and Migration of HaCaT Cells
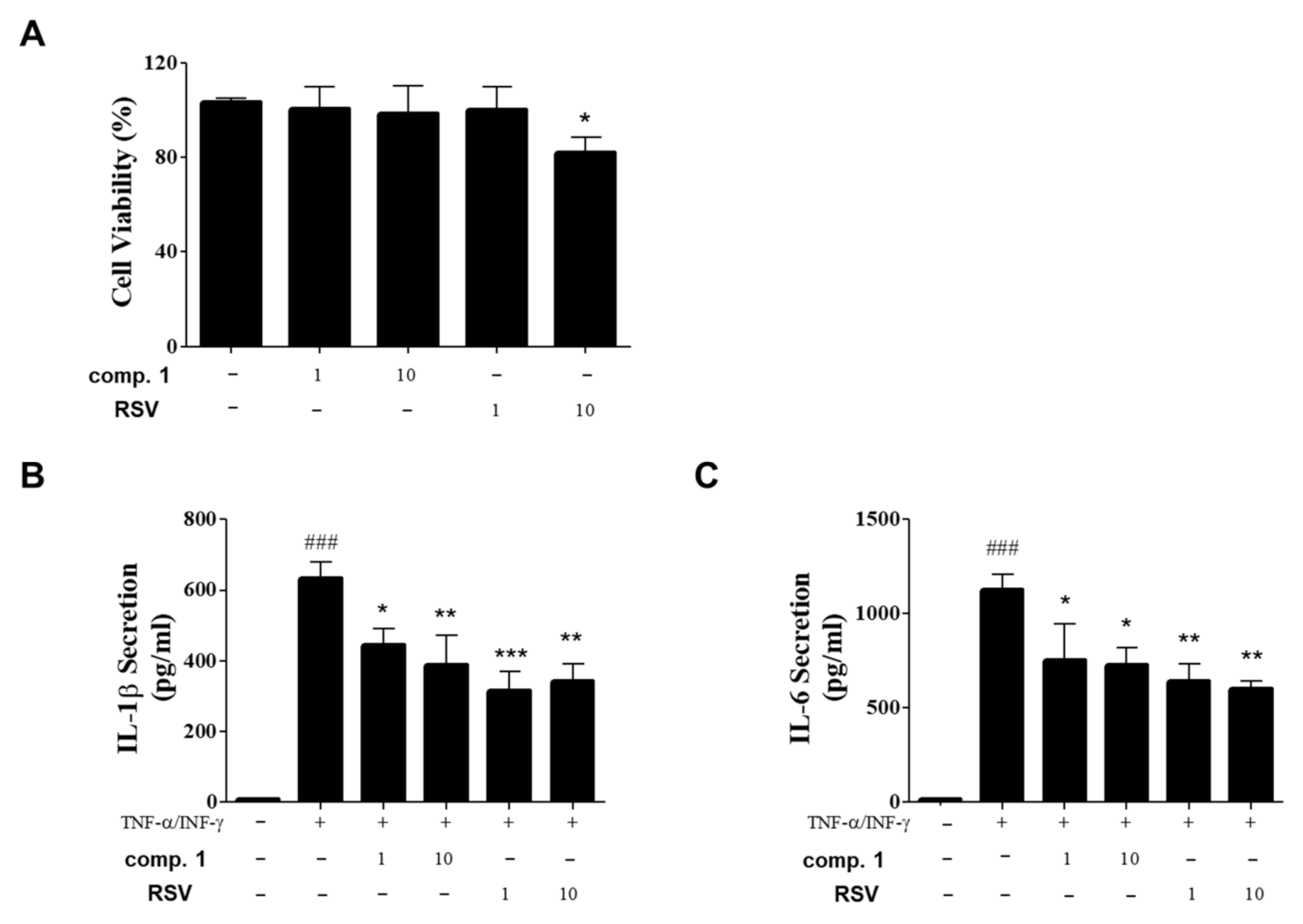
3.5. Inhibitory Effects of Resvebassianol A on the Inflammatory Cytokine Expression of TNF-α/INF-γ-Induced HIEC-6 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Xia, N.; Daiber, A.; Förstermann, U.; Li, H. Antioxidant effects of resveratrol in the cardiovascular system. Br. J. Pharmacol. 2017, 174, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Tang, F.; Liu, Q.; Xiao, J.; Cao, H.; Chen, X. Inhibition of resveratrol glucosides (REs) on advanced glycation endproducts (AGEs) formation: Inhibitory mechanism and structure-activity relationship. Nat. Prod. Res. 2020, 34, 2490–2494. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Suleria, H.A.R.; Ahmad, B.; Peters, D.G.; Mubarak, M.S. A comprehensive review of the health perspectives of resveratrol. Food Funct. 2017, 8, 4284–4305. [Google Scholar] [CrossRef] [PubMed]
- Cottart, C.H.; Nivet-Antoine, V.; Laguillier-Morizot, C.; Beaudeux, J.L. Resveratrol bioavailability and toxicity in humans. Mol. Nutr. Food Res. 2010, 54, 7–16. [Google Scholar] [CrossRef]
- Gambini, J.; Inglés, M.; Olaso, G.; Lopez-Grueso, R.; Bonet-Costa, V.; Gimeno-Mallench, L.; Mas-Bargues, C.; Abdelaziz, K.M.; Gomez-Cabrera, M.C.; Borras, C. Properties of resveratrol: In vitro and in vivo studies about metabolism, bioavailability, and biological effects in animal models and humans. Oxid. Med. Cell. Longev. 2015, 2015, 837042. [Google Scholar] [CrossRef] [Green Version]
- Howells, L.M.; Berry, D.P.; Elliott, P.J.; Jacobson, E.W.; Hoffmann, E.; Hegarty, B.; Brown, K.; Steward, W.P.; Gescher, A.J. Phase I randomized, double-blind pilot study of micronized resveratrol (SRT501) in patients with hepatic metastases—Safety, pharmacokinetics, and pharmacodynamics. Cancer Prev. Res. 2011, 4, 1419–1425. [Google Scholar] [CrossRef] [Green Version]
- de Carvalho, C.C.C.R.; da Fonseca, M.M.R. Biotransformation of terpenes. Biotechnol. Adv. 2006, 24, 134–142. [Google Scholar] [CrossRef]
- Hu, X.; Liu, Y.; Li, D.; Feng, W.; Ni, H.; Cao, S.; Lu, F.; Li, Y. An innovative biotransformation to produce resveratrol by Bacillus safensis. RSC Adv. 2019, 9, 15448–15456. [Google Scholar] [CrossRef] [Green Version]
- Cichewicz, R.H.; Kouzi, S.A.; Hamann, M.T. Dimerization of resveratrol by the grapevine pathogen Botrytis cinerea. J. Nat. Prod. 2000, 63, 29–33. [Google Scholar] [CrossRef]
- Cichewicz, R.H.; Kouzi, S.A. Biotransformation of resveratrol to piceid by Bacillus cereus. J. Nat. Prod. 1998, 61, 1313–1314. [Google Scholar] [CrossRef]
- Wang, L.; Wu, Y.; Chen, Y.; Zou, J.; Li, X. Biotransformation of resveratrol: New prenylated trans-resveratrol synthesized by Aspergillus sp. SCSIOW2. Molecules 2016, 21, 883. [Google Scholar] [CrossRef] [Green Version]
- Westwood, G.S.; Huang, S.-W.; Keyhani, N.O. Allergens of the entomopathogenic fungus Beauveria bassiana. Clin. Mol. Allergy 2005, 3, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasikumar, P.; Lekshmy, K.; Sini, S.; Prabha, B.; Kumar, N.A.; Sivan, V.V.; Jithin, M.M.; Jayamurthy, P.; Shibi, I.G.; Radhakrishnan, K.V. Isolation and characterization of resveratrol oligomers from the stem bark of Hopea ponga (Dennst.) Mabb. and their antidiabetic effect by modulation of digestive enzymes, protein glycation and glucose uptake in L6 myocytes. J. Ethnopharmacol. 2019, 236, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Isaka, M.; Haritakun, R.; Supothina, S.; Choowong, W.; Mongkolsamrit, S. N-Hydroxypyridone alkaloids, chromone derivatives, and tetrahydroxanthones from the scale-insect pathogenic fungus Orbiocrella sp. BCC 33248. Tetrahedron 2014, 70, 9198–9203. [Google Scholar] [CrossRef]
- Bunyapaiboonsri, T.; Yoiprommarat, S.; Intereya, K.; Kocharin, K. New diphenyl ethers from the insect pathogenic fungus Cordyceps sp. BCC 1861. Chem. Pharm. Bull. 2007, 55, 304–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuephadungphan, W.; Helaly, S.E.; Daengrot, C.; Phongpaichit, S.; Luangsa-Ard, J.J.; Rukachaisirikul, V.; Stadler, M. Akanthopyrones A–D, α-pyrones bearing a 4-O-methyl-β-D-glucopyranose moiety from the spider-associated Ascomycete Akanthomyces novoguineensis. Molecules 2017, 22, 1202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isaka, M.; Palasarn, S.; Supothina, S.; Komwijit, S.; Luangsa-ard, J.J. Bioactive compounds from the scale insect pathogenic fungus Conoideocrella tenuis BCC 18627. J. Nat. Prod. 2011, 74, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Kornsakulkarn, J.; Thongpanchang, C.; Lapanun, S.; Srichomthong, K. Isocoumarin glucosides from the scale insect fungus Torrubiella tenuis BCC 12732. J. Nat. Prod. 2009, 72, 1341–1343. [Google Scholar] [CrossRef]
- Kornsakulkarn, J.; Saepua, S.; Srichomthong, K.; Supothina, S.; Thongpanchang, C. New mycotoxins from the scale insect fungus Aschersonia coffeae Henn. BCC 28712. Tetrahedron 2012, 68, 8480–8486. [Google Scholar] [CrossRef]
- Zhang, L.; Fasoyin, O.E.; Molnár, I.; Xu, Y. Secondary metabolites from hypocrealean entomopathogenic fungi: Novel bioactive compounds. Nat. Prod. Rep. 2020, 37, 1181–1206. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Zhang, L.; Wang, C.; Wang, X.; Xu, Y.-M.; Yu, H.; Wu, P.; Li, S.; Han, L.; Gunatilaka, A.A.L.; et al. Methylglucosylation of aromatic amino and phenolic moieties of drug-like biosynthons by combinatorial biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, E4980–E4989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraczek, M.; Szumiło, J.; Podlodowska, J.; Burdan, F. Resveratrol—Phytophenol with wide activity. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2012, 32, 143–146. [Google Scholar]
- Kang, M.C.; Cho, K.; Lee, J.H.; Subedi, L.; Yumnam, S.; Kim, S.Y. Effect of resveratrol-enriched rice on skin inflammation and pruritus in the NC/Nga mouse model of atopic dermatitis. Int. J. Mol. Sci. 2019, 20, 1428. [Google Scholar] [CrossRef] [Green Version]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.H.; Kim, S.Y. Resveratrol-enriched rice attenuates UVB-ROS-induced skin aging via downregulation of inflammatory cascades. Oxid. Med. Cell. Longev. 2017, 2017, 8379539. [Google Scholar] [CrossRef]
- Zhou, F.; Huang, X.; Pan, Y.; Cao, D.; Liu, C.; Liu, Y.; Chen, A. Resveratrol protects HaCaT cells from ultraviolet B-induced photoaging via upregulation of HSP27 and modulation of mitochondrial caspase-dependent apoptotic pathway. Biochem. Biophys. Res. Commun. 2018, 499, 662–668. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef]
- Chimento, A.; de Amicis, F.; Sirianni, R.; Sinicropi, M.S.; Puoci, F.; Casaburi, I.; Saturnino, C.; Pezzi, V. Progress to improve oral bioavailability and beneficial effects of resveratrol. Int. J. Mol. Sci. 2019, 20, 1381. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.M.; Liu, Z.H.; Liu, K.X. Progress in regulation of drug transporters and metabolic enzymes by resveratrol. Yao Xue Xue Bao Acta Pharm. Sin. 2016, 51, 860–865. [Google Scholar]
- Subedi, L.; Baek, S.H.; Kim, S.Y. Bioactive compounds of food: Their role in the prevention and treatment of diseases. Oxid. Med. Cell. Longev. 2018, 2018, 8092713. [Google Scholar]
- Huang, G.; Lv, M.; Hu, J.; Huang, K.; Xu, H. Glycosylation and activities of natural products. Mini Rev. Med. Chem. 2016, 16, 1013–1016. [Google Scholar] [CrossRef]
- Patocka, J. Bioactive metabolites of entomopathogenic fungi Beauveria bassiana. Mil. Med. Sci. Lett. 2016, 85, 80–88. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Position | δH | δC |
|---|---|---|
| 1 | 141.0 | |
| 2 | 6.46 d (2.0) | 106.0 |
| 3 | 159.8 | |
| 4 | 6.18 t (2.0) | 103.1 |
| 5 | 159.8 | |
| 6 | 6.46 d (2.0) | 106.0 |
| 1′ | 133.2 | |
| 2′ | 7.45 d (8.5) | 128.7 |
| 3′ | 7.07 d (8.5) | 118.0 |
| 4′ | 158.8 | |
| 5′ | 7.07 d (8.5) | 118.0 |
| 6′ | 7.45 d (8.5) | 128.7 |
| 1″ | 6.88 d (16.0) | 128.7 |
| 2″ | 6.99 d (16.0) | 129.1 |
| 1′′′ | 4.90 d (8.0) | 102.2 |
| 2′′′ | 3.47 dd (9.0, 8.0) | 75.1 |
| 3′′′ | 3.57 t (9.0) | 78.1 |
| 4′′′ | 3.21 t (9.0) | 80.7 |
| 5′′′ | 3.43 ddd (9.0, 5.0, 2.0) | 77.3 |
| 6′′′ | 3.71 dd (12.0, 5.0); 3.86 dd (12.0, 2.0) | 62.2 |
| 4′′′-OCH3 | 3.59 s | 61.1 |
| Gene Symbol | RefSeq | Gene Name | Fold-Change |
|---|---|---|---|
| CCL17 | NM_002987 | Chemokine (C-C motif) ligand 17 | 4.79 |
| FLG | NM_002016 | Filaggrin | 4.02 |
| HAS2-AS1 | NR_002835 | HAS2 antisense RNA 1 | 3.43 |
| IL17RE | NM_153483 | Interleukin 17 receptor E | 3.13 |
| TLR3 | NM_003265 | Toll-like receptor 3 | 3.00 |
| CERS3 | NM_178842 | Ceramide synthase 3 | 2.62 |
| IL11RA | NM_001142784 | Interleukin 11 receptor, alpha | −2.24 |
| IL7R | NM_002185 | Interleukin 7 receptor | −2.62 |
| IL32 | NM_001012631 | Interleukin 32 | −3.53 |
| IL1A | NM_000575 | Interleukin 1, alpha | −3.66 |
| CXCL8 | NM_000584 | Chemokine (C-X-C motif) ligand 8 | −3.90 |
| IL23A | NM_016584 | Interleukin 23, alpha subunit p19 | −7.53 |
| IL4I1 | NM_152899 | Interleukin 4 induced 1 | −9.30 |
| IL11 | NM_000641 | Interleukin 11 | −13.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ha, S.K.; Kang, M.C.; Lee, S.; Darlami, O.; Shin, D.; Choi, I.; Kim, K.H.; Kim, S.Y. Generation of Stilbene Glycoside with Promising Cell Rejuvenation Activity through Biotransformation by the Entomopathogenic Fungus Beauveria bassiana. Biomedicines 2021, 9, 555. https://doi.org/10.3390/biomedicines9050555
Ha SK, Kang MC, Lee S, Darlami O, Shin D, Choi I, Kim KH, Kim SY. Generation of Stilbene Glycoside with Promising Cell Rejuvenation Activity through Biotransformation by the Entomopathogenic Fungus Beauveria bassiana. Biomedicines. 2021; 9(5):555. https://doi.org/10.3390/biomedicines9050555
Chicago/Turabian StyleHa, Sang Keun, Min Cheol Kang, Seulah Lee, Om Darlami, Dongyun Shin, Inwook Choi, Ki Hyun Kim, and Sun Yeou Kim. 2021. "Generation of Stilbene Glycoside with Promising Cell Rejuvenation Activity through Biotransformation by the Entomopathogenic Fungus Beauveria bassiana" Biomedicines 9, no. 5: 555. https://doi.org/10.3390/biomedicines9050555
APA StyleHa, S. K., Kang, M. C., Lee, S., Darlami, O., Shin, D., Choi, I., Kim, K. H., & Kim, S. Y. (2021). Generation of Stilbene Glycoside with Promising Cell Rejuvenation Activity through Biotransformation by the Entomopathogenic Fungus Beauveria bassiana. Biomedicines, 9(5), 555. https://doi.org/10.3390/biomedicines9050555