
Neuroactive Amino Acid Profile in Autism Spectrum Disorder: Results from a Clinical Sample
 , , , , and
, , , , and
Abstract
:1. Introduction
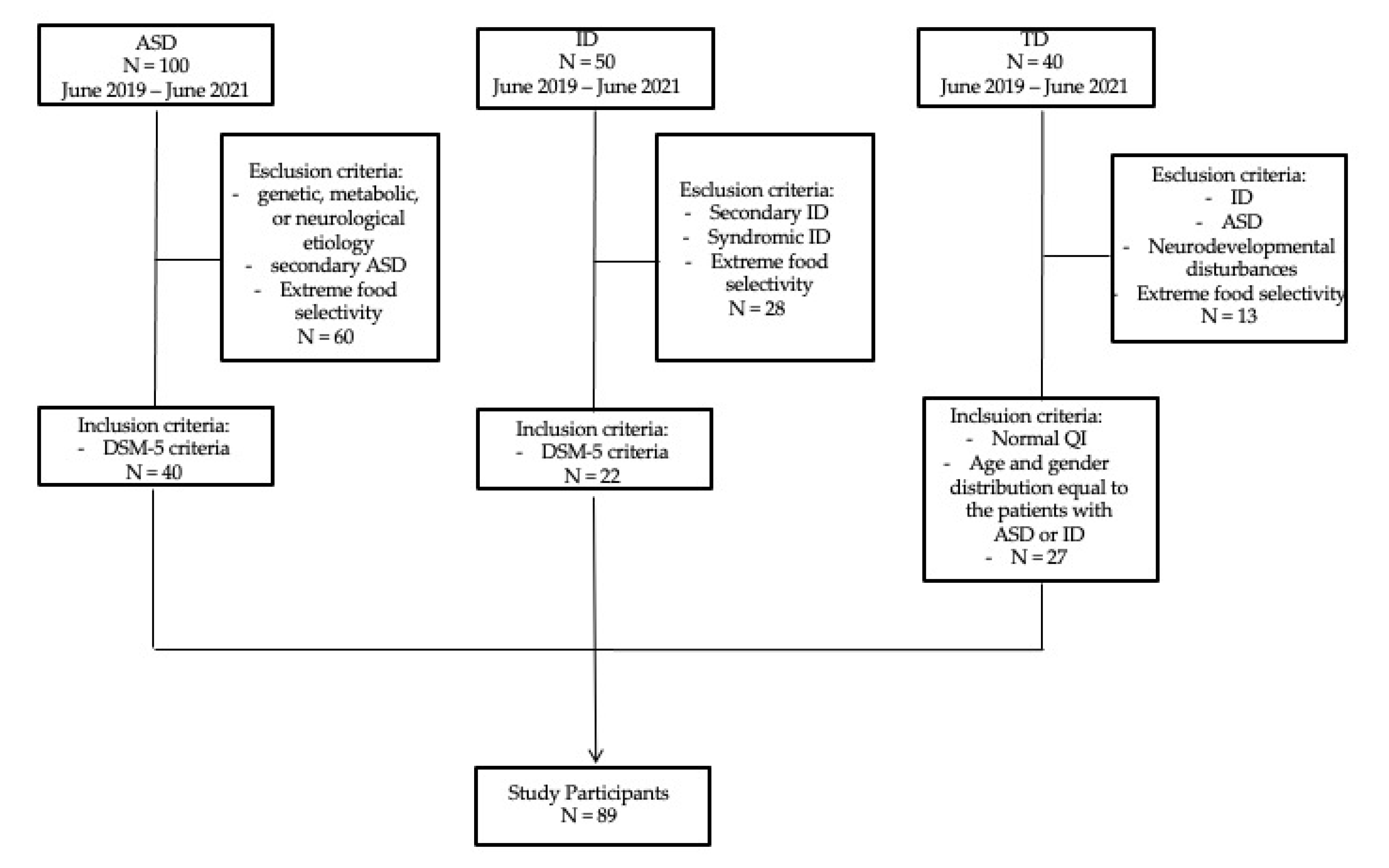
2. Materials and Methods
Ethical Statement
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; Text Revision; American Psychiatric Association: Washington, DC, USA, 2022. [Google Scholar]
- Lai, M.C.; Lombardo, M.V.; Baron-Cohen, S. Autism. Lancet 2014, 383, 896–910. [Google Scholar] [CrossRef]
- Zheng, H.F.; Wang, W.Q.; Li, X.M.; Rauw, G.; Baker, G.B. Body fluid levels of neuroactive amino acids in autism spectrum disorders: A review of the literature. Amino Acids. 2017, 49, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Bugajska, J.; Berska, J.; Wojtyto, T.; Bik-Multanowski, M.; Sztefko, K. The amino acid profile in blood plasma of young boys with autism. Psychiatr. Pol. 2017, 51, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Coyle, J.T. Glutamate and schizophrenia: Beyond the dopamine hypothesis. Cell. Mol. Neurobiol. 2006, 26, 365–384. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Rutter, M.; Le Couteur, A. Autism diagnostic interview-revised: A revised version of a diagnostic interview for caregivers of individuals with possible pervasive developmental disorders. J. Autism Dev. Disord. 1994, 24, 659–685. [Google Scholar] [CrossRef] [PubMed]
- Lord, C.; Risi, S.; Lambrecht, L.; Cook, E.H., Jr.; Leventhal, B.L.; DiLavore, P.C.; Pickles, A.; Rutter, M. The Autism Diagnostic Observation Schedule—Generic: A Standard Measure of Social and Communication Deficits Associated with the Spectrum of Autism. J. Autism Dev. Disord. 2000, 30, 205–223. [Google Scholar] [CrossRef]
- Luiz, D.M.; Foxcroft, C.D.; Povey, J.-L. The Griffiths Scales of Mental Development: A Factorial Validity Study. South Afr. J. Psychol. 2006, 36, 192–214. [Google Scholar] [CrossRef]
- O’Donnell, L. The Wechsler Intelligence Scale for Children. In Practitioner's Guide to Assessing Intelligence and Achievement, 4th ed.; Naglieri, J.A., Goldstein, S., Eds.; John Wiley & Sons Inc.: Hoboken, NJ, USA, 2009; pp. 153–190. [Google Scholar]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef] [Green Version]
- Barone, R.; Bastin, J.; Djouadi, F.; Singh, I.; Karim, M.A.; Ammanamanchi, A.; McCarty, P.J.; Delhey, L.; Shannon, R.; Casabona, A.; et al. Mitochondrial Fatty Acid β-Oxidation and Resveratrol Effect in Fibroblasts from Patients with Autism Spectrum Disorder. J. Pers. Med. 2021, 11, 510. [Google Scholar] [CrossRef]
- Arnold, G.L.; Hyman, S.L.; Mooney, R.A.; Kirby, R.S. Plasma amino acids profiles in children with autism: Potential risk of nutritional deficiencies. J. Autism Dev. Disord. 2003, 33, 449–454. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Audhya, T.; McDonough-Means, S.; Rubin, R.A.; Quig, D.; Geis, E.; Gehn, E.; Loresto, M.; Mitchell, J.; Atwood, S.; et al. Nutritional and metabolic status of children with autism vs. neurotypical children, and the association with autism severity. Nutr. Metab. 2011, 8, 34. [Google Scholar] [CrossRef] [Green Version]
- TirouvanziamTetyana, R.; Obukhanych, T.V.; Laval, J.; Aronov, P.A.; Libove, R.; Banerjee, A.G.; Parker, K.; O’Hara, R.; Herzenberg, L.A.; Hardan, A.Y. Distinct plasma profile of polar neutral amino acids, leucine, and glutamate in children with Autism Spectrum Disorders. J. Autism Dev. Disord. 2012, 42, 827–836. [Google Scholar] [CrossRef]
- ElBaz, F.M.; Zaki, M.M.; Youssef, A.M.; ElDorry, G.F.; Elalfy, D.Y. Study of plasma amino acid levels in children with autism: An Egyptian sample. Egypt. J. Med. Hum. Genet. 2014, 15, 181–186. [Google Scholar] [CrossRef] [Green Version]
- Tu, W.J.; Chen, H.; He, J. Application of LC-MS/MS analysis of plasma amino acids profiles in children with autism. J. Clin. Biochem. Nutr. 2012, 51, 248–249. [Google Scholar] [CrossRef] [Green Version]
- Saleem, T.H.; Shehata, G.A.; Toghan, R.; Sakhr, H.M.; Bakri, A.H.; Desoky, T.; Hamdan, F.R.A.; Mohamed, N.F.; Hassan, M.H. Assessments of amino acids, ammonia and oxidative stress among cohort of Egyptian autistic children: Correlations with electroencephalogram and disease severity. Neuropsychiatr. Dis. Treat. 2020, 16, 11–24. [Google Scholar] [CrossRef] [Green Version]
- Boccuto, L.; Chen, C.-F.; Pittman, A.R.; Skinner, C.D.; McCartney, H.J.; Jones, K.; Bochner, B.R.; Stevenson, R.E.; Schwartz, C.E. Decreased tryptophan metabolism in patients with autism spectrum disorders. Mol. Autism 2013, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Gevi, F.; Zolla, L.; Gabriele, S.; Persico, A.M. Urinary Metabolomics of Young Italian Autistic Children Supports Abnormal Tryptophan and Purine Metabolism. Mol. Autism 2016, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Naushad, S.M.; Jain, J.M.; Prasad, C.K.; Naik, U.; Akella, R.R. Autistic children exhibit distinct plasma amino acid profile. Indian J. Biochem. Biophys. 2013, 50, 474–478. [Google Scholar] [PubMed]
- Brister, D.; Rose, S.; Delhey, L.; Tippett, M.; Jin, Y.; Gu, H.; Frye, R.E. Metabolomic Signatures of Autism Spectrum Disorder. J. Pers. Med. 2022, 12, 1727. [Google Scholar] [CrossRef]
- Kelly, R.S.; Boulin, A.; Laranjo, N.; Lee-Sarwar, K.; Chu, S.H.; Yadama, A.P.; Carey, V.; Litonjua, A.A.; Lasky-Su, J.; Weiss, S.T. Metabolomics and Communication Skills Development in Children; Evidence from the Ages and Stages Questionnaire. Metabolites 2019, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Tărlungeanu, D.C.; Deliu, E.; Dotter, C.P.; Kara, M.; Janiesch, P.C.; Scalise, M.; Galluccio, M.; Tesulov, M.; Morelli, E.; Sonmez, F.M.; et al. Impaired amino acid transport at the blood brain barrier is a cause of autism spectrum disorder. Cell 2016, 167, 1481–1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascio, L.; Chen, C.; Pauly, R.; Srikanth, S.; Jones, K.; Skinner, C.D.; Stevenson, R.E.; Schwartz, C.E.; Boccuto, L. Abnormalities in the genes that encode Large Amino Acid Transporters increase the risk of Autism Spectrum Disorder. Mol. Genet. Genomic. Med. 2020, 8, e1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aloisi, E.; Le Corf, K.; Dupuis, J.; Zhang, P.; Ginger, M.; Labrousse, V.; Spatuzza, M.; Georg Haberl, M.; Costa, L.; Shigemoto, R.; et al. Altered surface mGluR5 dynamics provoke synaptic NMDAR dysfunction and cognitive defects in Fmr1 knockout mice. Nat. Commun. 2017, 8, 1103. [Google Scholar] [CrossRef] [Green Version]
- D’Antoni, S.; Spatuzza, M.; Bonaccorso, C.M.; Musumeci, S.A.; Ciranna, L.; Nicoletti, F.; Huber, K.M.; Catania, M.V. Dysregulation of group-I metabotropic glutamate (mGlu) receptor mediated signalling in disorders associated with Intellectual Disability and Autism. Neurosci. Biobehav. Rev. 2014, 46, 228–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Z.; Zhu, T.; Qu, Y.; Mu, D. Blood Glutamate Levels in Autism Spectrum Disorder: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0158688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Shi, X.-J.; Liu, H.; Mao, X.; Gui, L.-N.; Wang, H.; Cheng, Y. Oxidative stress marker aberrations in children with autism spectrum disorder: A systematic review and meta-analysis of 87 studies (N = 9109). Transl. Psychiatry 2021, 11, 15. [Google Scholar] [CrossRef]
- Delhey, L.M.; Tippett, M.; Rose, S.; Bennuri, S.C.; Slattery, J.C.; Melnyk, S.; James, S.J.; Frye, R.E. Comparison of Treatment for Metabolic Disorders Associated with Autism:Reanalysis of Three Clinical Trials. Front. Neurosci. 2018, 12, 19. [Google Scholar] [CrossRef]
- Aldred, S.; Moore, K.M.; Fitzgerald, M.; Waring, R.H. Plasma amino acid levels in children with autism and their families. J. Autism Dev. Disord. 2003, 33, 93–97. [Google Scholar] [CrossRef]
- Smith, A.M.; Natowicz, M.R.; Braas, D.; Ludwig, M.A.; Ney, D.M.; Donley, E.L.R.; Burrier, R.E.; Amaral, D.G. A Metabolomics Approach to Screening for Autism Risk in the Children's Autism Metabolome Project. Autism Res. 2020, 13, 1270–1285. [Google Scholar] [CrossRef]
- Novarino, G.; El-Fishawy, P.; Kayserili, H.; Meguid, N.A.; Scott, E.M.; Schroth, J.; Silhavy, J.L.; Kara, M.; Khalil, R.O.; Ben-Omran, T.; et al. Mutations in BCKD-kinase lead to a potentially treatable form of autism with epilepsy. Science 2012, 338, 394–397. [Google Scholar] [CrossRef] [Green Version]
- García-Cazorla, A.; Oyarzabal, A.; Fort, J.; Robles, C.; Castejón, E.; Ruiz-Sala, P.; Bodoy, S.; Merinero, B.; Lopez-Sala, A.; Dopazo, J.; et al. Two novel mutations in the BCKDK (branched-chain keto-acid dehydrogenase kinase) gene are responsible for a neurobehavioral deficit in two pediatric unrelated patients. Hum. Mutat. 2014, 35, 470–477. [Google Scholar] [CrossRef]
- Du, C.; Liu, W.J.; Yang, J.; Zhao, S.S.; Liu, H.X. The Role of Branched-Chain Amino Acids and Branched-Chain α-Keto Acid Dehydrogenase Kinase in Metabolic Disorders. Front. Nutr. 2022, 9, 932670. [Google Scholar] [CrossRef]
- Barone, R.; Alaimo, S.; Messina, M.; Pulvirenti, A.; Bastin, J.; Ferro, A.; Frye, R.E.; Rizzo, R.; MIMIC-Autism group. A Subset of Patients With Autism Spectrum Disorders Show a Distinctive Metabolic Profile by Dried Blood Spot Analyses. Front. Psychiatry 2018, 9, 636. [Google Scholar] [CrossRef] [Green Version]
- Waisbren, S.E.; Members of the Urea Cycle Disorders Consortium; Cuthbertson, D.; Burgard, P.; Holbert, A.; McCarter, R.; Cederbaum, S. Biochemical markers and neuropsychological functioning in distal urea cycle disorders. J. Inherit. Metab. Dis. 2018, 41, 657–667. [Google Scholar] [CrossRef]


{kind=link}
| Amino Acid | ASD (μmol/L) | ID (μmol/L) | TD (μmol/L) | ANOVA | Bonferroni Correction | ||
|---|---|---|---|---|---|---|---|
| ASD vs. TD | ASD vs. ID | TD vs. ID | |||||
| Ala | 238.8 ± 69.3 | 236.9 ± 73.7 | 269.41 ± 91.5 | F = 1.522 p = 0.224 η2p = 0.03 | p = 0.354 | p = 1.000 | p = 0.45 |
| Arg | 22.25 ± 12.3 | 16.81 ± 7.6 | 18.75 ± 12.9 | F = 1.751 p = 0.180 η2p = 0.04 | p = 0.678 | p = 0.238 | p = 1 |
| Cit | 22.52 ± 4.8 | 22.3 ± 4.9 | 21.8 ± 5.8 | F = 0.177 p = 0.838 η2p = 0.004 | p = 1 | p = 1 | p = 1 |
| Gln | 458.2 ± 109.4 | 459.6 ± 97.9 | 450.5 ± 122.8 | F = 0.052 p = 0.950 η2p = 0.001 | p = 1 | p = 1 | p = 1 |
| Glu | 86.8 ± 37.3 | 85 ± 26 | 87.7 ± 26.2 | F = 0.045 p = 0.956 η2p = 0.001 | p = 1 | p = 1 | p = 1 |
| Gly | 226.3 ± 66.2 | 222.4 ± 50 | 247.2 ± 99.1 | F = 0.861 p = 0.426 η2p = 0.02 | p = 0.787 | p = 1 | p = 0.75 |
| Leu | 125.2 ± 25.5 | 118.4 ± 22.1 | 131.4 ± 32.4 | F = 1.395 p = 0.253 η2p = 0.03 | p = 1 | p = 1 | p = 0.253 |
| Met | 11.4 ± 4.4 | 11 ± 2.9 | 12.3 ± 3.5 | F = 0.741 p = 0.480 η2p = 0.02 | p = 1 | p = 1 | p = 0.721 |
| Orn | 71.4 ± 17.3 | 78.2 ± 13.6 | 99.9 ± 61.2 | F = 5.148 * p = 0.008 * η2p = 0.11 | p = 0.006 * | p = 1 | p = 0.119 |
| Phe | 39.6 ± 7.8 | 40.5 ± 6.5 | 44.7 ± 10 | F = 3.301 * p = 0.042 * η2p = 0.07 | p = 0.043 * | p = 1 | p = 0.226 |
| Pro | 107.5 ± 37.9 | 116.6 ± 50 | 116.9 ± 46.6 | F = 0.503 p = 0.607 η2p = 0.01 | p = 1 | p = 1 | p = 1 |
| Tyr | 47.9 ± 9.3 | 53.30 ± 11.6 | 58.7 ± 21.3 | F = 4.560 * p = 0.013 * η2p = 0.1 | p = 0.010 * | p = 0.492 | p = 0.581 |
| Val | 158.1 ± 34.1 | 54.4 ± 34.5 | 155.2 ± 35.4 | F = 0.101 p = 0.904 η2p = 0.002 | p = 1 | p = 1 | p = 1 |
| Amino Acid Ratio | ASD (μmol/L) | ID (μmol/L) | TD (μmol/L) | ANOVA | Bonferroni Correction | ||
|---|---|---|---|---|---|---|---|
| ASD vs. TD | ASD vs. ID | TD vs. ID | |||||
| Leu+Val/Phe+Tyr | 3.3 ± 0.5 | 2.9 ± 0.5 | 2.8 ± 0.5 | F = 6.641 * p = 0.002 * η2p = 0.13 * | p = 0.003 * | p = 0.053 | p = 1 |
| Cit/Arg | 1.5 ± 1.3 | 1.7 ± 1.3 | 1.8 ± 1.3 | F = 0.255 p = 0.776 η2p = 0.006 | p = 1 | p = 1 | p = 1 |
| Cit/Phe | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.5 ± 0.2 | F = 2.176 p = 0.120 η2p = 0.05 | p = 0.121 | p = 1 | p = 0.702 |
| Leu/Phe | 3.2 ± 0.5 | 3 ± 0.5 | 3 ± 0.6 | F = 1.990 p = 0.143 η2p = 0.04 | p = 0.350 | p = 0.267 | p = 1 |
| Met/Leu | 0.1 ± 0.03 | 0.1 ± 0.03 | 0.1 ± 0.02 | F = 0.348 p = 0.707 η2p = 0.008 | p = 1 | p = 1 | p = 1 |
| Met/Phe | 0.3 ± 0.1 | 0.3 ± 0.1 | 0.3 ± 0.1 | F = 0.052 p = 0.950 η2p = 0.001 | p = 1 | p = 1 | p = 1 |
| Phe/Tyr | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.9 ± 0.4 | F = 0.435 p = 0.649 η2p = 0.01 | p = 1 | p = 1 | p = 1 |
| Glu/Gln | 0.2 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | F = 0.138 p = 0.871 η2p = 0.003 | p = 1 | p = 1 | p = 1 |
| Tyr/Leu | 0.4 ± 0.1 | 0.5 ± 0.1 | 0.5 ± 0.1 | F = 5.214 * p = 0.007 * η2p = 0.11 * | p = 0.044 * | p = 0.017 * | p = 1 |
| TyMet | 5 ± 2.5 | 5.2 ± 1.8 | 4.9 ± 1.4 | F = 0.090 p = 0.914 η2p = 0.002 | p = 1 | p = 1 | p = 1 |
| Val/Phe | 4+ 0.72 | 3.9 ± 0.7 | 3.6 ± 0.7 | F = 3.738 * p = 0.028 ;η2p = 0.08 * | p = 0.023 * | p = 0.927 | p = 0.473 |
| Metabolite | TD Reference Range (10th and 90th Percentile) (μmol/L) | ASD below RR (μmol/L) | ASD above RR (μmol/L) |
|---|---|---|---|
| Ala | 206.00–401.50 | 35% | 0% |
| Arg | 4.01–34.30 | 7.5% | 17.5% |
| Cit | 14.80–29.30 | 7.5% | 2.5% |
| Gln | 283.00–596.00 | 5% | 12.5% |
| Glu | 62.50–130.00 | 12.5% | 7.5% |
| Gly | 122.00–350.50 | 7.5% | 7.5% |
| Leu | 91.00–172.00 | 5% | 5% |
| Met | 8.61–16.45 | 22.5% | 10% |
| Orn | 56.70–141.50 | 15% | 0% |
| Phe | 33.40–60.55 | 17.5% | 2.5% |
| Pro | 59.40–164.50 | 5% | 10% |
| Tyr | 40.20–86.15 | 20% | 0% |
| Val | 117.00–210.50 | 7.5% | 12.5% |
| Leu+Val/Phe+Tyr | 2.17–3.41 | 0% | 30% |
| Cit/Arg | 0.69–2.90 | 17.5% | 7.5% |
| Cit/Phe | 0.35–0.69 | 2.5% | 22.5% |
| Leu/Phe | 2.58–3.88 | 7.5% | 5% |
| Met/Leu | 0.07–0.13 | 20% | 7.5% |
| Met/Phe | 0.18–0.37 | 17.5% | 17.5% |
| Phe/Tyr | 0.54–0.98 | 0% | 15% |
| Glu/Gln | 0.13–0-28 | 7.5% | 12.5% |
| Tyr/Leu | 0.32–0-58 | 10% | 0% |
| Tyr/Met | 3.34–6.94 | 20% | 12.5% |
| Val/Phe | 2.74–4.43 | 5% | 35% |
| Metabolite | IQ/DQ | ADOS-2 SA | ADOS-2 RRB | ADOS-2 TS |
|---|---|---|---|---|
| Ala | r = 0.0034 | r = −0.3080 | r = −0.1343 | r = −0.2980 |
| p = 0.9839 | p = 0.0812 | p = 0.4562 | p = 0.0921 | |
| Arg | r = 0.0452 | r = 0.0467 | r = 0.2065 | r = 0.1075 |
| p = 0.7904 | p = 0.7965 | p = 0.2490 | p = 0.5514 | |
| Cit | r = 0.0196 | r = 0.0825 | r = 0.4801 * | r = 0.2287 |
| p = 0.9082 | p = 0.6479 | p = 0.0047 * | p = 0.2005 | |
| Gln | r = −0.0644 | r = 0.2175 | r = 0.0066 | r = 0.0066 |
| p = 0.7048 | p = 0.2241 | p = 0.9708 | p = 0.9708 | |
| Glu | r = −0.1589 | r = 0.1285 | r = −0.1926 | r = 0.0420 |
| p = 0.3476 | p = 0.4762 | p = 0.2829 | p = 0.0.8209 | |
| Gly | r = 0.1389 | r = 0.0153 | r = 0.0503 | r = 0.0294 |
| p = 0.4124 | p = 0.9327 | p = 0.7810 | p = 0.8709 | |
| Leu | r = 0.1676 | r = 0.0500 | r = −0.0645 | r = 0.0195 |
| p = 0.3213 | p = 0.7823 | p = 0.7215 | p = 0.9144 | |
| Met | r = −0.0063 | r = −0.0574 | r = 0.2053 | r = 0.0209 |
| p = 0.9707 | p = 0.7470 | p = 0.2516 | p = 0.9081 | |
| Orn | r = −0.01176 | r = 0.0021 | r = 0.2628 | r = 0.0898 |
| p = 0.4881 | p = 0.9908 | p = 0.1395 | p = 0.6192 | |
| Phe | r = −0.1359 | r = −0.0420 | r = 0.1144 | r = 0.0038 |
| p = 0.4227 | p = 0.8164 | p = 0.5261 | p = 0.9831 | |
| Pro | r = −0.0830 | r = 0.0251 | r = 0.1149 | r = 0.0591 |
| p = 0.6251 | p = 0.8896 | p = 0.05244 | p = 0.7437 | |
| Tyr | r = 0.1805 | r = −0.0565 | r = 0.1749 | r = 0.0122 |
| p = 0.2850 | p = 0.7548 | p = 0.3303 | p = 0.9463 | |
| Val | r = 0.0514 | r = −0.1138 | r = 0.0639 | r = −0.0721 |
| p = 0.7627 | p = 0.5283 | p = 0.7238 | p = 0.6902 | |
| Leu+Val/Phe+Tyr | r = 0.0694 | r = 0.0050 | r = −0.2094 | r = −0.0661 |
| p = 0.6830 | p = 0.9780 | p = 0.2421 | p = 0.7148 | |
| Cit/Arg | r = −0.1241 | r = 0.0429 | r = −0.1872 | r = −0.0275 |
| p = 0.4643 | p = 0.8124 | p = 0.2969 | p = 0.8794 | |
| Cit/Phe | r = 0.1019 | r = 0.1080 | r = 0.3341 | r = 0.2007 |
| p = 0.5482 | p = 0.5498 | p = 0.0574 | p = 0.2628 | |
| Leu/Phe | r = 0.2826 | r = 0.1491 | r = −0.1977 | r = 0.0562 |
| p = 0.0901 | p = 0.4077 | p = 0.2701 | p = 0.7651 | |
| Met/Leu | r = −0.1353 | r = −0.0700 | r = 0.2952 | r = 0.0415 |
| p = 0.4245 | p = 0.6988 | p = 0.0953 | p = 0.8188 | |
| Met/Phe | r = 0.0718 | r = −0.0526 | r = 0.2172 | r = 0.0296 |
| p = 0.6727 | p = 0.7711 | p = 0.2246 | p = 0.8703 | |
| Phe/Tyr | r = −0.2990 | r = 0.0003 | r = −0.1152 | r = −0.0518 |
| p = 0.0722 | p = 0.9987 | p = 0.3885 | p = 0.7748 | |
| Glu/Gln | r = −0.1186 | r = 0.1717 | r = −0.2950 | r = 0.0421 |
| p = 0.4843 | p = 0.3395 | p = 0.0956 | p = 0.8159 | |
| Tyr/Leu | r = 0.0171 | r = −0.0933 | r = 0.2267 | r = −0.0006 |
| p = 0.9200 | p = 0.6057 | p = 0.2046 | p = 0.9972 | |
| Tyr/Met | r = 0.0165 | r = 0.0902 | r = −0.1493 | r = 0.0240 |
| p = 0.9226 | p = 0.6178 | p = −0.4070 | p = 0.8944 | |
| Val/Phe | r = 0.1407 | r = −0.0887 | r = −0.0674 | r = −0.0955 |
| p = 0.4061 | p = 0.6234 | p = 0.7092 | p = 0.5971 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Randazzo, M.; Prato, A.; Messina, M.; Meli, C.; Casabona, A.; Rizzo, R.; Barone, R. Neuroactive Amino Acid Profile in Autism Spectrum Disorder: Results from a Clinical Sample. Children 2023, 10, 412. https://doi.org/10.3390/children10020412
Randazzo M, Prato A, Messina M, Meli C, Casabona A, Rizzo R, Barone R. Neuroactive Amino Acid Profile in Autism Spectrum Disorder: Results from a Clinical Sample. Children. 2023; 10(2):412. https://doi.org/10.3390/children10020412
Chicago/Turabian StyleRandazzo, Martina, Adriana Prato, MariaAnna Messina, Concetta Meli, Antonino Casabona, Renata Rizzo, and Rita Barone. 2023. "Neuroactive Amino Acid Profile in Autism Spectrum Disorder: Results from a Clinical Sample" Children 10, no. 2: 412. https://doi.org/10.3390/children10020412
APA StyleRandazzo, M., Prato, A., Messina, M., Meli, C., Casabona, A., Rizzo, R., & Barone, R. (2023). Neuroactive Amino Acid Profile in Autism Spectrum Disorder: Results from a Clinical Sample. Children, 10(2), 412. https://doi.org/10.3390/children10020412