
Anti-Inflammatory Properties of Diet: Role in Healthy Aging

 , , and
, , and 
Abstract
:1. Aging and Inflammation: Inflammaging
2. Diet and Inflammation
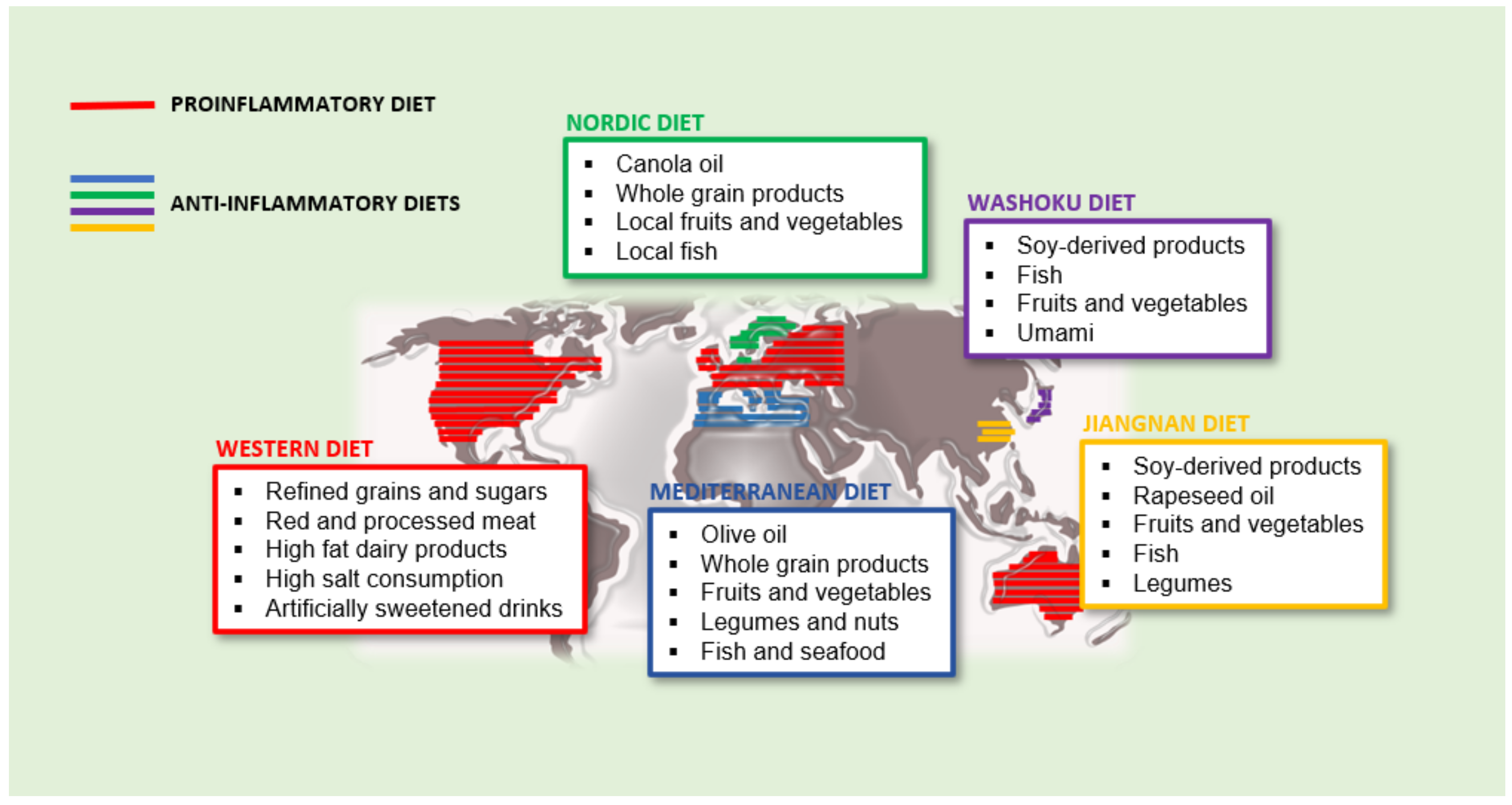
2.1. The Western Diet: An Example of Proinflammatory Dietary Pattern
2.2. Anti-Inflammatory Diets
2.2.1. Mediterranean Diet
2.2.2. Nordic Diet (ND)
2.2.3. Asian Diet
Japanese Traditional Diet: Washoku
Chinese Traditional Diet: Jiangnan Diet
3. The Anti-Inflammatory Compounds Present in the Diet: Classification and Mechanism of Action
3.1. Classification
3.2. Mechanism of Action
3.2.1. ω-3 Polyunsaturated Fatty Acids (ω-3 LC-PUFAs)
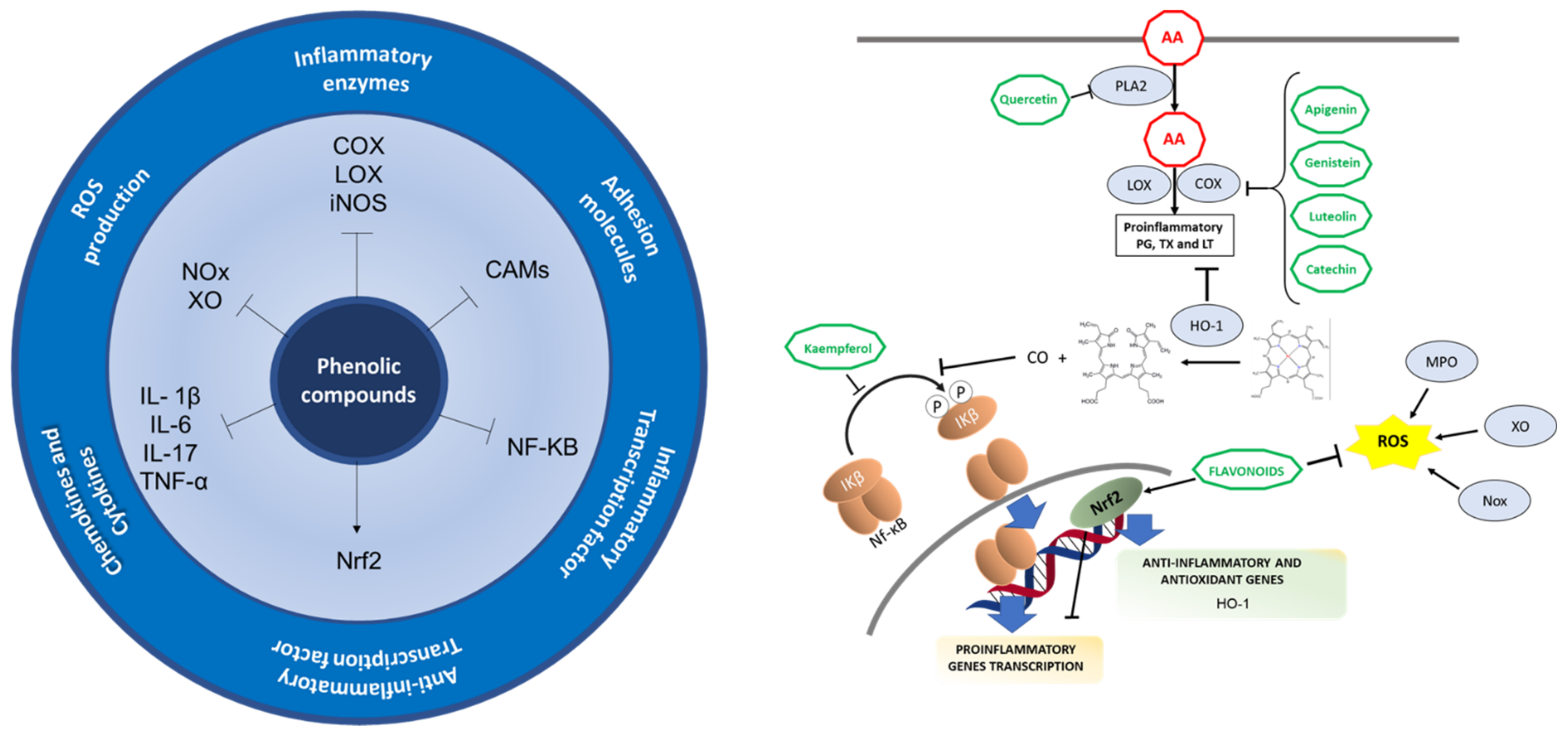
3.2.2. Phenolic Compounds and Polyphenols
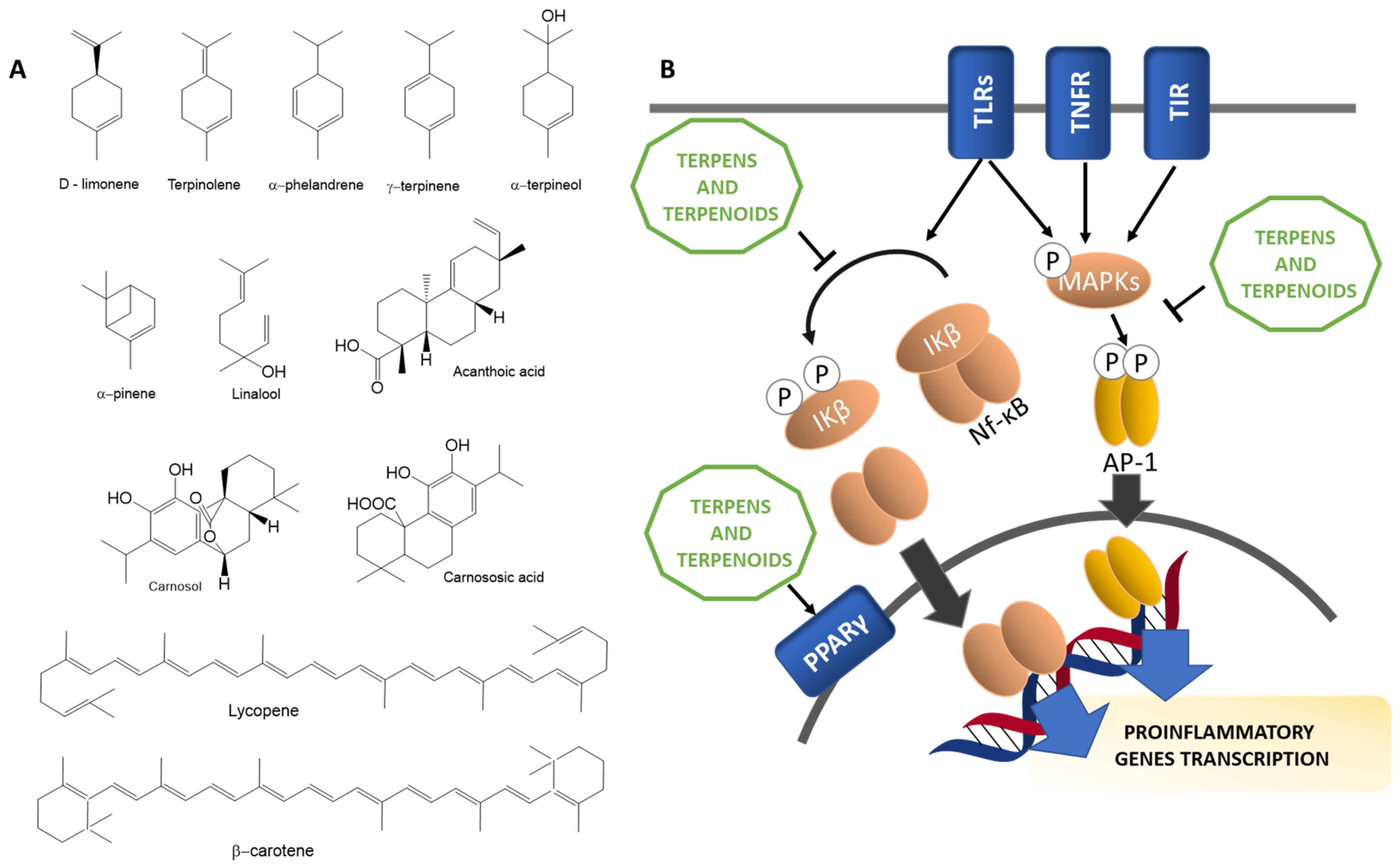
3.2.3. Terpenes and Terpenoids
4. Bioavailability
5. An Anti-Inflammatory Dietary Pattern: What and How Much to Eat
6. Inflammation, Oxidative Stress, and Hormesis
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | arachidonic acid |
| AA/EPA | arachidonic/eicosapentaenoic acid ratio |
| AGEs | advanced glycation end products |
| ALS | amyotrophic lateral sclerosis |
| CO | carbon monoxide |
| COX | cyclooxygenases |
| CRP | C-reactive protein |
| CVD | cardiovascular disease |
| DHA | docosahezaenoic acid |
| DII | dietary inflammation index |
| EPA | eicosapentaenoic acid |
| GI | glycemic index |
| GL | glycemic load |
| GPR120 | G-protein coupled receptor 120 |
| ICAM-1 | Intercellular Adhesion Molecule 1 |
| IKKs | IκB Kinases |
| IKKβ | IκB Kinase beta |
| IL | interleukins |
| iNOS | nitric oxide synthase |
| LOX | lipoxygenases |
| LPS | lipopolysaccharides |
| MAPK | mitogen-activated protein kinase |
| MaRs | maresins |
| MD | Mediterranean Diet |
| NCD | noncommunicable diseases |
| ND | Nordic Diet |
| NO | nitric oxide |
| NF-κB | nuclear factor kappa b |
| PDs | protectins |
| PG | prostaglandins |
| PLA2 | phospholipase A2 |
| PPAR | peroxisome proliferator-activated receptor |
| PRRs | Pattern Recognition Receptors |
| PUFAs | polyunsaturated fatty acids |
| ROS | radical oxidative species |
| RvD1 | resolvin D1 |
| RvE1 | resolvin E1 |
| TAB1 | TAK-1 binding proteins |
| TAK1 | transforming growth factor-β-activated kinase 1 |
| TIR | Toll/IL-1 receptor |
| TNF-α | necrosis factor tumor alpha |
| TLRs | including toll-like receptor |
| TXs | thromboxanes |
| VCAM | Vascular Adhesion Molecule |
References
- Jin, K. Modern Biological Theories of Aging. Aging Dis. 2010, 1, 72–74. [Google Scholar]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Inflammaging: Disturbed interplay between autophagy and inflammasomes. Aging 2012, 4, 166–175. [Google Scholar] [CrossRef] [Green Version]
- Green, D.R.; Galluzzi, L.; Kroemer, G. Mitochondria and the autophagy-inflammation-cell death axis in organismal aging. Science 2011, 333, 1109–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galland, L. Diet and inflammation. Nutr. Clin. Pract. 2010, 25, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Albers, R.; Antoine, J.-M.; Blum, S.; Bourdet-Sicard, R.; Ferns, G.A.; Folkerts, G.; Friedmann, P.S.; Frost, G.S.; Guarner, F.; et al. Inflammatory disease processes and interactions with nutrition. Br. J. Nutr. 2009, 101 (Suppl. 1), S1–S45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra-Majem, L.; Román-Viñas, B.; Sanchez-Villegas, A.; Guasch-Ferré, M.; Corella, D.; La Vecchia, C. Benefits of the Mediterranean diet: Epidemiological and molecular aspects. Mol. Asp. Med. 2019, 67, 1–55. [Google Scholar] [CrossRef]
- Shivappa, N.; Steck, S.E.; Hurley, T.G.; Hussey, J.R.; Hébert, J.R. Designing and developing a literature-derived, population-based dietary inflammatory index. Public Health Nutr. 2014, 17, 1689–1696. [Google Scholar] [CrossRef] [Green Version]
- Bloomfield, H.E.; Kane, R.; Koeller, E.; Greer, N.; MacDonald, R.; Wilt, T. Benefits and Harms of the Mediterranean Diet Compared to Other Diets; VA Evidence-based Synthesis Program Reports; Department of Veterans Affairs (US): Washington, DC, USA, 2015.
- Iikuni, N.; Lam, Q.L.K.; Lu, L.; Matarese, G.; La Cava, A. Leptin and Inflammation. Curr. Immunol. Rev. 2008, 4, 70–79. [Google Scholar] [CrossRef]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef]
- Karaskova, E.; Velganova-Veghova, M.; Geryk, M.; Foltenova, H.; Kucerova, V.; Karasek, D. Role of Adipose Tissue in Inflammatory Bowel Disease. Int. J. Mol. Sci. 2021, 22, 4266. [Google Scholar] [CrossRef] [PubMed]
- Bettiga, A.; Fiorio, F.; Di Marco, F.; Trevisani, F.; Romani, A.; Porrini, E.; Salonia, A.; Montorsi, F.; Vago, R. The Modern Western Diet Rich in Advanced Glycation End-Products (AGEs): An Overview of Its Impact on Obesity and Early Progression of Renal Pathology. Nutrients 2019, 11, 1748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uribarri, J.; del Castillo, M.D.; de la Maza, M.P.; Filip, R.; Gugliucci, A.; Luevano-Contreras, C.; Macías-Cervantes, M.H.; Markowicz Bastos, D.H.; Medrano, A.; Menini, T.; et al. Dietary advanced glycation end products and their role in health and disease. Adv. Nutr. 2015, 6, 461–473. [Google Scholar] [CrossRef]
- Davis, K.E.; Prasad, C.; Vijayagopal, P.; Juma, S.; Imrhan, V. Advanced Glycation End Products, Inflammation, and Chronic Metabolic Diseases: Links in a Chain? Crit. Rev. Food Sci. Nutr. 2016, 56, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Yacoub, R.; Nugent, M.; Cai, W.; Nadkarni, G.N.; Chaves, L.D.; Abyad, S.; Honan, A.M.; Thomas, S.A.; Zheng, W.; Valiyaparambil, S.A.; et al. Advanced glycation end products dietary restriction effects on bacterial gut microbiota in peritoneal dialysis patients; A randomized open label controlled trial. PLoS ONE 2017, 12, e0184789. [Google Scholar] [CrossRef] [Green Version]
- Di Daniele, N.; Noce, A.; Vidiri, M.F.; Moriconi, E.; Marrone, G.; Annicchiarico-Petruzzelli, M.; D’Urso, G.; Tesauro, M.; Rovella, V.; De Lorenzo, A. Impact of Mediterranean diet on metabolic syndrome, cancer and longevity. Oncotarget 2017, 8, 8947–8979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alessandro, A.; De Pergola, G.; Silvestris, F. Mediterranean Diet and cancer risk: An open issue. Int. J. Food Sci. Nutr. 2016, 67, 593–605. [Google Scholar] [CrossRef]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef]
- Buettner, D.; Skemp, S. Blue Zones: Lessons from the World’s Longest Lived. Am. J. Lifestyle Med. 2016, 10, 318–321. [Google Scholar] [CrossRef]
- Keys, A.; Menotti, A.; Karvonen, M.J.; Aravanis, C.; Blackburn, H.; Buzina, R.; Djordjevic, B.S.; Dontas, A.S.; Fidanza, F.; Keys, M.H. The diet and 15-year death rate in the seven countries study. Am. J. Epidemiol. 1986, 124, 903–915. [Google Scholar] [CrossRef]
- Martínez-González, M.A.; Salas-Salvadó, J.; Estruch, R.; Corella, D.; Fitó, M.; Ros, E. Predimed Investigators Benefits of the Mediterranean Diet: Insights from the PREDIMED Study. Prog. Cardiovasc. Dis. 2015, 58, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Martinez, O.; Ruiz, C.; Gutierrez-Ibanez, A.; Illescas-Montes, R.; Melguizo-Rodriguez, L. Benefits of Olive Oil Phenolic Compounds in Disease Prevention. Endocr. Metab. Immune Disord. Drug Targets 2018, 18, 333–340. [Google Scholar] [CrossRef]
- Estruch, R.; Ros, E.; Salas-Salvadó, J.; Covas, M.-I.; Corella, D.; Arós, F.; Gómez-Gracia, E.; Ruiz-Gutiérrez, V.; Fiol, M.; Lapetra, J.; et al. Primary Prevention of Cardiovascular Disease with a Mediterranean Diet Supplemented with Extra-Virgin Olive Oil or Nuts. N. Engl. J. Med. 2018, 378, e34. [Google Scholar] [CrossRef]
- Casas, R.; Sacanella, E.; Urpí-Sardà, M.; Corella, D.; Castañer, O.; Lamuela-Raventos, R.-M.; Salas-Salvadó, J.; Martínez-González, M.-A.; Ros, E.; Estruch, R. Long-Term Immunomodulatory Effects of a Mediterranean Diet in Adults at High Risk of Cardiovascular Disease in the PREvención con DIeta MEDiterránea (PREDIMED) Randomized Controlled Trial. J. Nutr. 2016, 146, 1684–1693. [Google Scholar] [CrossRef]
- Lopez-Moreno, J.; Garcia-Carpintero, S.; Gomez-Delgado, F.; Jimenez-Lucena, R.; Vals-Delgado, C.; Alcala-Diaz, J.F.; Roncero-Ramos, I.; Rangel-Zuñiga, O.A.; Yubero-Serrano, E.M.; Malagon, M.M.; et al. Endotoxemia is modulated by quantity and quality of dietary fat in older adults. Exp. Gerontol. 2018, 109, 119–125. [Google Scholar] [CrossRef]
- Juárez-Hernández, E.; Chávez-Tapia, N.C.; Uribe, M.; Barbero-Becerra, V.J. Role of bioactive fatty acids in nonalcoholic fatty liver disease. Nutr. J. 2016, 15, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tutino, V.; De Nunzio, V.; Caruso, M.G.; Bonfiglio, C.; Franco, I.; Mirizzi, A.; De Leonardis, G.; Cozzolongo, R.; Giannuzzi, V.; Giannelli, G.; et al. Aerobic Physical Activity and a Low Glycemic Diet Reduce the AA/EPA Ratio in Red Blood Cell Membranes of Patients with NAFLD. Nutrients 2018, 10, 1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, J.; Davison, G.; Marcora, S.M.; Mauger, A.R. Effect of a Mediterranean Type Diet on Inflammatory and Cartilage Degradation Biomarkers in Patients with Osteoarthritis. J. Nutr. Health Aging 2017, 21, 562–566. [Google Scholar] [CrossRef] [Green Version]
- Mayr, H.L.; Itsiopoulos, C.; Tierney, A.C.; Kucianski, T.; Radcliffe, J.; Garg, M.; Willcox, J.; Thomas, C.J. Ad libitum Mediterranean diet reduces subcutaneous but not visceral fat in patients with coronary heart disease: A randomised controlled pilot study. Clin. Nutr. ESPEN 2019, 32, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Morate, E.; Gimeno-Mallench, L.; Stromsnes, K.; Sanz-Ros, J.; Román-Domínguez, A.; Parejo-Pedrajas, S.; Inglés, M.; Olaso, G.; Gambini, J.; Mas-Bargues, C. Relationship between Diet, Microbiota, and Healthy Aging. Biomedicines 2020, 8, 287. [Google Scholar] [CrossRef] [PubMed]
- Meslier, V.; Laiola, M.; Roager, H.M.; De Filippis, F.; Roume, H.; Quinquis, B.; Giacco, R.; Mennella, I.; Ferracane, R.; Pons, N.; et al. Mediterranean diet intervention in overweight and obese subjects lowers plasma cholesterol and causes changes in the gut microbiome and metabolome independently of energy intake. Gut 2020, 69, 1258–1268. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: The NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayr, H.L.; Itsiopoulos, C.; Tierney, A.C.; Ruiz-Canela, M.; Hebert, J.R.; Shivappa, N.; Thomas, C.J. Improvement in dietary inflammatory index score after 6-month dietary intervention is associated with reduction in interleukin-6 in patients with coronary heart disease: The AUSMED heart trial. Nutr. Res. 2018, 55, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Gudban, N.; Yehuda, I.; Nasir, W.; Soboh, S.; Tamir, S.; Blum, A. Effect of Telemedicine Dietary Intervention for Endothelial Function in Patients with Type 2 Diabetes Mellitus on Mediterranean Diet. Isr. Med. Assoc. J. 2021, 23, 89–93. [Google Scholar] [PubMed]
- Papadaki, A.; Scott, J.A. The impact on eating habits of temporary translocation from a Mediterranean to a Northern European environment. Eur. J. Clin. Nutr. 2002, 56, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neale, E.P.; Batterham, M.J.; Tapsell, L.C. Consumption of a healthy dietary pattern results in significant reductions in C-reactive protein levels in adults: A meta-analysis. Nutr. Res. 2016, 36, 391–401. [Google Scholar] [CrossRef]
- Lankinen, M.; Uusitupa, M.; Schwab, U. Nordic Diet and Inflammation—A Review of Observational and Intervention Studies. Nutrients 2019, 11, 1369. [Google Scholar] [CrossRef] [Green Version]
- Tuomainen, M.; Kärkkäinen, O.; Leppänen, J.; Auriola, S.; Lehtonen, M.; Savolainen, M.J.; Hermansen, K.; Risérus, U.; Åkesson, B.; Thorsdottir, I.; et al. Quantitative assessment of betainized compounds and associations with dietary and metabolic biomarkers in the randomized study of the healthy Nordic diet (SYSDIET). Am. J. Clin. Nutr. 2019, 110, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Vogt, J.K.; Kristensen, M.; Hansen, L.B.S.; Ibrügger, S.; Mærkedahl, R.B.; Bahl, M.I.; Lind, M.V.; Nielsen, R.L.; Frøkiær, H.; et al. Whole grain-rich diet reduces body weight and systemic low-grade inflammation without inducing major changes of the gut microbiome: A randomised cross-over trial. Gut 2019, 68, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulven, S.M.; Holven, K.B.; Rundblad, A.; Myhrstad, M.C.W.; Leder, L.; Dahlman, I.; Mello, V.D.; de Schwab, U.; Carlberg, C.; Pihlajamäki, J.; et al. An Isocaloric Nordic Diet Modulates RELA and TNFRSF1A Gene Expression in Peripheral Blood Mononuclear Cells in Individuals with Metabolic Syndrome—A SYSDIET Sub-Study. Nutrients 2019, 11, 2932. [Google Scholar] [CrossRef] [Green Version]
- Leder, L.; Kolehmainen, M.; Narverud, I.; Dahlman, I.; Myhrstad, M.C.W.; de Mello, V.D.; Paananen, J.; Carlberg, C.; Schwab, U.; Herzig, K.-H.; et al. Effects of a healthy Nordic diet on gene expression changes in peripheral blood mononuclear cells in response to an oral glucose tolerance test in subjects with metabolic syndrome: A SYSDIET sub-study. Genes Nutr. 2016, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, A.S.; Ninomiya, K.; Uneyama, H. The Role of the Japanese Traditional Diet in Healthy and Sustainable Dietary Patterns around the World. Nutrients 2018, 10, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coe, C.L.; Miyamoto, Y.; Love, G.D.; Karasawa, M.; Kawakami, N.; Kitayama, S.; Ryff, C.D. Cultural and life style practices associated with low inflammatory physiology in Japanese adults. Brain Behav. Immun. 2020, 90, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.-W.; Yang, C.-Y.; Wu, H.-Y.; Liu, K.-L.; Su, C.-T.; Wu, C.-K.; Lee, J.-K.; Chiang, C.-K.; Cheng, H.-T.; Lien, Y.-C.; et al. Metabolic syndrome and abdominal fat are associated with inflammation, but not with clinical outcomes, in peritoneal dialysis patients. Cardiovasc. Diabetol. 2013, 12, 86. [Google Scholar] [CrossRef] [Green Version]
- Deng, K.; Zhang, X.; Liu, Y.; Cheng, G.P.; Zhang, H.P.; Wang, T.; Wang, L.; Li, W.M.; Wang, G.; Wood, L. Visceral obesity is associated with clinical and inflammatory features of asthma: A prospective cohort study. Allergy Asthma Proc. 2020, 41, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Sakane, N.; Osaki, N.; Takase, H.; Suzuki, J.; Suzukamo, C.; Nirengi, S.; Suganuma, A.; Shimotoyodome, A. The study of metabolic improvement by nutritional intervention controlling endogenous GIP (Mini Egg study): A randomized, cross-over study. Nutr. J. 2019, 18, 52. [Google Scholar] [CrossRef] [Green Version]
- Kushida, M.; Sugawara, S.; Asano, M.; Yamamoto, K.; Fukuda, S.; Tsuduki, T. Effects of the 1975 Japanese diet on the gut microbiota in younger adults. J. Nutr. Itional Biochem. 2019, 64, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Hempe, J.M.; McCarter, R.J.; Li, S.; Fonseca, V.A. Association between Inflammation and Biological Variation in Hemoglobin A1c in U.S. Nondiabetic Adults. J. Clin. Endocrinol. Metab. 2015, 100, 2364–2371. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lin, X.; Bloomgarden, Z.T.; Ning, G. The Jiangnan diet, a healthy diet pattern for Chinese. J. Diabetes 2020, 12, 365–371. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Ma, G.; Zhai, F.; Li, Y.; Hu, Y.; Feskens, E.J.M.; Yang, X. Dietary patterns and glucose tolerance abnormalities in Chinese adults. Diabetes Care 2009, 32, 1972–1976. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Zhang, X.; Xiang, Y.-B.; Yang, G.; Li, H.; Gao, Y.-T.; Zheng, W.; Shu, X.-O. Adherence to dietary guidelines and mortality: A report from prospective cohort studies of 134,000 Chinese adults in urban Shanghai. Am. J. Clin. Nutr. 2014, 100, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Database on Polyphenol Content in Foods-Phenol-Explorer. Available online: http://phenol-explorer.eu/ (accessed on 1 July 2021).
- Ford, L.; Theodoridou, K.; Sheldrake, G.N.; Walsh, P.J. A critical review of analytical methods used for the chemical characterisation and quantification of phlorotannin compounds in brown seaweeds. Phytochem. Anal. 2019, 30, 587–599. [Google Scholar] [CrossRef]
- FoodData Central. Available online: https://fdc.nal.usda.gov/index.html (accessed on 1 July 2021).
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A Comprehensive Review of Chemistry, Sources and Bioavailability of Omega-3 Fatty Acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Yerena, A.; Vallverdú-Queralt, A.; Mols, R.; Augustijns, P.; Lamuela-Raventós, R.M.; Escribano-Ferrer, E. Absorption and Intestinal Metabolic Profile of Oleocanthal in Rats. Pharmaceutics 2020, 12, 134. [Google Scholar] [CrossRef] [Green Version]
- Wiktorowska-Owczarek, A.; Berezińska, M.; Nowak, J.Z. PUFAs: Structures, metabolism and functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta BBA Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, S. Chapter Fatty Acids: From Membrane Ingredients to Signaling Molecules. In Biochemistry and Health Benefits of Fatty Acids; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Rucker, D.; Dhamoon, A.S. Physiology, Thromboxane A2. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Wang, B.; Wu, L.; Chen, J.; Dong, L.; Chen, C.; Wen, Z.; Hu, J.; Fleming, I.; Wang, D.W. Metabolism pathways of arachidonic acids: Mechanisms and potential therapeutic targets. Signal Transduct. Target. Ther. 2021, 6, 1–30. [Google Scholar]
- Tsuge, K.; Inazumi, T.; Shimamoto, A.; Sugimoto, Y. Molecular mechanisms underlying prostaglandin E2-exacerbated inflammation and immune diseases. Int. Immunol. 2019, 31, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Lau, K.E.; Lui, F. Physiology, Prostaglandin I2. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci.Ence Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- Layé, S.; Nadjar, A.; Joffre, C.; Bazinet, R.P. Anti-inflammatory effects of omega-3 fatty acids in the brain: Physiological mechanisms and relevance to pharmacology. Pharmacol. Rev. 2018, 70, 12–38. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Levy, B.D. Resolvins in inflammation: Emergence of the pro-resolving superfamily of mediators. J. Clin. Investig. 2018, 128, 2657–2669. [Google Scholar] [CrossRef] [PubMed]
- Duvall, M.G.; Levy, B.D. DHA- and EPA-derived resolvins, protectins, and maresins in airway inflammation. Eur. J. Pharmacol. 2016, 785, 144–155. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. Mechanisms of action of (n-3) fatty acids. J. Nutr. 2012, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.R.; Lei, C.Q. TAK1-TABs Complex: A Central Signalosome in Inflammatory Responses. Front. Immunol. 2021, 11. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Im, D.S. FFA4 (GPR120) as a fatty acid sensor involved in appetite control, insulin sensitivity and inflammation regulation. Mol. Asp. Med. 2018, 64, 92–108. [Google Scholar] [CrossRef]
- Gaspar, R.C.; Veiga, C.B.; Bessi, M.P.; Dátilo, M.N.; Sant’Ana, M.R.; Rodrigues, P.B.; de Moura, L.P.; da Silva, A.S.R.; Santos, G.A.; Catharino, R.R.; et al. Unsaturated fatty acids from flaxseed oil and exercise modulate GPR120 but not GPR40 in the liver of obese mice: A new anti-inflammatory approach. J. Nutr. Biochem. 2019, 66, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [Green Version]
- Calder, P.C. N-3 PUFA and Inflammation: From membrane to nucleus and from bench to bedside. In Proceedings of the Nutrition Society; Cambridge University Press: London, UK, 2020; Volume 79, pp. 404–416. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Draayer, K.; Kalman, D.S. Palm fruit bioactive complex (Pfbc), a source of polyphenols, demonstrates potential benefits for inflammaging and related cognitive function. Nutrients 2021, 13, 1127. [Google Scholar] [CrossRef] [PubMed]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Stromsnes, K.; Mas-Bargues, C.; Gambini, J.; Gimeno-Mallench, L. Protective Effects of Polyphenols Present in Mediterranean Diet on Endothelial Dysfunction. Available online: https://www.hindawi.com/journals/omcl/2020/2097096/ (accessed on 7 August 2020).
- Khor, B.H.; Narayanan, S.S.; Sahathevan, S.; Gafor, A.H.A.; Daud, Z.A.M.; Khosla, P.; Sabatino, A.; Fiaccadori, E.; Chinna, K.; Karupaiah, T. Efficacy of nutritional interventions on inflammatory markers in haemodialysis patients: A systematic review and limited meta-analysis. Nutrients 2018, 10, 397. [Google Scholar] [CrossRef] [Green Version]
- Ambriz-Pérez, D.L.; Leyva-López, N.; Gutierrez-Grijalva, E.P.; Heredia, J.B. Phenolic compounds: Natural alternative in inflammation treatment. A Review. Cogent Food Agric. 2016, 2. [Google Scholar] [CrossRef]
- Lago, J.H.G.; Toledo-Arruda, A.C.; Mernak, M.; Barrosa, K.H.; Martins, M.A.; Tibério, I.F.L.C.; Prado, C.M. Structure-Activity association of flavonoids in lung diseases. Molecules 2014, 19, 3570–3595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzocchella, L.; Fantini, M.; Benvenuto, M.; Masuelli, L.; Tresoldi, I.; Modesti, A.; Bei, R. Dietary Flavonoids: Molecular Mechanisms of Action as Anti-Inflammatory Agents. Recent Pat. Inflamm. Allergy Drug Discov. 2011, 5, 200–220. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta BBA Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Song, B.; Cho, K.S.; Lee, I.S. Therapeutic potential of volatile terpenes and terpenoids from forests for inflammatory diseases. Int. J. Mol. Sci. 2020, 21, 2187. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.S.; Lim, Y.R.; Lee, K.; Lee, J.; Lee, J.H.; Lee, I.S. Terpenes from forests and human health. Toxicol. Res. 2017, 33, 97–106. [Google Scholar] [CrossRef]
- José Serrano Vega, R.; Campos Xolalpa, N.; Josabad Alonso Castro, A.; Pérez González, C.; Pérez Ramos, J.; Pérez Gutiérrez, S. Terpenes from Natural Products with Potential Anti-Inflammatory Activity. In Terpenes and Terpenoids; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef] [Green Version]
- Heming, M.; Gran, S.; Jauch, S.L.; Fischer-Riepe, L.; Russo, A.; Klotz, L.; Hermann, S.; Schäfers, M.; Roth, J.; Barczyk-Kahlert, K. Peroxisome proliferator-activated receptor-γ modulates the response of macrophages to lipopolysaccharide and glucocorticoids. Front. Immunol. 2018, 9, 1. [Google Scholar] [CrossRef]
- Kim, D.S.; Lee, H.J.; Jeon, Y.D.; Han, Y.H.; Kee, J.Y.; Kim, H.J.; Shin, H.J.; Kang, J.; Lee, B.S.; Kim, S.H.; et al. Alpha-Pinene Exhibits Anti-Inflammatory Activity Through the Suppression of MAPKs and the NF-κB Pathway in Mouse Peritoneal Macrophages. Am. J. Chin. Med. 2015, 43, 731–742. [Google Scholar] [CrossRef]
- Kaminska, B. MAPK signalling pathways as molecular targets for anti-inflammatory therapy—From molecular mechanisms to therapeutic benefits. Biochim. Biophys. Acta BBA Proteins Proteom. 2005, 1754, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Qiu, X.; Yang, J. Comparing the In Vitro Antitumor, Antioxidant and Anti-Inflammatory Activities between Two New Very Long Chain Polyunsaturated Fatty Acids, Docosadienoic Acid (DDA) and Docosatrienoic Acid (DTA), and Docosahexaenoic Acid (DHA). Nutr. Cancer 2020, 1, 11. [Google Scholar] [CrossRef]
- Wellhauser, L.; Belsham, D.D. Activation of the omega-3 fatty acid receptor GPR120 mediates anti-inflammatory actions in immortalized hypothalamic neurons. J. Neuroinflamm. 2014, 11, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, X.; Gong, L.F.; Wu, Y.F.; Lin, Z.; Jiang, B.J.; Wu, L.; Yu, K.H. Urolithin A targets the PI3K/Akt/NF-κB pathways and prevents IL-1β-induced inflammatory response in human osteoarthritis: In vitro and in vivo studies. Food Funct. 2019, 10, 6135–6146. [Google Scholar] [CrossRef]
- Komatsu, W.; Kishi, H.; Yagasaki, K.; Ohhira, S. Urolithin A attenuates pro-inflammatory mediator production by suppressing PI3-K/Akt/NF-κB and JNK/AP-1 signaling pathways in lipopolysaccharide-stimulated RAW264 macrophages: Possible involvement of NADPH oxidase-derived reactive oxygen species. Eur. J. Pharmacol. 2018, 833, 411–424. [Google Scholar] [CrossRef]
- Ding, S.L.; Pang, Z.Y.; Chen, X.M.; Li, Z.; Liu, X.X.; Zhai, Q.L.; Huang, J.M.; Ruan, Z.Y. Urolithin a attenuates IL-1β-induced inflammatory responses and cartilage degradation via inhibiting the MAPK/NF-κB signaling pathways in rat articular chondrocytes. J. Inflamm. 2020, 17, 13. [Google Scholar] [CrossRef] [Green Version]
- Nile, S.H.; Ko, E.Y.; Kim, D.H.; Keum, Y.S. Screening of ferulic acid related compounds as inhibitors of xanthine oxidase and cyclooxygenase-2 with anti-inflammatory activity. Rev. Bras. Farmacogn. 2016, 26, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liu, J. Anti-Inflammatory Effects of p-coumaric Acid in LPS-Stimulated RAW264.7 Cells: Involvement of NF-κB and MAPKs Pathways. Med. Chem. 2016, 6, 327–330. [Google Scholar] [CrossRef]
- Lampiasi, N.; Montana, G. An in vitro inflammation model to study the Nrf2 and NF-κB crosstalk in presence of ferulic acid as modulator. Immunobiology 2018, 223, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zheng, Y.; Luo, Y.; Du, Y.; Zhang, X.; Fu, J. Curcumin inhibits LPS-induced neuroinflammation by promoting microglial M2 polarization via TREM2/TLR4/NF-κB pathways in BV2 cells. Mol. Immunol. 2019, 116, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Calabriso, N.; Scoditti, E.; Massaro, M.; Pellegrino, M.; Storelli, C.; Ingrosso, I.; Giovinazzo, G.; Carluccio, M.A. Multiple anti-inflammatory and anti-atherosclerotic properties of red wine polyphenolic extracts: Differential role of hydroxycinnamic acids, flavonols and stilbenes on endothelial inflammatory gene expression. Eur. J. Nutr. 2016, 55, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Csaki, C.; Nebrich, S.; Mobasheri, A. Resveratrol suppresses interleukin-1β-induced inflammatory signaling and apoptosis in human articular chondrocytes: Potential for use as a novel nutraceutical for the treatment of osteoarthritis. Biochem. Pharmacol. 2008, 76, 1426–1439. [Google Scholar] [CrossRef]
- Martinez, J.; Moreno, J.J. Effect of resveratrol, a natural polyphenolic compound, on reactive oxygen species and prostaglandin production. Biochem. Pharmacol. 2000, 59, 865–870. [Google Scholar] [CrossRef]
- Liang, Y.C.; Huang, Y.T.; Tsai, S.H.; Lin-Shiau, S.Y.; Chen, C.F.; Lin, J.K. Suppression of inducible cyclooxygenase and inducible nitric oxide synthase by apigenin and related flavonoids in mouse macrophages. Carcinogenesis 1999, 20, 1945–1952. [Google Scholar] [CrossRef] [Green Version]
- Miles, E.A.; Zoubouli, P.; Calder, P.C. Differential anti-inflammatory effects of phenolic compounds from extra virgin olive oil identified in human whole blood cultures. Nutrition 2005, 21, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Mu, M.M.; Chakravortty, D.; Sugiyama, T.; Koide, N.; Takahashi, K.; Mori, I.; Yoshida, T.; Yokochi, T. The Inhibitory Action of Quercetin on Lipopolysaccharide-Induced Nitric Oxide Production in RAW 264.7 Macrophage Cells. J. Endotoxin Res. 2001, 7, 431–438. [Google Scholar] [CrossRef]
- Chen, J.C.; Ho, F.M.; Chao, P.D.L.; Chen, C.P.; Jeng, K.C.G.; Hsu, H.B.; Lee, S.T.; Wen, T.W.; Lin, W.W. Inhibition of iNOS gene expression by quercetin is mediated by the inhibition of IκB kinase, nuclear factor-kappa B and STAT1, and depends on heme oxygenase-1 induction in mouse BV-2 microglia. Eur. J. Pharmacol. 2005, 521, 9–20. [Google Scholar] [CrossRef]
- Wu, Y.R.; Choi, H.J.; Kang, Y.G.; Kim, J.K.; Shin, J.W. In vitro study on anti-inflammatory effects of epigallocatechin-3-gallate-loaded nano- and microscale particles. Int. J. Nanomed. 2017, 12, 7007–7013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Y.; Chiou, Y.S.; Pan, M.H.; Shahidi, F. Anti-inflammatory activity of lipophilic epigallocatechin gallate (EGCG) derivatives in LPS-stimulated murine macrophages. Food Chem. 2012, 134, 742–748. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, Z.R. Anti-inflammatory effects of (−)-epicatechin in lipopolysaccharide-stimulated raw 264.7 macrophages. Trop. J. Pharm. Res. 2014, 13, 1415–1419. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.W.; Tan, X.; Sun, J.Y.; Gu, C.M.; Liu, C.; Guo, X. Catechin attenuates TNF-α induced inflammatory response via AMPK-SIRT1 pathway in 3T3-L1 adipocytes. PLoS ONE 2019, 14, e0217090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Peng, W.H.; Tsai, K.D.; Hsu, S.L. Luteolin suppresses inflammation-associated gene expression by blocking NF-κB and AP-1 activation pathway in mouse alveolar macrophages. Life Sci. 2007, 81, 1602–1614. [Google Scholar] [CrossRef]
- Du, Z.R.; Feng, X.Q.; Li, N.; Qu, J.X.; Feng, L.; Chen, L.; Chen, W.F. G protein-coupled estrogen receptor is involved in the anti-inflammatory effects of genistein in microglia. Phytomedicine 2018, 43, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Kim, B.; Yang, Y.; Pham, T.X.; Park, Y.K.; Manatou, J.; Koo, S.I.; Chun, O.K.; Lee, J.Y. Berry anthocyanins suppress the expression and secretion of proinflammatory mediators in macrophages by inhibiting nuclear translocation of NF-κB independent of NRF2-mediated mechanism. J. Nutr. Biochem. 2014, 25, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.-W.; Lee, W.S.; Shin, S.C.; Kim, G.-Y.; Choi, B.T.; Choi, Y.H. Anthocyanins Downregulate Lipopolysaccharide-Induced Inflammatory Responses in BV2 Microglial Cells by Suppressing the NF-κB and Akt/MAPKs Signaling Pathways. Int. J. Mol. Sci. 2013, 14, 1502–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, D.X.; Yanagita, T.; Uto, T.; Masuzaki, S.; Fujii, M. Anthocyanidins inhibit cyclooxygenase-2 expression in LPS-evoked macrophages: Structure-activity relationship and molecular mechanisms involved. Biochem. Pharmacol. 2005, 70, 417–425. [Google Scholar] [CrossRef]
- Liang, N.; Sang, Y.; Liu, W.; Yu, W.; Wang, X. Anti-Inflammatory Effects of Gingerol on Lipopolysaccharide-Stimulated RAW 264.7 Cells by Inhibiting NF-κB Signaling Pathway. Inflammation 2018, 41, 835–845. [Google Scholar] [CrossRef]
- Dugasani, S.; Pichika, M.R.; Nadarajah, V.D.; Balijepalli, M.K.; Tandra, S.; Korlakunta, J.N. Comparative antioxidant and anti-inflammatory effects of [6]-gingerol, [8]-gingerol, [10]-gingerol and [6]-shogaol. J. Ethnopharmacol. 2010, 127, 515–520. [Google Scholar] [CrossRef]
- Scotece, M.; Gómez, R.; Conde, J.; Lopez, V.; Gómez-Reino, J.J.; Lago, F.; Smith, A.B.; Gualillo, O. Further evidence for the anti-inflammatory activity of oleocanthal: Inhibition of MIP-1α and IL-6 in J774 macrophages and in ATDC5 chondrocytes. Life Sci. 2012, 91, 1229–1235. [Google Scholar] [CrossRef]
- Hoon Cha, J.; Kyoung Kim, W.; Wha Ha, A.; Hwan Kim, M.; Jeong Chang, M. Anti-inflammatory effect of lycopene in SW480 human colorectal cancer cells. Nutr. Res. Pract. 2017, 11, 90–96. [Google Scholar]
- Bai, S.K.; Lee, S.J.; Na, H.J.; Ha, K.S.; Han, J.A.; Lee, H.; Kwon, Y.G.; Chung, C.K.; Kim, Y.M. β-carotene inhibits inflammatory gene expression in lipopolysaccharide-stimulated macrophages by suppressing redox-based NF-κB activation. Exp. Mol. Med. 2005, 37, 323–334. [Google Scholar] [CrossRef]
- Maione, F.; Cantone, V.; Pace, S.; Chini, M.G.; Bisio, A.; Romussi, G.; Pieretti, S.; Werz, O.; Koeberle, A.; Mascolo, N.; et al. Anti-inflammatory and analgesic activity of carnosol and carnosic acid in vivo and in vitro and in silico analysis of their target interactions. Br. J. Pharmacol. 2017, 174, 1497–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwager, J.; Richard, N.; Fowler, A.; Seifert, N.; Raederstorff, D. Carnosol and related substances modulate chemokine and cytokine production in macrophages and chondrocytes. Molecules 2016, 21, 465. [Google Scholar] [CrossRef] [Green Version]
- Minatel, I.O.; Borges, C.V.; Ferreira, M.I.; Gomez, H.A.G.; Chen, C.-Y.O.; Lima, G.P.P. Phenolic Compounds: Functional Properties, Impact of Processing and Bioavailability. In Phenolic Compounds—Biological Activity; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Grigore, A. Plant Phenolic Compounds as Immunomodulatory Agents. In Phenolic Compounds—Biological Activity; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Jafarirad, S.; Ayoobi, N.; Karandish, M.; Jalali, M.-T.; Haghighizadeh, M.H.; Jahanshahi, A. Dark Chocolate Effect on Serum Adiponectin, Biochemical and Inflammatory Parameters in Diabetic Patients: A Randomized Clinical Trial. Int. J. Prev. Med. 2018, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Kuebler, U.; Arpagaus, A.; Meister, R.E.; von Känel, R.; Huber, S.; Ehlert, U.; Wirtz, P.H. Dark chocolate attenuates intracellular pro-inflammatory reactivity to acute psychosocial stress in men: A randomized controlled trial. Brain Behav. Immun. 2016, 57, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenols | Ref | Source (mg/100 g Fresh or 100 mL) |
|---|---|---|
| Ferulic acid | [53] | Green raw Olive (3), Aubergine (3), Refined Flour (14), Whole Grain Flour (72), Dark Chocolate (24) |
| Quercetin | [53] | Dark Chocolate (25), Blank Elderberry (42), Capers (33), Raw Shallot (2) |
| Kaempferol | [53] | Capers (104), Cumin (39), Common bean [Black] (2), Common bean [White] (0,9) and (o-glucoside 40) |
| Cyanidin | [53] | Common bean [Black] (1.6), Strawberry (0.5), Red raspberry (0.5), Black Olive (11), Red lettuce (0.6), Black elderberry (o-glucoside 794) |
| Epicatechin | [53] | Cocoa powder (158), Cider apple (29), Black Olive (4.1), broad bean (23), Blackberry (12) |
| Caffeic acid | [53] | Bread Rye and Whole Grain Flour (0.8), Parboiled Rice (0.34), Black Chokeberry (141), Dried Common Sage (26), Oregano (21), Rosemary (10), Green Chicory (2.3), Dried Common Thyme (21), Raw Potato (1,6) |
| Resveratrol | [53] | Lingonberry (3), Red currant (1.6), Red Wine (3–0.25), Red Wine (o.gluciside 0.3), Green Grape (0.24), Dark Chocolate (o-glucoside 0.1) |
| Tangeretin | [53] | Orange Juice (0,03–0.06) |
| Catechin | [53] | Red wine (6.8), Dark Chocolate (21), Powder Cocoa (108), Barley (1.2), Cider apple (5.6), Rhubarb (2.2), Plum (4.6), Rosemary (2.7), Pecan Nut (7,2), Broad been (13), Lentils (0.3) |
| Urolithin | [53] | Pomegranate Juice (Punicalagin 44), Red Raspberry (76) |
| Apigenin | [53] | Extra Virgin Olive Oil (1.2), Commo Sage (2.4), Oregano (3.3), Rosemary (0.6), Black Olive (o-glucoside 8.2), Artichoke (o-glucoside 7.4), Celery Seed (o-glucoside 111), Lentils (o-glucoside 0.62) |
| Carnosol | [53] | Rosemary (53) |
| Luteolin | [53] | Artichoke heads (42), thyme (40), olive oil extra (0.4) |
| Genistein | [53] | Soy (10–0.1), Black bean (0.6), White Bean (0.3) |
| Phlorotannins | [54] | Fucus vesiculosus, dry (24), Ascophyllum nodosum dry (16) |
| Epicatechin | [53] | Cocoa Powder (158), Apple Cider (29), Black Olive (4.1), Broad Bean (23), Blackberry (12) |
| Gallic acid | [53] | Blackberry (4.7), Red Wine (3.6), Black Tea (4.7), Cloves (458), Oregano (5.2), Vinegar (2.6), Chestnut (480), Chicory (26) |
| Gingerol | [53] | Ginger Dried (187) |
| β-carotene | [55] | Baked Sweet Potatoes (11), Carrot Raw (8.3), Spinach (6.3), Romaine Lettuce (5.2) |
| ω-3 fatty acids | [56] | Cod (Gadus morhua) (40), Perca (perca fluviatis) (35), Salmon (2000), Mackerel (2300), Herring (1500) |
| Oleocanthal | [57] | Virgin Olive Oil (5.4) |
| Type | Compound | Mechanism | Model | Dose Exhibiting Effects | Reference |
|---|---|---|---|---|---|
| Polyunsaturated Fatty Acids | |||||
| ω-3 LC-PUFA | Docosahexaenoic acid (DHA) | Reduction of intracellular ROS levels Decreased expressions of IL-6, IFN-γ, MCP-1, TNF-α, IL-1β and IL-10 | PMA-stimulated monocyte cell line THP-1 | 300 µM | [90] |
| Reduction in phosphorylation and activation of TAK1 and NF-κB Decreased TNF-α production Activation of GP120, thus blocking IKK-β/NF-κB pathway by retaining TAB1 through β-arrestin-2 | TNF-α-stimulated rHypoE-7 cell line from the embryonic rat hypothalamus | 100 µM | [91] | ||
| ω-3 LC-PUFA | Docosatrienoic acid (DTA) | Reduction of intracellular ROS levels Decreased expressions of IL-6, IFN-γ, MCP-1, TNF-α, IL-1β and IL-10 | PMA-stimulated monocyte cell line THP-1 | 300 µM | [90] |
| ω-6 LC-PUFA | Docosadienoic acid (DDA) | Reduction of intracellular ROS levels Decreased expressions of IL-6, IFN-γ, MCP-1, TNF-α, IL-1β and IL-10 | PMA-stimulated monocyte cell line THP-1 | 300 µM | [90] |
| Simple Phenolic Compounds | |||||
| Hydrobenzoic acid | Urolithin A | Downregulation of COX-2 and iNOS expression and decreased PGE2 an NO generation Inhibition of TNF-α and IL-6 production Prevention of IκB-α phosphorylation and p65 translocation into the nucleus Decreased PI3K and Akt phosphorylation, thus amelioration of PI3K/Akt/NF-κB pathway activation | Primary human osteoarthritis chondrocytes | 30 µM | [92] |
| Inhibition of Akt and JNK phosphorylation Suppression of NF-κB and AP-1 activation Inhibition of NADPH oxidase (NOX) and reduction of intracellular accumulation of ROS Reduction in TNF-α and IL-6 production | LPS-activated RAW264.7 macrophages ICR-mouse resident peritoneal macrophages | 10–20–40 µM | [93] | ||
| Inhibition of phosphorylation of MAPK pathway members (ERK1/2, JNK, p38) Suppression of COX-2, iNOS and MMPsInhibition of p65 phosphorylation and NF-κB pathway activation | IL-1β stimulated primary chondrocytes from Sprague Dawley rats | 15 µM | [94] | ||
| Hydrobenzoic acid | Gallic acid | Inhibition of xanthine oxidase Inhibition of COX-2 Inhibition of TNF-α, IL-6 | LPS-stimulated THP-1 cells | 10 µM | [95] |
| Hydroxycinnamic acid | p-Coumaric acid | Suppression of COX-2, iNOS, TNF-α and IL-1β expression Inhibition of IκB-α phosphorylation and nuclear translocation of p65, preventing activation of NF-κB pathway Inhibition of phosphorylation of ERK1/2 and JNK | LPS-activated RAW264.7 macrophages | 50–100 µg/mL | [96] |
| Hydroxycinnamic acid | Ferulic acid | Inhibition of xanthine oxidase Inhibition of COX-2 Inhibition of TNF-α, IL-6 | LPS-stimulated THP-1 | 10 µM | [95] |
| Inhibition of NF-κB translocation into the nucleus Inhibition of IKK/TAK1 activation Inhibition of Nrf2 pathway by impairing the reduction of cysteine residues in Keap1 Reduced MnSOD and TLR4 protein levels | LPS-activated RAW264.7 macrophages | 100 µM | [97] | ||
| Hydroxycinnamic acid | Caffeic acid | Inhibition of xanthine oxidase Inhibition of COX-2 Inhibition of TNF-α, IL-6 | LPS-stimulated THP-1 | 10 µM | [95] |
| Hydroxicinnamic acid | Curcumin | Inhibition of the synthesis of proinflammatory mediators IL-1β, IL-6, and iNOS and NO production Increased production of anti-inflammatory cytokines IL-6, IL-10, and Arg-1 Microglial polarization to M2 phenotype (decreased iNOS and increased CD206 immunoreactivity) Increased TREM2 expression and inhibition of TLR4 expression and p65 NF-κB phosphorylation | Murine BV2 microglial cell line | 5–10 µM | [98] |
| Polyphenols | |||||
| Stilbene | Resveratrol | Inhibition of NF-κB and AP-1 activation Downregulation of ICAM-1, VCAM-1, E-selectin expression Downregulation of MCP-1, M-CSF expression | Human umbilical vein endothelial cells (HUVEC) | 10–25 µM | [99] |
| Inhibition of IL-1β-induced VEGF, COX-2, MMP-3, MMP-9 gene expressionPrevention of IκB-α degradation (by inhibiting proteasome function) and NF-κB activation | Isolated primary human articular chondrocytes | 100 µM | [100] | ||
| Inhibition of ROS (O2−, H2O2) production Inhibition of arachidonic acid release, COX-2 induction and PGE2 synthesis | CD-1-mouse resident peritoneal macrophages stimulated by LPS or PMA | 30 µM | [101] | ||
| Flavonol | Kaempferol | Suppression of COX-2 and iNOS expression Inhibition of PGE2 production | LPS-activated RAW264.7 macrophages | 15–25 µM | [102] |
| Inhibition of NF-κB and AP-1 activation Downregulation of ICAM-1, VCAM-1, E-selectin expression Inhibition of MCP-1 synthesis and secretion | Human umbilical vein endothelial cells (HUVEC) | 10–25 µM | [99] | ||
| Inhibition of PGE2 production | LPS-stimulated human mononuclear cells | 10–100 µM | [103] | ||
| Flavonol | Myricetin | Reduction in NF-κB and AP-1 activation Downregulation of ICAM-1, VCAM-1, E-selectin expression Inhibition of MCP-1 synthesis and secretion | Human umbilical vein endothelial cells (HUVEC) | 10–25 µM | [99] |
| Flavonol | Quercetin | Inhibition of NO production and iNOS protein expression Stabilization of IκB-α and IκB-β preventing NF-κB activation Inhibition of activation of MAPK (Erk1/2, p38, JNK/SAPK) Inhibition of phosphorylation and activation of JAK/STAT | LPS-activated RAW264.7 macrophages | 50 µM | [104] |
| Inhibition of NF-κB and AP-1 activation Downregulation of ICAM-1, VCAM-1, E-selectin expression Downregulation of MCP-1, M-CSF expression | Human umbilical vein endothelial cells (HUVEC) | 10–25 µM | [99] | ||
| Inhibition of iNOS expression and NO production Attenuation of IκB-α phosphorylation and NF-κB activation Inhibition of DNA binding activity of AP-1 and STAT1 Induction of heme oxygenase-1 (HO-1) expression, possibly through Src family members | Murine BV2 microglial cell line | 3–10–30 µM | [105] | ||
| Flavanol | Epigallocatechin 3-gallate/epigallocatechin gallate (EGCG) | Suppression of TNF-α, IL-1β, and IL-6 expression | Human dermal fibroblasts | 50 µM | [106] |
| Downregulation of COX-2 and iNOS expression | LPS-activated RAW264.7 macrophages | 20–40 µM | [107] | ||
| Flavanol | (-)-epicatechins | Inhibition of the production of NO, PGE2, TNF-α, IL-6 | LPS-activated RAW264.7 macrophages | 5–25–50 µM | [108] |
| Flavanol | Catechin | Inhibition of gene expression of pro-inflammatory cytokines (IL-1α, IL-1β, IL-6, IL-12p35) and enzymes (COX-2, iNOS) Up-regulation of anti-inflammatory cytokines (IL-4, IL10) Inhibition of the activation of NF-κB, AMPK, FOXO3a and SIRT1 | TNF-α induced 3T3-L1 adipocytes | 10–25–50–100 μg/mL | [109] |
| Flavone | Apigenin | Suppression of COX-2 and iNOS expression Inhibition of PGE2 and NO production Inhibition of IKK activity preventing NF-κB activation | LPS-activated RAW264.7 macrophages | 5–15–25 µM | [102] |
| Flavone | Luteolin | Suppression of TNF-α, IL-6, iNOS, COX-2 gene expression Decreased Akt and IKK phosphorylation Blockade of NF-κB and AP-1 activation Inhibition of ROS generation | LPS-activated RAW264.7 macrophages | 5–10–25 µM | [110] |
| Isoflavone | Genistein | Suppression of COX-2 and iNOS expression Inhibition of PGE2 production | LPS-activated RAW264.7 macrophages | 15–25 µM | [102] |
| Suppression of protein expression of iNOS, COX-2, TNF-α, IL-1β, IL-6 Prevention of MAPKs and NF-κB pathways activation Up-regulation of G protein-coupled estrogen receptor (GPER) gene expression | Murine BV2 microglial cell line Primary microglia cell culture | 10 µM | [111] | ||
| Anthocyanins (Flavonoid) | Anthocyanins-rich extract from berries | Reduction in TNF-α secretion Inhibition of NF-κB translocation into the nucleus Reduction of cellular ROS levels Downregulation of IL-1β and NADPH oxidases 1 (NOX-1) expression | LPS-activated RAW264.7 macrophages | 20 µg/mL | [112] |
| Anthocyanins (Flavonoid) | Anthocyanins-rich extract | Inhibition of COX-2 and iNOS expression, and inhibition of PGE2 and NO production Inhibition of TNF-α and IL-1β expression Prevention of IκB-α degradation and NF-κB translocation Inhibition of Akt and MAPKs (ERK, JNK, p38) phosphorylation | Murine BV2 microglial cell line | 50–100 µg/mL | [113] |
| Anthocyanins (Flavonoid) | Delphinidin | Inhibition of COX-2 expression Inhibition of phosphorylation of c-Jun, thus inhibiting AP-1 Inhibition of NF-κB activation by blocking IκB-α degradation and p65 translocation Inhibition of the three MAPK signalling pathways (JNK, ERK, p38) | LPS-activated RAW264.7 macrophages | 50–75–100 µM | [114] |
| Other compunds | Gingerol | Decreased PGE2 and NO secretion Decreased COX-2, iNOS, TNF-α, IL-1β, IL-6 expression Inhibition of NF-κB activation by suppressing phosphorylation of IκBα and p65 | LPS-activated RAW264.7 macrophages | 50–100–200–300 μg/mL | [115] |
| Decreased of ROS production (in PMNs) Inhibition of NO production (in macrophages) Inhibition of PGE2 production (in macrophages) | Human polymorphonuclear neutrophils (PMN) LPS-activated RAW264.7 macrophages | 6 µM | [116] | ||
| Other compunds | Oleocanthal | Inhibition of iNOS expression and NO production Suppression of MIP-1α, IL-6, IL-1β, TNF-α,and GM-CSF expression | Murine macrophages J774 Murine chondrocytes ATDC5 | 15–50 µM | [117] |
| Terpenes and Terpenoids | |||||
| Terpenoid (carotenoid) | Lycopene | Downregulation of TNF-α, IL-1β, IL-6, iNOS, and COX-2 expression Inhibition of PGE2 and NO production Decreased JNK, ERK and NF-κB protein expression | LPS-stimulated SW 480 human colorectal cancer cells | 20–30 µM | [118] |
| Terpenoid (carotenoid) | β-carotene | Suppression of expression of COX-2, iNOS and TNF-α and IL-1β pro-forms PGE2, NO and ROS production Inhibition of IκB-α degradation and NF-κB activation | LPS-activated RAW264.7 macrophages BALB/c mice peritoneal macrophages stimulated by LPS | 50 µM | [119] |
| Diterpene | Carnosol and carnosic acid | Inhibition of LOX and mPGES-1 (microsomal PGE2 synthase-1) enzymes and formation of arachidonic acid-derived eicosanoids | Activated human primary monocytes and neutrophils | 3–30 µM | [120] |
| Reduction in NO and PGE2 production Inhibition of COX-2 activity Downregulation of iNOS, IL-1α, IL-6 or CXCL10/IP-10 genes expression (in macrophages) Downregulation of CCL5/RANTES and CXCL10/IP-10 gene expression (in SW1353 cells) Downregulation of interleukine (IL-1α, IL-1β, IL-6) and chemokine genes expression (CXCL8/IL-8, CCL20/MIP-3α, CCL5/RANTES) Inhibition of NF-κB (p65) translocation into the nucleus | LPS-stimulated RAW264.7 macrophages IL-1β activated chondrosarcoma SW1353 cells IL-1β activated primary human articular chondrocytes | 12.5 µM | [121] | ||
| Breakfast | g | Anti-Inflammatory Compound (mg/100 g) | mg | ||
| Wholemeal bread | 60 | 72 | Ferulic Acid | 43.20 | |
| Olive oil | 10 | 5.4 | Oleocanthal | 0.54 | |
| 1.2 | Apigenin | 0.12 | |||
| 0.4 | Luteolin | 0.04 | |||
| Soy milk | 200 | 3.7 | Genistein | 7.40 | |
| Blackberries | 50 | 4.7 | Gallic Acid | 2.35 | |
| 12 | Epicathechin | 6.00 | |||
| Red currant | 50 | 1.6 | Resveratrol | 0.80 | |
| Morning Snack | g | Anti-inflammatory compound (mg/100 g) | mg | ||
| Pommegranate juice | 200 | 44 | Punicalgin | 88.00 | |
| Pecan nuts | 30 | 7.2 | Catechin | 2.16 | |
| Chesnuts | 40 | 480 | Gallic Acid | 192.00 | |
| Lunch | g | Anti-inflammatory compound (mg/100 g) | mg | ||
| Salad | Rhubarb | 122 | 2.2 | Catechin | 2.68 |
| Black olives | 16 | 4.1 | Epicatechin | 0.65 | |
| 8.2 | Apigenin | 1.31 | |||
| Green olives | 16 | 3 | Ferulic Acid | 0.48 | |
| Capers | 8.6 | 33 | Quercetin | 2.83 | |
| 104 | Kaempferol | 8.94 | |||
| Tomate | 100 | 4.2 | Lycopene | 4.20 | |
| Vinegar | 10 | 2.6 | Gallic Acid | 0.26 | |
| Olive oil | 10 | 5.4 | Oleocanthal | 0.54 | |
| 1.2 | Apigenin | 0.12 | |||
| 0.4 | Luteolin | 0.04 | |||
| Lentils and vegetables stew | Lentils | 40 | 0.3 | Catechin | 0.12 |
| Black bean | 20 | 0.6 | Genistein | 0.12 | |
| 2 | Kaempferol | 0.40 | |||
| Carrots | 50 | 8.3 | Beta-carotene | 4.15 | |
| Tomate | 90 | 4.2 | Lycopene | 3.78 | |
| Shallot | 25 | 2 | Quercetin | 0.50 | |
| Olive oil | 10 | 5.4 | Oleocanthal | 0.54 | |
| 1.2 | Apigenin | 0.12 | |||
| 0.4 | Luteolin | 0.04 | |||
| Oregano | 1 | 21 | Caffeic Acid | 0.21 | |
| 3.3 | Apigenin | 0.03 | |||
| 5.2 | Gallic Acid | 0.05 | |||
| Cumin | 1 | 39 | Kaempferol | 0.39 | |
| Cloves | 1 | 458 | Gallic Acid | 4.58 | |
| Rye Bread | 60 | 0.8 | Epicathechin | 0.48 | |
| Red wine | 100 | 3 | Resveratrol | 3.00 | |
| 6.8 | Catechin | 6.80 | |||
| 3.6 | Gallic Acid | 3.60 | |||
| Plum | 200 | 4.6 | Catechin | 9.20 | |
| Dark chocolate | 20 | 25 | Quercetin | 5.00 | |
| 0.1 | Resveratrol | 0.02 | |||
| 21 | Catechin | 4.20 | |||
| 24 | Ferulic Acid | 4.80 | |||
| Afternoon Snack | g | Anti-inflammatory compound (mg/100 g) | mg | ||
| Orange juice | 200 | 0.06 | Tangerin | 0.12 | |
| Pecan nut | 30 | 7.2 | Catechin | 2.16 | |
| Dinner | g | Anti-inflammatory compound (mg/100 g) | mg | ||
| Grilled salmon with vegetables | Salmon | 150 | 2000 | ω-3 PUFA | 3000.00 |
| Olive oil | 10 | 5.4 | Oleocanthal | 0.54 | |
| 1.2 | Apigenin | 0.12 | |||
| 0.4 | Luteolin | 0.04 | |||
| Aubergine | 150 | 3 | Ferulic Acid | 4.50 | |
| Artichocke | 70 | 7.4 | Apigenin | 5.18 | |
| Rosemary | 1 | 10 | Caffeic Acid | 0.10 | |
| 2.7 | Catechin | 0.027 | |||
| 0.6 | Apigenin | 0.006 | |||
| Bread rye | 60 | 0.8 | Epicathechin | 0.48 | |
| Red wine | 100 | 3 | Resveratrol | 3.00 | |
| 6.8 | Catechin | 6.80 | |||
| 3.6 | Gallic Acid | 3.60 | |||
| Green Grape | 200 | 0.4 | Resveratrol | 0.80 | |
| Dark chocolate | 20 | 25 | Quercetin | 5.00 | |
| 0.1 | Resveratrol | 0.02 | |||
| 21 | Catechin | 4.20 | |||
| 24 | Ferulic Acid | 4.80 | |||
| Compound | Total mg |
|---|---|
| Ferulic Acid | 57.78 |
| Oleocanthal | 2.16 |
| Apigenin | 7.00 |
| Genistein | 7.52 |
| Gallic Acid | 204.09 |
| Epicathechin | 7.62 |
| Resveratrol | 7.64 |
| Punicalgin | 88.00 |
| Catechin | 41.02 |
| Quercetin | 13.30 |
| Kaempferol | 9.73 |
| Lycopene | 7.98 |
| Luteolin | 0.16 |
| beta-carotene | 4.15 |
| Caffeic Acid | 0.31 |
| Tangerin | 0.12 |
| n-PUFA | 3000.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stromsnes, K.; Correas, A.G.; Lehmann, J.; Gambini, J.; Olaso-Gonzalez, G. Anti-Inflammatory Properties of Diet: Role in Healthy Aging. Biomedicines 2021, 9, 922. https://doi.org/10.3390/biomedicines9080922
Stromsnes K, Correas AG, Lehmann J, Gambini J, Olaso-Gonzalez G. Anti-Inflammatory Properties of Diet: Role in Healthy Aging. Biomedicines. 2021; 9(8):922. https://doi.org/10.3390/biomedicines9080922
Chicago/Turabian StyleStromsnes, Kristine, Angela G. Correas, Jenny Lehmann, Juan Gambini, and Gloria Olaso-Gonzalez. 2021. "Anti-Inflammatory Properties of Diet: Role in Healthy Aging" Biomedicines 9, no. 8: 922. https://doi.org/10.3390/biomedicines9080922
APA StyleStromsnes, K., Correas, A. G., Lehmann, J., Gambini, J., & Olaso-Gonzalez, G. (2021). Anti-Inflammatory Properties of Diet: Role in Healthy Aging. Biomedicines, 9(8), 922. https://doi.org/10.3390/biomedicines9080922