
Oviductal Telocytes in Patients with Uterine Myoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Ethical Approval
2.3. Tissue Processing
2.4. Routine Histology
2.5. Immunofluorescence
2.6. Microscopic Examination
3. Results
3.1. General Structure of the Human Oviduct
3.2. IHC Analysis of Telocytes in the Fallopian Tubes
3.3. IHC Analysis of Nerve Fibers and Its Interactions with Oviduct Telocytes
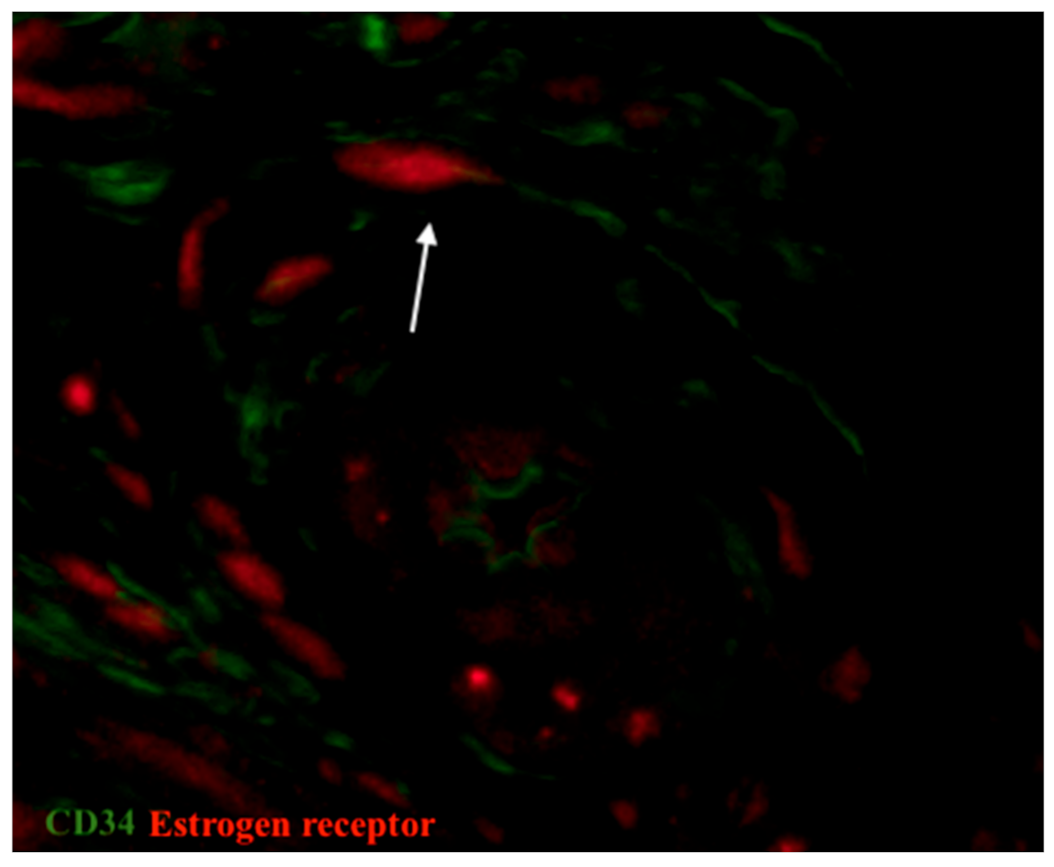
3.4. IHC Analysis of Estrogen and Progesterone Receptor Expression
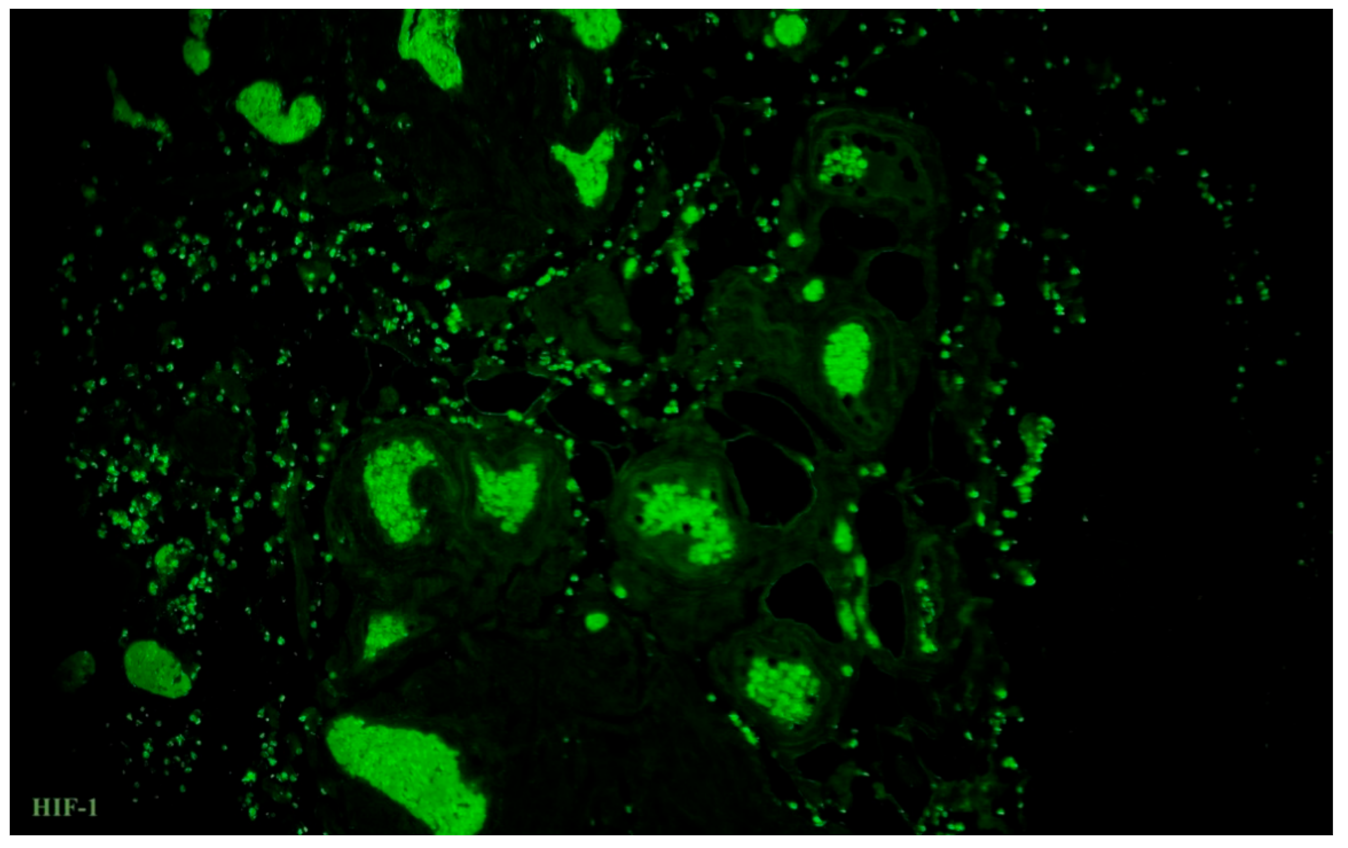
3.5. Immunolabelling of Telocytes with Makers of Hypoxia and Ion Channels
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lyons, R.; Saridogan, E.; Djahanbakhch, O. The reproductive significance of human Fallopian tube cilia. Hum. Reprod. Updat. 2006, 12, 363–372. [Google Scholar] [CrossRef]
- Abdelhamed, Z.A.; Ryan, T.A.; Fuller, M.; Coulson-Gilmer, C.; Abdelmottaleb, D.I.; Wang, T.-L.; Kaun, J.-C.; Wang, P.; Hutson, R.; Wilkinson, N.; et al. Characterization of Primary Cilia in Normal Fallopian Tube Epithelium and Serous Tubal Intraepithelial Carcinoma. Int. J. Gynecol. Cancer 2018, 28, 1535–1544. [Google Scholar] [CrossRef] [Green Version]
- Ferenczy, A.; Richart, R.M.; Agate, F.J.; Purkerson, M.L.; Dempsey, E.W. Scanning Electron Microscopy of the Human Fallopian Tube. Science 1972, 175, 783–784. [Google Scholar] [CrossRef]
- Patek, E. The epithelium of the human Fallopian tube. A surface ultrastructural and cytochemical study. Acta Obstet. Gynecol. Scand. Suppl. 1974, 31, 1–28. [Google Scholar] [PubMed]
- Costache, A.; Purcarea, V.L.; Cergan, R.; Dumitru, M.; Briceag, I.; Sajin, M.; Ispas, A.T. Fallopian tubes–literature review of anatomy and etiology in female infertility. J. Med. Life 2015, 8, 129–131. [Google Scholar]
- Zhu, M.; Iwano, T.; Takeda, S. Estrogen and EGFR Pathways Regulate Notch Signaling in Opposing Directions for Multi-Ciliogenesis in the Fallopian Tube. Cells 2019, 8, 933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleksandrovych, V.; Sajewicz, M.; Walocha, J.A.; Gil, K. Tubal telocytes: Factor infertility reason? Folia Med Crac. 2016, 56, 17–23. [Google Scholar] [PubMed]
- Almasry, S.M.; Elfayomy, A.K.; El-Sherbiny, M.H. Regeneration of the Fallopian Tube Mucosa Using Bone Marrow Mesenchymal Stem Cell Transplantation After Induced Chemical Injury in a Rat Model. Reprod. Sci. 2017, 25, 773–781. [Google Scholar] [CrossRef]
- Yamazaki, K.; Eyden, B.P. Ultrastructural and immunohistochemical studies of stromal cells in lamina propria of human fallopian tube ampullar mucosa: The recognition of ‘CD34 positive reticular network’ and its putative function for immune surveillance. J. Submicrosc. Cytol. Pathol. 1996, 28, 325–337. [Google Scholar]
- Ardighieri, L.; Lonardi, S.; Moratto, D.; Facchetti, F.; Shih, I.-M.; Vermi, W.; Kurman, R.J. Characterization of the Immune Cell Repertoire in the Normal Fallopian Tube. Int. J. Gynecol. Pathol. 2014, 33, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Chen, J.; Sun, Y.; Huang, X.; Xu, H.; Zhang, X. Loss of nerve fibers in the oviduct isthmus in women with hydrosalpinx. Acta Histochem. 2013, 115, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Herrera, G.G.B.; Lierz, S.L.; A Harris, E.; Donoghue, L.J.; Hewitt, S.C.; Rodriguez, K.F.; Jefferson, W.N.; Lydon, J.P.; DeMayo, F.J.; Williams, C.J.; et al. Oviductal Retention of Embryos in Female Mice Lacking Estrogen Receptor α in the Isthmus and the Uterus. Endocrinology 2019, 161. [Google Scholar] [CrossRef]
- Shao, R.; Feng, Y.; Zou, S.; Weijdegård, B.; Wu, G.; Brännström, M.; Billig, H. The role of estrogen in the pathophysiology of tubal ectopic pregnancy. Am. J. Transl. Res. 2012, 4, 269–278. [Google Scholar]
- Li, S.; O’Neill, S.R.S.; Zhang, Y.; Holtzman, M.J.; Takemaru, K.; Korach, K.S.; Winuthayanon, W. Estrogen receptor α is required for oviductal transport of embryos. FASEB J. 2016, 31, 1595–1607. [Google Scholar] [CrossRef] [Green Version]
- Halbert, S.A.; Becker, D.R.; Szal, S.E. Ovum Transport in the Rat Oviductal Ampulla in the Absence of Muscle Contractility1. Biol. Reprod. 1989, 40, 1131–1136. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Lemarié, A.; Zou, S.; Li, X.; Cui, P.; Weijdegård, B.; Wu, G.; Shao, R.; Billig, H.; Feng, Y. The Regulation of Nitric Oxide Synthase Isoform Expression in Mouse and Human Fallopian Tubes: Potential Insights for Ectopic Pregnancy. Int. J. Mol. Sci. 2014, 16, 49–67. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Li, D.; Johns, R.A. Expression of endothelial nitric oxide synthase in ciliated epithelia of rats. J. Histochem. Cytochem. 2003, 51, 81–87. [Google Scholar] [CrossRef] [Green Version]
- Popescu, L.M.; Faussone-Pellegrini, M.-S. TELOCYTES—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [Green Version]
- Cretoiu, S.M.; Popescu, L.M. Telocytes revisited. Biomol. Concepts 2014, 5, 353–369. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovych, V.; Pasternak, A.; Basta, P.; Sajewicz, M.; A Walocha, J.; Gil, K. Telocytes: Facts, speculations and myths (Review article). Folia Med. Crac. 2017, 57, 5–22. [Google Scholar] [PubMed]
- Cretoiu, S.M.; Cretoiu, D.; Marin, A.B.; Radu, B.M.; Popescu, L.M. Telocytes: Ultrastructural, immunohistochemical and electrophysiological characteristics in human myometrium. Reproduction 2013, 145, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovych, V.; Walocha, J.A.; Gil, K. Telocytes in female reproductive system (human and animal). J. Cell. Mol. Med. 2016, 20, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovych, V.; Pasternak, A.; Gil, K. Telocytes in the architecture of uterine fibroids. Folia Med. Crac. 2019, 59, 33–44. [Google Scholar]
- Popescu, L.; Ciontea, S.M.; Cretoiu, D.; Hinescu, M.; Radu, E.; Ionescu, N.; Ceausu, M.; Gherghiceanu, M.; Braga, R.I.; Vasilescu, F.; et al. Novel type of interstitial cell (Cajal-like) in human fallopian tube. J. Cell. Mol. Med. 2005, 9, 479–523. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, S.M. Immunohistochemistry of Telocytes in the Uterus and Fallopian Tubes. Telocytes 2016, 913, 335–357. [Google Scholar] [CrossRef]
- Cretoiu, S.M.; Cretoiu, D.; Suciu, L.; Popescu, L.M. Interstitial Cajal-like cells of human Fallopian tube express estrogen and progesterone receptors. J. Mol. Histol. 2009, 40, 387–394. [Google Scholar] [CrossRef]
- Roatesi, I.; Radu, B.M.; Cretoiu, D.; Cretoiu, S.M. Uterine Telocytes: A Review of Current Knowledge1. Biol. Reprod. 2015, 93, 10. [Google Scholar] [CrossRef]
- Yang, X.-J.; Wei, W.; Zhao, J.; Zheng, F.-Y. Inhibitory effects of methotrexate on spontaneous motility and Cajal-like type of tubal interstitial cells in rabbit oviduct. Fertil. Steril. 2012, 98, 215–221. [Google Scholar] [CrossRef]
- Abdelhafeez, H.; Soliman, S. New Description of Telocyte Sheaths in the Bovine Uterine Tube: An Immunohistochemical and Scanning Microscopic Study. Cells Tissues Organs 2016, 203, 295–315. [Google Scholar] [CrossRef]
- Richter, M.; Kostin, S. The failing human heart is characterized by decreased numbers of telocytes as result of apoptosis and altered extracellular matrix composition. J. Cell. Mol. Med. 2015, 19, 2597–2606. [Google Scholar] [CrossRef]
- Matyja, A.; Gil, K.; Pasternak, A.; Sztefko, K.; Gajda, M.; Tomaszewski, K.; Matyja, M.; Walocha, J.A.; Kulig, J.; Thor, P. Telocytes: New insight into the pathogenesis of gallstone disease. J. Cell. Mol. Med. 2013, 17, 734–742. [Google Scholar] [CrossRef] [PubMed]
- Milia, A.F.; Ruffo, M.; Manetti, M.; Rosa, I.; Conte, D.; Fazi, M.; Messerini, L.; Ibba-Manneschi, L. Telocytes in Crohn’s disease. J. Cell. Mol. Med. 2013, 17, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovych, V.; Kurnik-Łucka, M.; Bereza, T.; Białas, M.; Pasternak, A.; Cretoiu, D.; Walocha, J.A.; Gil, K. The Autonomic Innervation and Uterine Telocyte Interplay in Leiomyoma Formation. Cell Transplant. 2019, 28, 619–629. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Gil, A.; Wrona, A. Sex steroid hormone receptors of telocytes—Potential key role in leiomyoma de-velopment. Folia Med. Crac. 2020, 60, 81–95. [Google Scholar]
- Mokhtar, D.M.; Abdelhafeez, H.; Abou-Elmagd, A.; Hassan, A.H.S. Melatonin administration induced reactivation in the seminal gland of the soay rams during non-breeding season: An ultrastructural and morphometrical study. J. Morphol. 2015, 277, 231–243. [Google Scholar] [CrossRef] [PubMed]
- Ibba-Manneschi, L.; Rosa, I.; Manetti, M. Telocyte implications in human pathology: An overview. Semin. Cell Dev. Biol. 2016, 55, 62–69. [Google Scholar] [CrossRef]
- Aida-Yasuoka, K.; Peoples, C.; Yasuoka, H.; Hershberger, P.; Thiel, K.; A Cauley, J.; A MedsgerJr, T.; A Feghali-Bostwick, C. Estradiol promotes the development of a fibrotic phenotype and is increased in the serum of patients with systemic sclerosis. Arthritis Res. Ther. 2013, 15, R10. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, A.; Sakuma, K.; Shimamoto, C.; Ito, S.; Daikoku, E.; Ohmichi, M.; Ushiroyama, T.; Ueki, M.; Kuwabara, H.; Mori, H.; et al. Ciliary beat frequency controlled by oestradiol and progesterone during ovarian cycle in guinea-pig Fallopian tube. Exp. Physiol. 2010, 95, 819–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janas, P.; Kucybała, I.; Radoń-Pokracka, M.; Huras, H. Telocytes in the female reproductive system: An overview of up-to-date knowledge. Adv. Clin. Exp. Med. 2018, 27, 559–565. [Google Scholar] [CrossRef]
- Abdel-Maksoud, F.M.; Abdelhafeez, H.; Soliman, S. Morphological changes of telocytes in camel efferent ductules in response to seasonal variations during the reproductive cycle. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Marini, M.; Rosa, I.; Guasti, D.; Gacci, M.; Sgambati, E.; Ibba-Manneschi, L.; Manetti, M. Reappraising the microscopic anatomy of human testis: Identification of telocyte networks in the peritubular and intertubular stromal space. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Pawlicki, P.; Hejmej, A.; Milon, A.; Lustofin, K.; Płachno, B.J.; Tworzydlo, W.; Gorowska-Wojtowicz, E.; Pawlicka, B.; Kotula-Balak, M.; Bilinska, B. Telocytes in the mouse testicular interstitium: Implications of G-protein-coupled estrogen receptor (GPER) and estrogen-related receptor (ERR) in the regulation of mouse testicular interstitial cells. Protoplasma 2018, 256, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.; Zhu, Q.; Yan, M.; Li, C.; Yuan, J.; Qin, G.; Zhang, J. Levonorgestrel decreases cilia beat frequency of human fallopian tubes and rat oviducts without changing morphological structure. Clin. Exp. Pharmacol. Physiol. 2015, 42, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Salama, N.M. Immunohistochemical characterization of telocytes in ratuterus in different reproductive states. Egypt. J. Histol. 2013, 36, 185–194. [Google Scholar] [CrossRef]
- Jankovic, S.; Protic, B.; Jankovic, S. Contractile effect of acetylcholine on isolated isthmic segment of fallopian tubes. Methods Find. Exp. Clin. Pharmacol. 2004, 26, 87. [Google Scholar] [CrossRef]
- Šteffl, M.; Schweiger, M.; Wessler, I.; Kunz, L.; Mayerhofer, A.; Amselgruber, W.M. Non-neuronal acetylcholine and choline acetyltransferase in oviductal epithelial cells of cyclic and pregnant pigs. Anat. Embryol. 2006, 211, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Noreikat, K.; Wolff, M.; Kummer, W.; Kölle, S. Ciliary Activity in the Oviduct of Cycling, Pregnant, and Muscarinic Receptor Knockout Mice1. Biol. Reprod. 2012, 86, 120. [Google Scholar] [CrossRef] [PubMed]
- Wolff, M.; Noreikat, K.; Ibañez-Tallon, I.; Lips, K.S.; Kölle, S.; Kummer, W. Cholinergic receptors in the murine oviduct: Inventory and coupling to intracellular calcium concentration. Life Sci. 2012, 91, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, S.T.; Svalø, J.; Nielsen, K.; Larsen, T.; Jørgensen, J.C.; Bouchelouche, P. Immunolocalization and expression of small-conductance calcium-activated potassium channels in human myometrium. J. Cell. Mol. Med. 2012, 16, 3001–3008. [Google Scholar] [CrossRef]
- Keating, N.; Quinlan, L.R. Small conductance potassium channels drive ATP-activated chloride secretion in the oviduct. Am. J. Physiol. Physiol. 2012, 302, C100–C109. [Google Scholar] [CrossRef]
- Jung, C.; Fernández-Dueñas, V.; Plata, C.; Garcia-Elias, A.; Ciruela, F.; Fernández-Fernández, J.M.; Valverde, M.A. Functional coupling of GABAA/B receptors and the channel TRPV4 mediates rapid progesterone signaling in the oviduct. Sci. Signal. 2018, 11, eaam6558. [Google Scholar] [CrossRef] [Green Version]
- Doerner, J.F.; Delling, M.; E Clapham, D. Ion channels and calcium signaling in motile cilia. eLife 2015, 4, e11066. [Google Scholar] [CrossRef]
- Chi, C.; Jiang, X.; Su, L.; Shen, Z.; Yang, X. In vitro morphology, viability and cytokine secretion of uterine telocyte-activated mouse peritoneal macrophages. J. Cell. Mol. Med. 2015, 19, 2741–2750. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.-J.; Cretoiu, D.; Shen, Z.-J.; Yang, X.-J. An in vitro investigation of telocytes-educated macrophages: Morphology, heterocellular junctions, apoptosis and invasion analysis. J. Transl. Med. 2018, 16, 1–12. [Google Scholar] [CrossRef]
- Yang, X.; Yang, J.; Liu, Z.; Yang, G.; Shen, Z. Telocytes damage in endometriosis-affected rat oviduct and potential impact on fertility. J. Cell. Mol. Med. 2014, 19, 452–462. [Google Scholar] [CrossRef]
- Popescu, L.M.; Ciontea, S.M.; Cretoiu, D.; Cretoiu, S.M. Interstitial Cajal-Like Cells in Human Uterus and Fallopian Tube. Ann. N. Y. Acad. Sci. 2007, 1101, 139–165. [Google Scholar] [CrossRef] [PubMed]
- Albulescu, R.; Tanase, C.; Codrici, E.; Popescu, D.I.; Cretoiu, S.M.; Popescu, L.M. The secretome of myocardial telocytes modulates the activity of cardiac stem cells. J. Cell. Mol. Med. 2015, 19, 1783–1794. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Nasu, K.; Yuge, A.; Kawano, Y.; Yoshimatsu, J.; Narahara, H. Interleukin-13 stimulates the secretion of vascular endothelial growth factor and soluble fms-like tyrosine kinase-1 by human oviductal epithelial cells. Eur. J. Obstet. Gynecol. Reprod. Biol. 2007, 133, 208–212. [Google Scholar] [CrossRef]
- Itoh, H.; Nasu, K.; Matsumoto, H.; Kawano, Y.; Yoshimatsu, J.; Narahara, H. Hypoxia regulates vascular endothelial growth factor and soluble fms-like tyrosine kinase-1 secretion by human oviductal epithelial cells and stromal fibroblasts. Fertil. Steril. 2006, 85, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Madu, C.O.; Lu, A.; Lu, Y. HIF-1α Promotes A Hypoxia-Independent Cell Migration. Open Biol. J. 2010, 3, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Laganà, A.S.; Vergara, D.; Favilli, A.; La Rosa, V.L.; Tinelli, A.; Gerli, S.; Noventa, M.; Vitagliano, A.; Triolo, O.; Rapisarda, A.M.C.; et al. Epigenetic and genetic landscape of uterine leiomyomas: A current view over a common gynecological disease. Arch. Gynecol. Obstet. 2017, 296, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Mas, A.; Diamond, M.; Al-Hendy, A.; Yang, Q. The Mechanism and Function of Epigenetics in Uterine Leiomyoma Development. Reprod. Sci. 2015, 23, 163–175. [Google Scholar] [CrossRef] [PubMed]
- González-Brusi, L.; Algarra, B.; Moros-Nicolás, C.; Izquierdo-Rico, M.J.; Avilés, M.; Jiménez-Movilla, M. A Comparative View on the Oviductal Environment during the Periconception Period. Biomolecules 2020, 10, 1690. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area of the Human Fallopian Tube | Density of TCs (%) |
|---|---|
| The border epithelium/lamina propria, a ‘belt’ 10 µm thick underneath the basement membrane of the endosalpinx epithelium | 18 ± 2 |
| The subepithelial portion of lamina propria (~20 µm thick) | 11.7 ± 0.9 |
| Area, containing the whole lamina propria thickness | 9 |
| Tunica muscularis | 7.8 ± 1.2 |
| Remaining zone beneath serosa | not assessed |
| Antibody | Catalog Number and Company | Dilution |
|---|---|---|
| Primary Antibodies | ||
| Polyclonal rabbit anti-PGP 9.5 | Z5116, Dako, Glostrup, Denmark | 1:100 |
| Polyclonal mouse anti-NOS | sc-7271, Santa Cruz, Dallas, Texas, USA | 1:100 |
| Monoclonal mouse anti-ChAT | sc-55557, Santa Cruz, Dallas, Texas, USA | 1:100 |
| Monoclonal mouse anti-CD31 | JC70A, Dako, Glostrup, Denmark | 1:100 |
| Polyclonal rabbit anti-c-kit | A4502, Dako, Glostrup, Denmark | 1:100 |
| Monoclonal mouse anti-CD34 | M7165, Dako, Glostrup, Denmark | 1:100 |
| Polyclonal goat anti-PDGFR alpha | AF-307-NA, R&D Systems, Minneapolis, Minnesota, USA | 1:100 |
| Monoclonal mouse anti-tryptase | M7052, Dako, Glostrup, Denmark | 1:100 |
| Monoclonal mouse anti-vimentin | Clone V9, Dako, Glostrup, Denmark | 1:50 |
| Monoclonal mouse progesterone-receptor | Clone PgR636, Dako, Glostrup, Denmark | 1:100 |
| Monoclonal mouse estrogen receptor | NCL-L-ER-6F11, Leica Biosystems, Newcastle upon Tyne, UK | 1:50 |
| Polyclonal goat anti-VEGF R1/Flt-1 | AF321, R&D Systems, Minneapolis, Minnesota, USA | 5 μg/mL |
| Monoclonal mouse anti-HIF-1 | ab16066, Abcam, Cambridge, UK | 1:100 |
| Monoclonal rabbit anti-CD34 | ab81289, Abcam, Cambridge, UK | 1:200 |
| Polyclonal rabbit anti-KCNN3 (SK3) | APC-025, Alomone Labs, Jerusalem, Israel | 1:800 |
| Secondary Antibodies | ||
| Alexa Fluor 594 Goat Anti-Mouse | 115-585-146, Jackson ImmunoResearch, Ely, UK | 1:400 |
| Alexa Fluor 488 Goat Anti-Mouse | 115-545-146, Jackson ImmunoResearch, Ely, UK | 1:400 |
| Alexa Fluor 594 Goat Anti-Rabbit | 111-585-144, Jackson ImmunoResearch, Ely, UK | 1:400 |
| Alexa Fluor 488 Rabbit Anti-Mouse | 315-545-045, Jackson ImmunoResearch, Ely, UK | 1:400 |
| Alexa Fluor 488 Goat Anti-Rabbit | 111-545-144, Jackson ImmunoResearch, Ely, UK | 1:400 |
| Alexa Fluor 594 Donkey Anti-Goat | 705-585-003, Jackson ImmunoResearch, Ely, UK | 1:400 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleksandrovych, V.; Wrona, A.; Bereza, T.; Pityński, K.; Gil, K. Oviductal Telocytes in Patients with Uterine Myoma. Biomedicines 2021, 9, 1060. https://doi.org/10.3390/biomedicines9081060
Aleksandrovych V, Wrona A, Bereza T, Pityński K, Gil K. Oviductal Telocytes in Patients with Uterine Myoma. Biomedicines. 2021; 9(8):1060. https://doi.org/10.3390/biomedicines9081060
Chicago/Turabian StyleAleksandrovych, Veronika, Anna Wrona, Tomasz Bereza, Kazimierz Pityński, and Krzysztof Gil. 2021. "Oviductal Telocytes in Patients with Uterine Myoma" Biomedicines 9, no. 8: 1060. https://doi.org/10.3390/biomedicines9081060
APA StyleAleksandrovych, V., Wrona, A., Bereza, T., Pityński, K., & Gil, K. (2021). Oviductal Telocytes in Patients with Uterine Myoma. Biomedicines, 9(8), 1060. https://doi.org/10.3390/biomedicines9081060