
Microbiota Depletion Promotes Human Rotavirus Replication in an Adult Mouse Model

 , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Rotavirus Stock Preparation
2.2. Donor Microbiota Preparation
2.3. Antibiotic Treatment and Microbiota Stool Transplant
2.4. Rotavirus Challenge
2.5. Quantification of Rotavirus from Stool Samples by RT-qPCR
2.6. Quantification of Cytokine and Glycosyltransferase mRNA Expression Level
2.7. Microbiota Profiling in Mice Stool Samples
3. Results
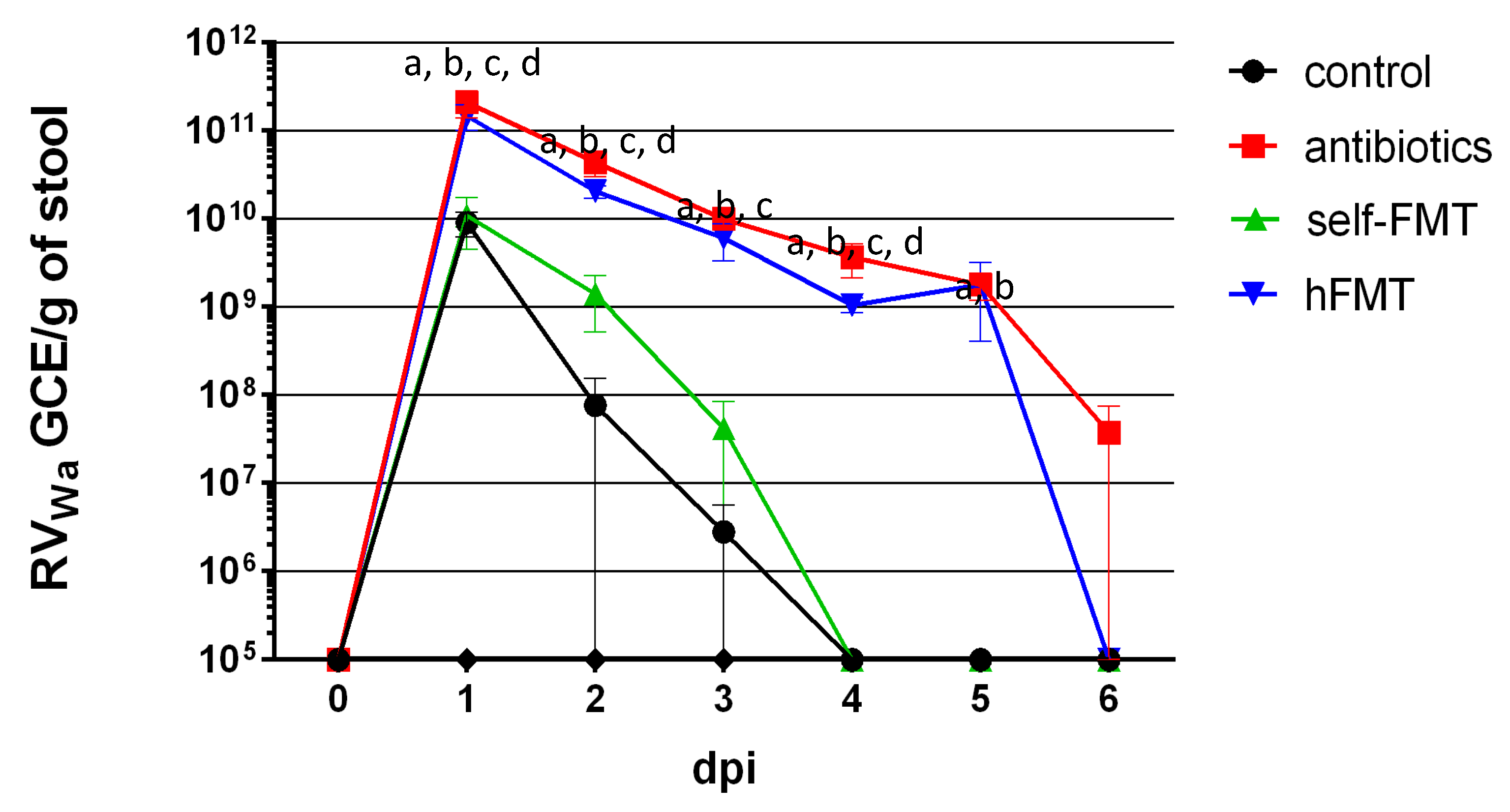
3.1. Mice with Depleted Microbiota and Transplanted with Human Feces Shed Wa Rotavirus for 6 Days
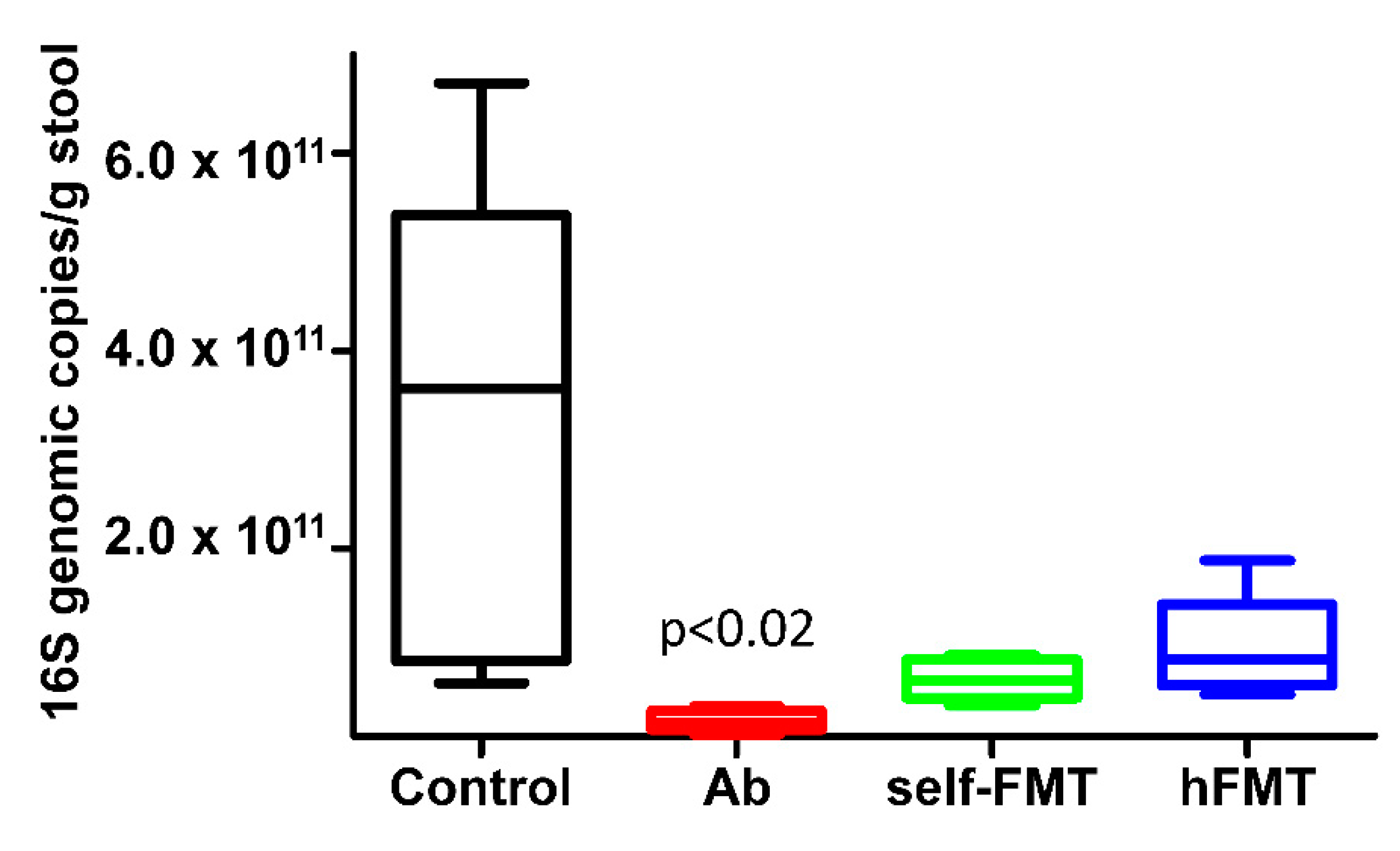
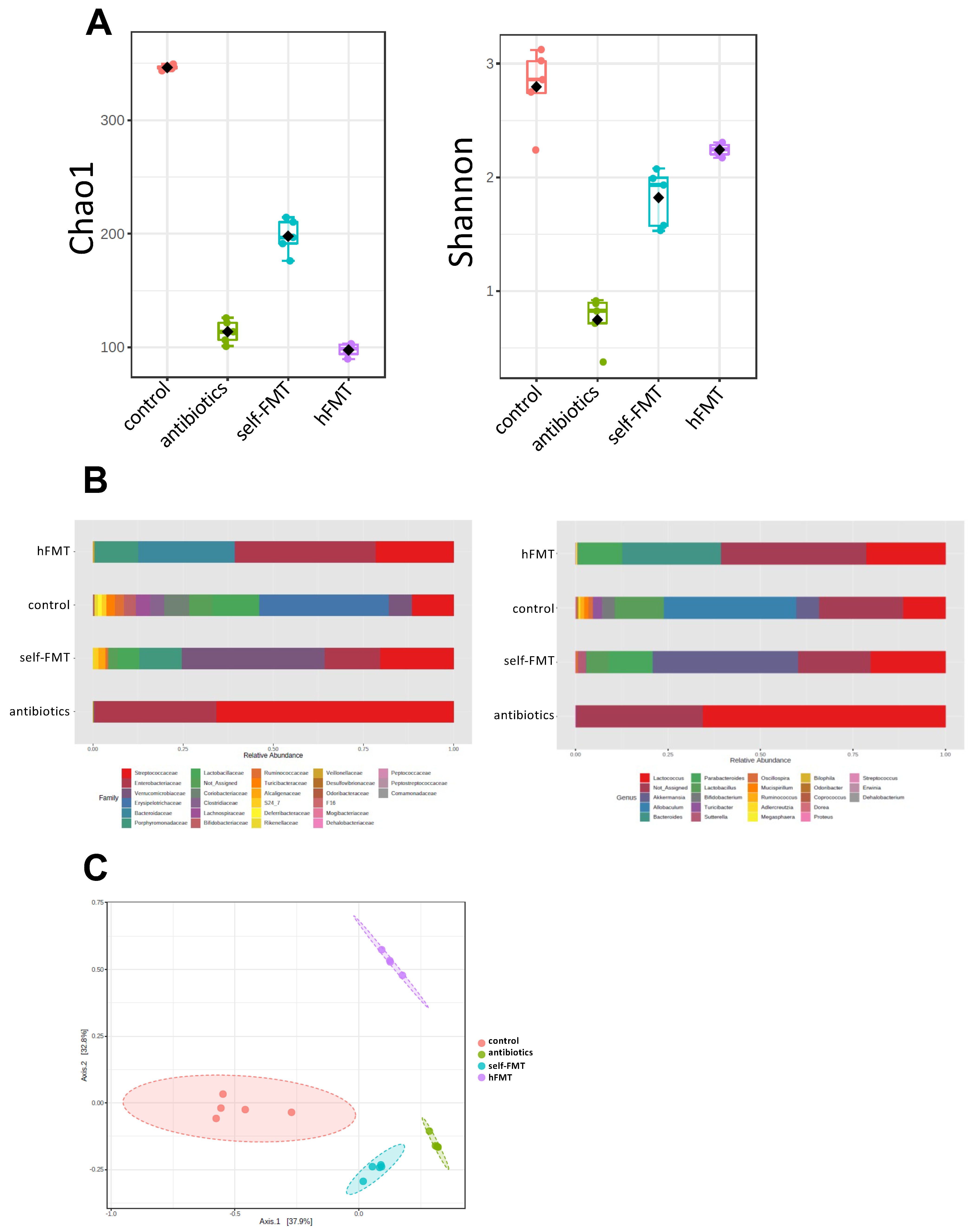
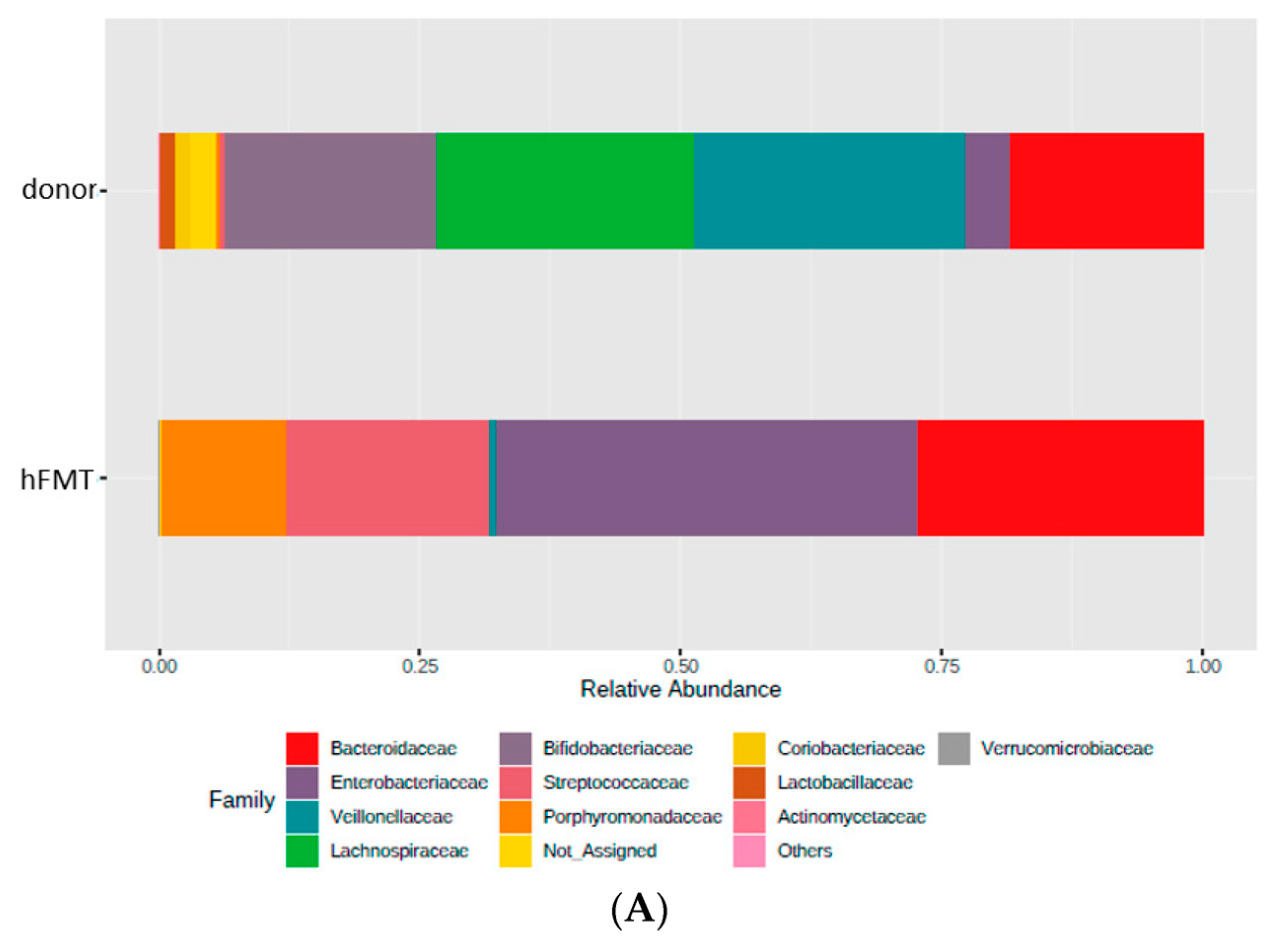
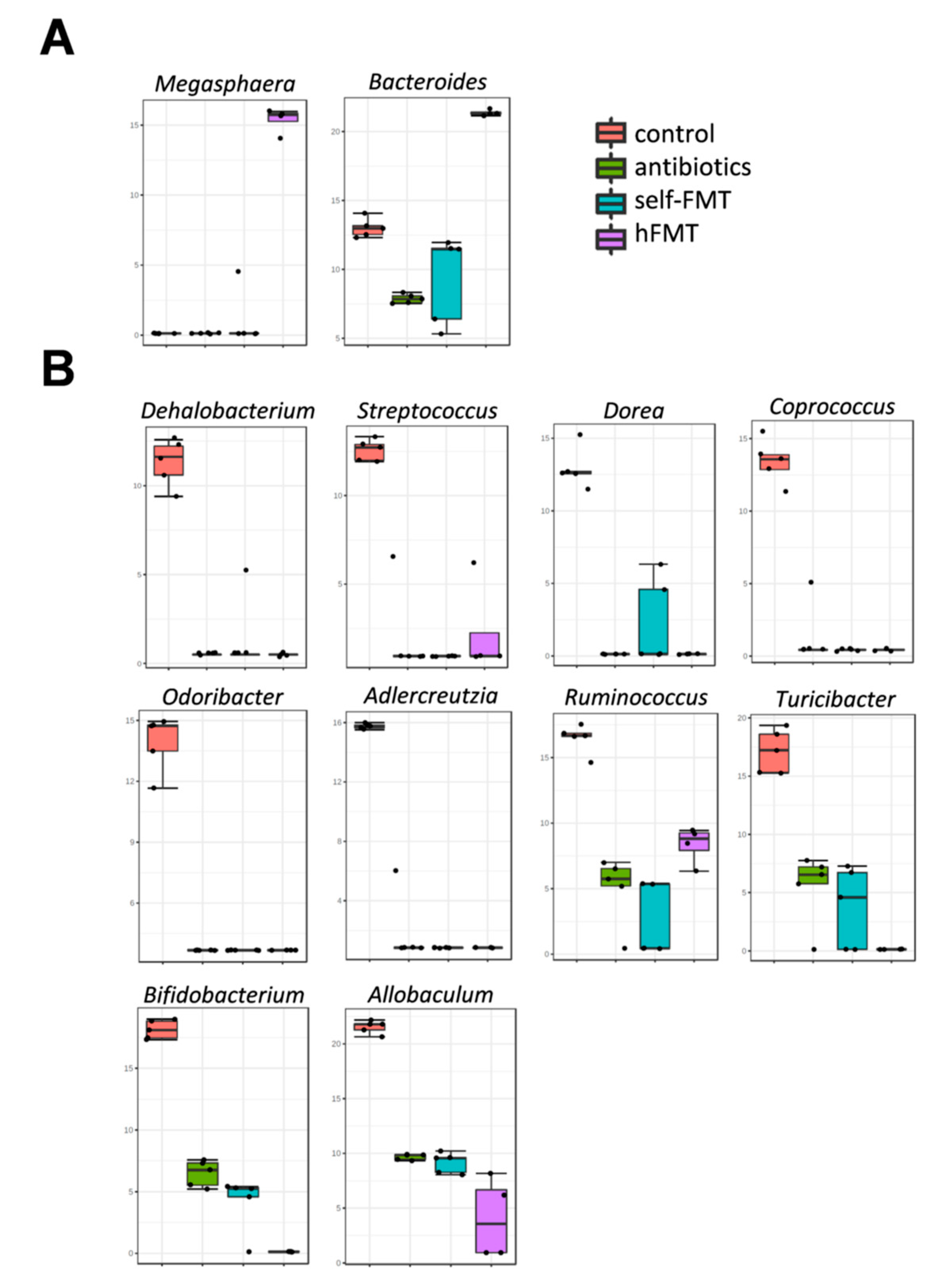
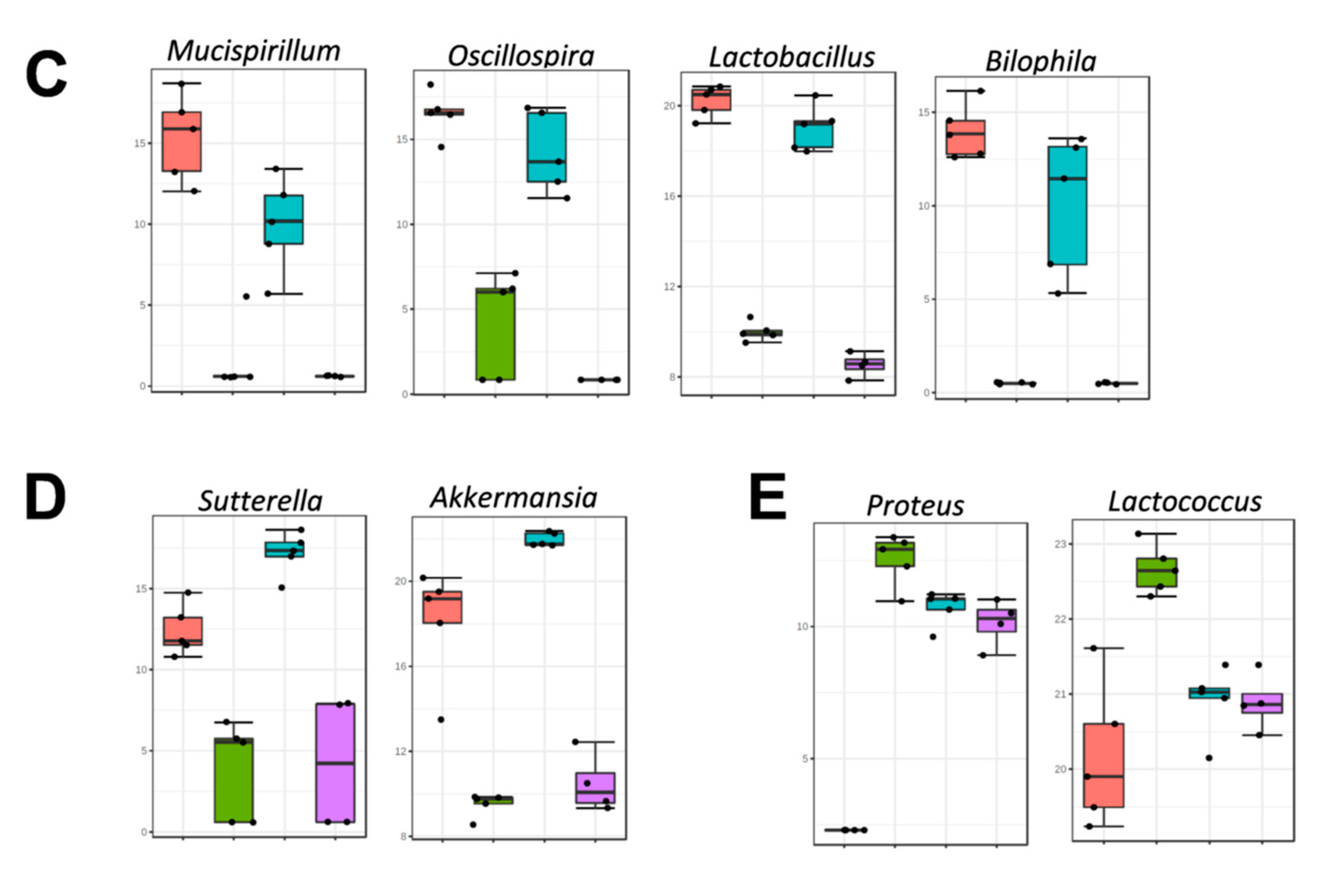
3.2. Impact of Fecal Microbiota Transplant on Mouse Microbiota
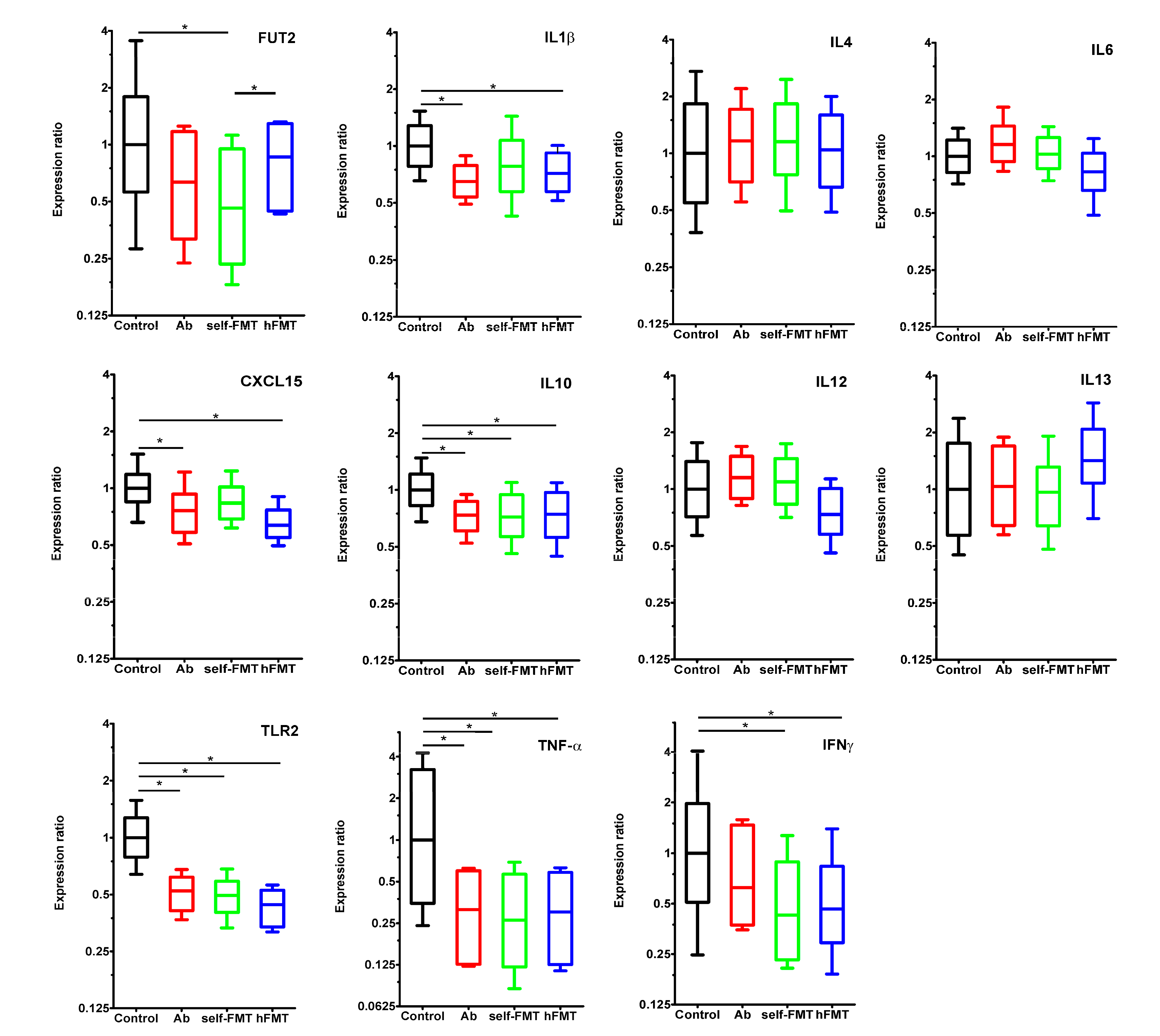
3.3. Immune and Epithelial Glycosylation Gene Expression in the Gut Associates with Human Rotavirus Infection in Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, H.; Naghavi, M.; Allen, C.; Barber, R.M.; Bhutta, Z.A.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; Coates, M.M.; et al. Global, regional, and national life expectancy, all-cause mortality, and cause-specific mortality for 249 causes of death, 1980-2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1459–1544. [Google Scholar] [CrossRef] [Green Version]
- Monedero, V.; Collado, M.C.; Rodríguez-Díaz, J. Therapeutic opportunities in intestinal microbiota-virus interactions. Trends Biotechnol. 2018, 36, 645–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monedero, V.; Buesa, J.; Rodríguez-Díaz, J. The interactions between host glycobiology, bacterial microbiota, and viruses in the gut. Viruses 2018, 10, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, A.H.J.; Hogarty, M.P.; Harris, V.C.; Baldridge, M.T. The complex interactions between rotavirus and the gut microbiota. Front. Cell. Infect. Microbiol. 2021, 10, 820. [Google Scholar] [CrossRef] [PubMed]
- Roth, A.N.; Grau, K.R.; Karst, S.M. Diverse mechanisms underlie enhancement of enteric viruses by the mammalian intestinal microbiota. Viruses 2019, 11, 760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, V.; Ali, A.; Fuentes, S.; Korpela, K.; Kazi, M.; Tate, J.; Parashar, U.; Wiersinga, W.J.; Giaquinto, C.; de Weerth, C.; et al. Rotavirus vaccine response correlates with the infant gut microbiota composition in Pakistan. Gut Microbes 2018, 9, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Harris, V.C.; Armah, G.; Fuentes, S.; Korpela, K.E.; Parashar, U.; Victor, J.C.; Tate, J.; de Weerth, C.; Giaquinto, C.; Wiersinga, W.J.; et al. Significant correlation between the infant gut microbiome and rotavirus vaccine response in rural Ghana. J. Infect. Dis. 2017, 215, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, R.; Chassaing, B.; Zhang, B.; Gewirtz, A.T. Antibiotic treatment suppresses rotavirus infection and enhances specific humoral immunity. J. Infect. Dis. 2014, 210, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Gozalbo-Rovira, R.; Rubio-Del-campo, A.; Santiso-Bellón, C.; Vila-Vicent, S.; Buesa, J.; Delgado, S.; Molinero, N.; Margolles, A.; Yebra, M.J.; Collado, M.C.; et al. Interaction of intestinal bacteria with human rotavirus during infection in children. Int. J. Mol. Sci. 2021, 22, 1010. [Google Scholar] [CrossRef]
- Goto, Y.; Uematsu, S.; Kiyono, H. Epithelial glycosylation in gut homeostasis and inflammation. Nat. Immunol. 2016, 17, 1244–1251. [Google Scholar] [CrossRef]
- Gozalbo-Rovira, R.; Ciges-Tomas, J.R.; Vila-Vicent, S.; Buesa, J.; Santiso-Bellón, C.; Monedero, V.; Yebra, M.J.; Marina, A.; Rodríguez-Díaz, J. Unraveling the role of the secretor antigen in human rotavirus attachment to histo-blood group antigens. PLoS Pathog. 2019, 15, e1007865. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ahmed, L.U.; Stuckert, M.R.; McGinnis, K.R.; Liu, Y.; Tan, M.; Huang, P.; Zhong, W.; Zhao, D.; Jiang, X.; et al. Molecular basis of P[II] major human rotavirus VP8* domain recognition of histo-blood group antigens. PLoS Pathog. 2020, 16, e1008386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramig, R.F. The effects of host age, virus dose, and virus strain on heterologous rotavirus infection of suckling mice. Microb. Pathog. 1988, 4, 189–202. [Google Scholar] [CrossRef]
- Arnold, M.; Patton, J.T.; McDonald, S.M. Culturing, storage, and quantification of rotaviruses. Curr. Protoc. Microbiol. 2009, 15. [Google Scholar] [CrossRef]
- Jothikumar, N.; Kang, G.; Hill, V.R. Broadly reactive TaqMan®assay for real-time RT-PCR detection of rotavirus in clinical and environmental samples. J. Virol. Methods 2009, 155, 126–131. [Google Scholar] [CrossRef]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubio-Del-Campo, A.; Gozalbo-Rovira, R.; Moya-Gonzálvez, E.M.; Alberola, J.; Rodríguez-Díaz, J.; Yebra, M.J. Infant gut microbiota modulation by human milk disaccharides in humanized microbiome mice. Gut Microbes 2021, 13, 1–20. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Rubio-Del-Campo, A.; Alcántara, C.; Collado, M.C.; Rodríguez-Díaz, J.; Yebra, M.J. Human milk and mucosa-associated disaccharides impact on cultured infant fecal microbiota. Sci. Rep. 2020, 10, 1–12. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Pérez-Ortín, R.; Vila-Vicent, S.; Carmona-Vicente, N.; Santiso-Bellón, C.; Rodríguez-Díaz, J.; Buesa, J. Histo-blood group antigens in children with symptomatic rotavirus infection. Viruses 2019, 11, 339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witjes, V.M.; Boleij, A.; Halffman, W. Reducing versus embracing variation as strategies for reproducibility: The microbiome of laboratory mice. Animals 2020, 10, 2415. [Google Scholar] [CrossRef]
- Schnepf, D.; Hernandez, P.; Mahlakõiv, T.; Crotta, S.; Sullender, M.E.; Peterson, S.T.; Ohnemus, A.; Michiels, C.; Gentle, I.; Dumoutier, L.; et al. Rotavirus susceptibility of antibiotic-treated mice ascribed to diminished expression of interleukin-22 2. bioRxiv 2021. [Google Scholar] [CrossRef]
- Li, N.; Ma, W.T.; Pang, M.; Fan, Q.L.; Hua, J.L. The commensal microbiota and viral infection: A comprehensive review. Front. Immunol. 2019, 10, 1551. [Google Scholar] [CrossRef]
- Abt, M.C.; Osborne, L.C.; Monticelli, L.A.; Doering, T.A.; Alenghat, T.; Sonnenberg, G.F.; Paley, M.A.; Antenus, M.; Williams, K.L.; Erikson, J.; et al. Commensal Bacteria Calibrate the Activation Threshold of Innate Antiviral Immunity. Immunity 2012, 37, 158–170. [Google Scholar] [CrossRef] [Green Version]
- Stefan, K.L.; Kim, M.V.; Iwasaki, A.; Kasper, D.L. Commensal microbiota modulation of natural resistance to virus infection. Cell 2020, 183, 1312–1324. [Google Scholar] [CrossRef] [PubMed]
- Yaron, J.R.; Ambadapadi, S.; Zhang, L.; Chavan, R.N.; Tibbetts, S.A.; Keinan, S.; Varsani, A.; Maldonado, J.; Kraberger, S.; Tafoya, A.M.; et al. Immune protection is dependent on the gut microbiome in a lethal mouse gammaherpesviral infection. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antunes, K.H.; Fachi, J.L.; de Paula, R.; da Silva, E.F.; Pral, L.P.; dos Santos, A.Á.; Dias, G.B.M.; Vargas, J.E.; Puga, R.; Mayer, F.Q.; et al. Microbiota-derived acetate protects against respiratory syncytial virus infection through a GPR43-type 1 interferon response. Nat. Commun. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.-L.; Wang, G.; Xie, J.-Y.; Li, H.; Liu, W.; Zhu, S.J. The intestinal microbiome primes host innate immunity against enteric virus 1 systemic infection through type I interferon 2. Res. Sq. 2021, 12, e00366-21. [Google Scholar]
- Thackray, L.B.; Handley, S.A.; Gorman, M.J.; Poddar, S.; Bagadia, P.; Briseño, C.G.; Theisen, D.J.; Tan, Q.; Hykes, B.L.; Lin, H.; et al. Oral antibiotic treatment of mice exacerbates the disease severity of multiple flavivirus infections. Cell Rep. 2018, 22, 3440–3453. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, S.; Vlasova, A.N.; Fischer, D.; Kumar, A.; Chattha, K.S.; Rauf, A.; Shao, L.; Langel, S.N.; Rajashekara, G.; Saif, L.J. Differential Effects of Escherichia coli Nissle and Lactobacillus rhamnosus Strain GG on Human Rotavirus Binding, Infection, and B Cell Immunity. J. Immunol. 2016, 196, 1780–1789. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Xiang, Y.; Li, N.; Wang, B.; Ai, H.; Wang, X.; Huang, L.; Zheng, Y. Protective effects of Lactobacillus rhamnosus GG against human rotavirus-induced diarrhoea in a neonatal mouse model. Pathog. Dis. 2013, 67, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, B.; Snipes-Magaldi, L.; Dennehy, P.; Keyserling, H.; Holman, R.C.; Bresee, J.; Gentsch, J.; Glass, R.I. Cytokines as mediators for or effectors against rotavirus disease in children. Clin. Diagn. Lab. Immunol. 2003, 10, 995–1001. [Google Scholar] [CrossRef] [Green Version]
- Kadooka, Y.; Tominari, K.; Sakai, F.; Yasui, H. Prevention of rotavirus-induced diarrhea by preferential secretion of IgA in breast milk via maternal administration of lactobacillus gasseri SBT2055. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Azevedo, M.S.P.; Gonzalez, A.; Zhang, W.; Saif, L.J.; Li, G.; Yousef, A.; Yuan, L. Toll-like receptor and innate cytokine responses induced by lactobacilli colonization and human rotavirus infection in gnotobiotic pigs. Vet. Immunol. Immunopathol. 2009, 127, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, B.; Saito, M.; Sakakibara, Y.; Hayashi, Y.; Yanagisawa, M.; Iwamori, M. Characterization of three members of murine α1, 2-fucosyltransferases: Change in the expression of the Se gene in the intestine of mice after administration of microbes. Arch. Biochem. Biophys. 2001, 388, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Candelero-Rueda, R.A.; Saif, L.J.; Vlasova, A.N. Infection of porcine small intestinal enteroids with human and pig rotavirus A strains reveals contrasting roles for histo-blood group antigens and terminal sialic acids. PLoS Pathog. 2021, 17, e1009237. [Google Scholar] [CrossRef] [PubMed]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Gomes Neto, J.C.; Boekschoten, M.V.; Schwab, C.; Urich, T.; et al. Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum schaedleri, a Core Member of the Murine Gut Microbiota. mSystems 2017, 2, e00171-16. [Google Scholar] [CrossRef] [Green Version]
- Herp, S.; Brugiroux, S.; Garzetti, D.; Ring, D.; Jochum, L.M.; Beutler, M.; Eberl, C.; Hussain, S.; Walter, S.; Gerlach, R.G.; et al. Mucispirillum schaedleri Antagonizes Salmonella Virulence to Protect Mice against Colitis. Cell Host Microbe 2019, 25, 681–694. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Natividad, J.M.; Lamas, B.; Pham, H.P.; Michel, M.L.; Rainteau, D.; Bridonneau, C.; Da Costa, G.; Van Hylckama Vlieg, J.; Sovran, B.; Chamignon, C.; et al. Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunctions in mice. Nat. Commun. 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di, J.B.; Gai, Z.T. Protective efficacy of probiotics on the treatment of acute rotavirus diarrhea in children: An updated meta-analysis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 9675–9683. [Google Scholar] [PubMed]
- Thompson, A.; Van Moorlehem, E.; Aich, P. Probiotic-induced priming of innate immunity to protect against rotaviral infection. Probiotics Antimicrob. Proteins 2010, 2, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Chattha, K.S.; Kandasamy, S.; Liu, Z.; Esseili, M.; Shao, L.; Rajashekara, G.; Saif, L.J. Lactobacilli and bifidobacteria promote immune homeostasis by modulating innate immune responses to human rotavirus in neonatal gnotobiotic pigs. PLoS ONE 2013, 8, e76962. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zou, J.; Zhang, Z.; Zhao, X.; Noriega, J.; Zhang, B.; Zhao, C.; Ingle, H.; Bittinger, K.; Mattei, L.M.; et al. Segmented filamentous bacteria prevent and cure rotavirus infection. Cell 2019, 179, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Diaz, J.; Garcia-Mantrana, I.; Vila-Vicent, S.; Gozalbo-Rovira, R.; Buesa, J.; Monedero, V.; Collado, M.C. Relevance of secretor status genotype and microbiota composition in susceptibility to rotavirus and norovirus infections in humans. Sci. Rep. 2017, 7, 1–10. [Google Scholar]
- Kawahara, T.; Makizaki, Y.; Oikawa, Y.; Tanaka, Y.; Maeda, A.; Shimakawa, M.; Komoto, S.; Moriguchi, K.; Ohno, H.; Taniguchi, K. Oral administration of Bifidobacterium bifidum G9-1 alleviates rotavirus gastroenteritis through regulation of intestinal homeostasis by inducing mucosal protective factors. PLoS ONE 2017, 12, e0173979. [Google Scholar] [CrossRef] [Green Version]
- Salminen, S.; Nybom, S.; Meriluoto, J.; Collado, M.C.; Vesterlund, S.; El-Nezami, H. Interaction of probiotics and pathogens—Benefits to human health? Curr. Opin. Biotechnol. 2010, 21, 157–167. [Google Scholar] [CrossRef]
- Almand, E.A.; Moore, M.D.; Outlaw, J.; Jaykus, L.-A. Human norovirus binding to select bacteria representative of the human gut microbiota. PLoS ONE 2017, 12, e0173124. [Google Scholar] [CrossRef]
- Jones, M.K.; Watanabe, M.; Zhu, S.; Graves, C.L.; Keyes, L.R.; Grau, K.R.; Gonzalez-Hernandez, M.B.; Iovine, N.M.; Wobus, C.E.; Vinje, J.; et al. Enteric bacteria promote human and mouse norovirus infection of B cells. Science 2014, 346, 755–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuss, S.K.; Best, G.T.; Etheredge, C.A.; Pruijssers, A.J.; Frierson, J.M.; Hooper, L.V.; Dermody, T.S.; Pfeiffer, J.K. Intestinal microbiota promote enteric virus replication and systemic pathogenesis. Science 2011, 334, 249–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Breiman, A.; le Pendu, J.; Uyttendaele, M. Binding to histo-blood group antigen-expressing bacteria protects human norovirus from acute heat stress. Front. Microbiol. 2015, 6, 659. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gozalbo-Rovira, R.; Santiso-Bellón, C.; Buesa, J.; Rubio-del-Campo, A.; Vila-Vicent, S.; Muñoz, C.; Yebra, M.J.; Monedero, V.; Rodríguez-Díaz, J. Microbiota Depletion Promotes Human Rotavirus Replication in an Adult Mouse Model. Biomedicines 2021, 9, 846. https://doi.org/10.3390/biomedicines9070846
Gozalbo-Rovira R, Santiso-Bellón C, Buesa J, Rubio-del-Campo A, Vila-Vicent S, Muñoz C, Yebra MJ, Monedero V, Rodríguez-Díaz J. Microbiota Depletion Promotes Human Rotavirus Replication in an Adult Mouse Model. Biomedicines. 2021; 9(7):846. https://doi.org/10.3390/biomedicines9070846
Chicago/Turabian StyleGozalbo-Rovira, Roberto, Cristina Santiso-Bellón, Javier Buesa, Antonio Rubio-del-Campo, Susana Vila-Vicent, Carlos Muñoz, María J. Yebra, Vicente Monedero, and Jesús Rodríguez-Díaz. 2021. "Microbiota Depletion Promotes Human Rotavirus Replication in an Adult Mouse Model" Biomedicines 9, no. 7: 846. https://doi.org/10.3390/biomedicines9070846
APA StyleGozalbo-Rovira, R., Santiso-Bellón, C., Buesa, J., Rubio-del-Campo, A., Vila-Vicent, S., Muñoz, C., Yebra, M. J., Monedero, V., & Rodríguez-Díaz, J. (2021). Microbiota Depletion Promotes Human Rotavirus Replication in an Adult Mouse Model. Biomedicines, 9(7), 846. https://doi.org/10.3390/biomedicines9070846