
Perinatal Inflammation: Could Partial Blocking of Cell Adhesion Molecule Function Be a Solution?

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
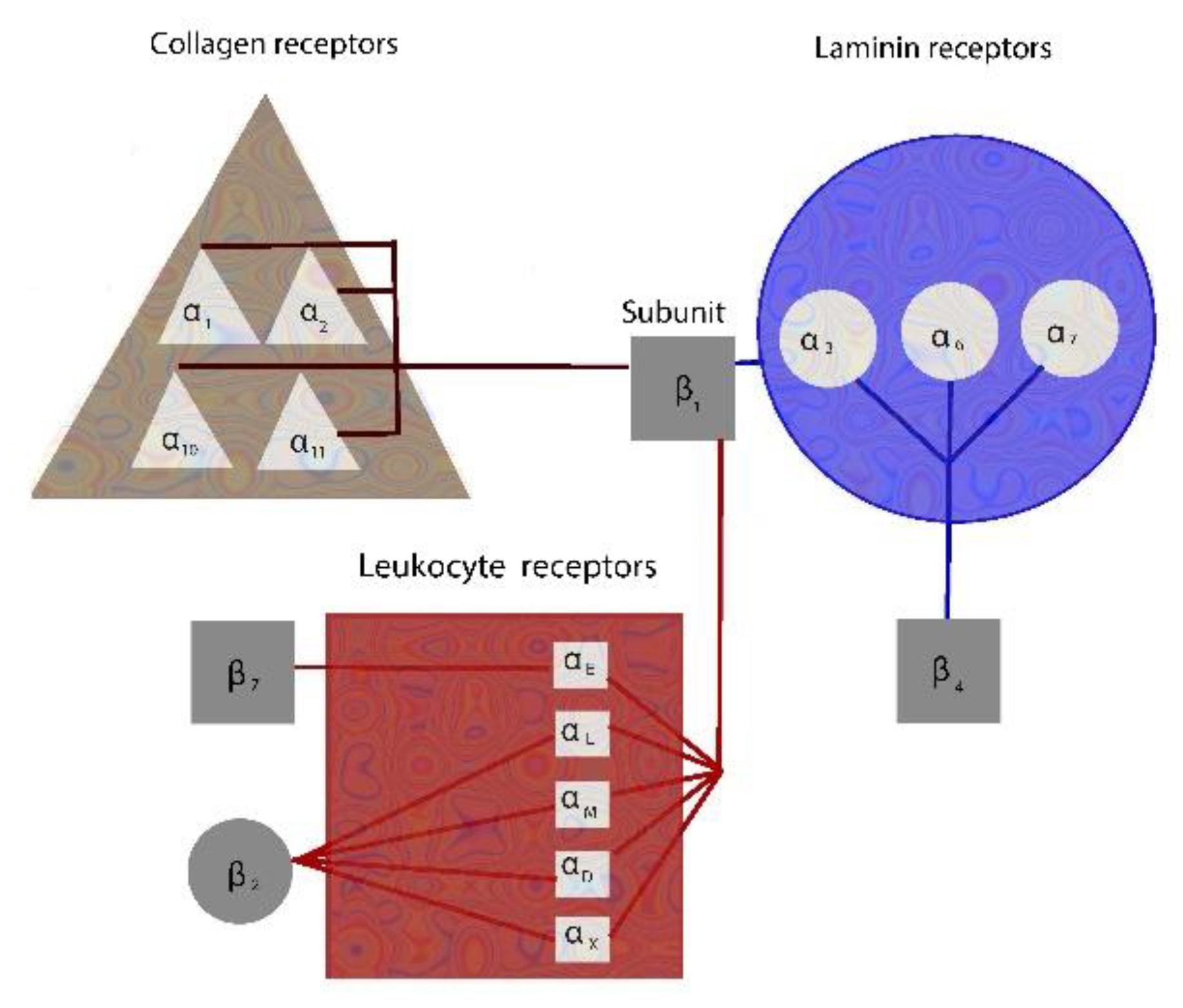
2. Integrins Mediate Cell-Cell and Cell-Matrix Interactions
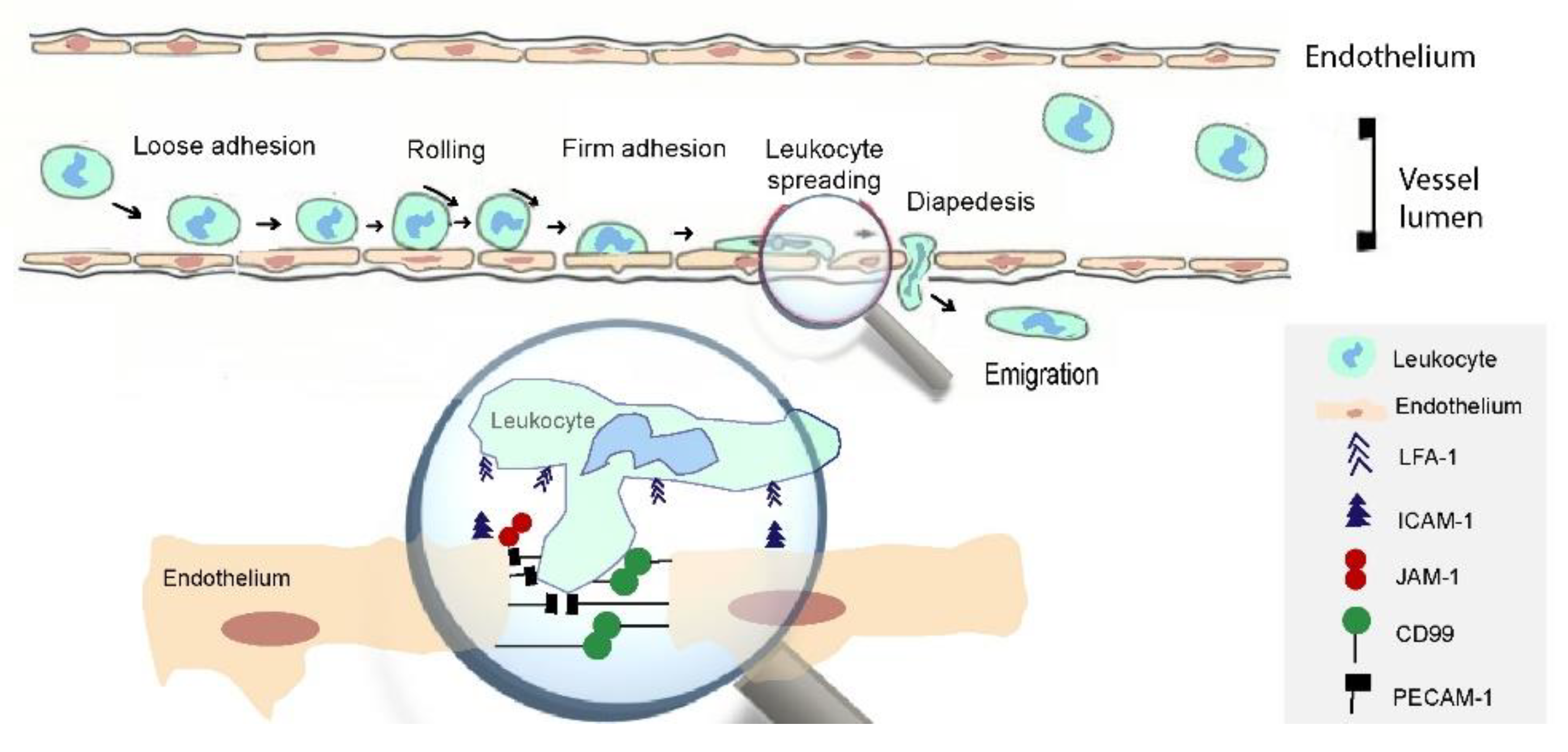
3. Selectins Mediate Leukocyte Interactions with Endothelium and Platelets
4. Role of Immunoglobulin Gene (Ig) Super Family in Leukocyte Adherence
5. Integrin Function in the Perinatal Period
- (a)
- β-2 subfamily integrins
- (b)
- β-1 and β-3 subfamily integrins
6. Selectin Function in the Perinatal Period
7. Ig Superfamily Function in the Perinatal Period
8. Genetic Mutations Result in Leukocyte Adhesion Deficiency
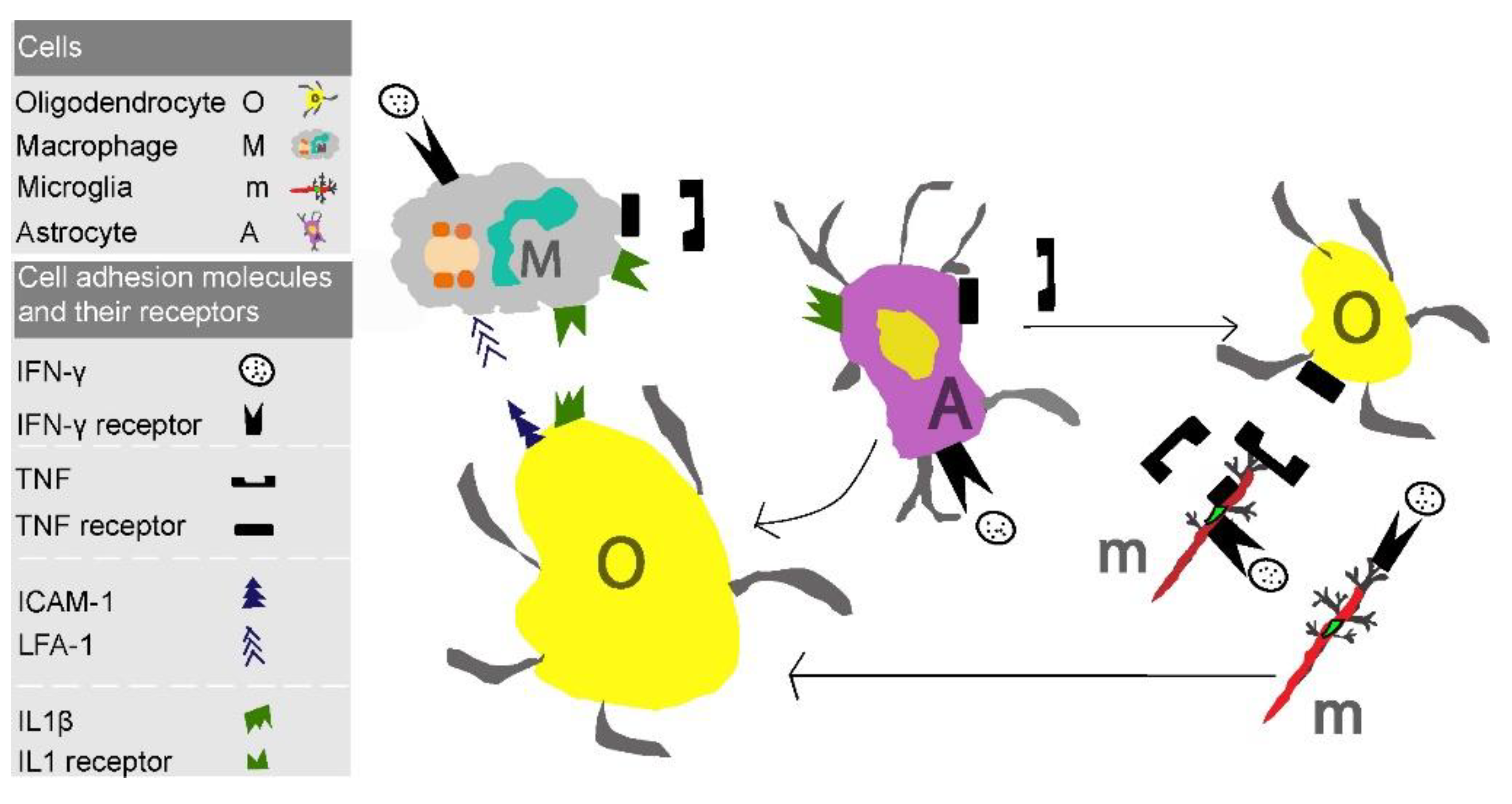
9. Future Perspectives
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Vrachnis, N.; Vitoratos, N.; Iliodromiti, Z.; Deligeoroglou, E.; Creatsas, G. Intrauterine inflammation and preterm delivery. Ann. N. Y. Acad. Sci. 2010, 1205, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Iliodromiti, Z.; Zygouris, D.; Sifakis, S.; Pappa, K.I.; Tsikouras, P.; Salakos, N.; Daniilidis, A.; Siristadidis, C.; Vrachnis, N. Acute lung injury in preterm fetuses and neonates: Mechanisms and molecular pathways. J. Matern. Fetal Neonatal Med. 2013, 26, 1696–1704. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.C.; Hughes, B.J.; Smith, C.W. Abnormal mobility of neonatal polymorphonuclear leukocytes. Relationship to impaired redistribution of surface adhesion sites by chemotactic factor or colchicine. J. Clin. Investig. 1981, 68, 863–874. [Google Scholar] [CrossRef] [Green Version]
- Iliodromiti, Z.; Iliodromiti, Z.; Anastasiadis, A.; Varras, M.; Pappa, K.I.; Siristatidis, C.; Bakoulas, V.; Mastorakos, G.; Vrachnis, N. Monocyte function in the fetus and the preterm neonate: Immaturity combined with functional impairment. Mediat. Inflamm. 2013, 2013, 753752. [Google Scholar] [CrossRef] [Green Version]
- Carr, R. Neutrophil production and function in newborn infants. Br. J. Haematol. 2000, 110, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Vrachnis, N.; Malamitsi-Puchner, A.; Samoli, E.; Botsis, D.; Iliodromiti, Z.; Baka, S.; Hassiakos, D.; Creatsas, G. Elevated mid-trimester amniotic fluid ADAM-8 concentrations as a potential risk factor for preterm delivery. J. Soc. Gynecol. Investig. 2006, 13, 186–190. [Google Scholar] [CrossRef]
- Malamitsi-Puchner, A.; Vrachnis, N.; Samoli, E.; Baka, S.; Alexandrakis, G.; Puchner, K.P.; Iliodromiti, Z.; Hassiakos, D. Investigation of midtrimester amniotic fluid factors as potential predictors of term and preterm deliveries. Mediat. Inflamm. 2006, 2006, 94381. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.W. 3. Adhesion molecules and receptors. J. Allergy Clin. Immunol. 2008, 121 (Suppl. 2), S375–S379; quiz S414. [Google Scholar] [CrossRef]
- Zarbock, A.; Ley, K. Neutrophil adhesion and activation under flow. Microcirculation 2009, 16, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Giancotti, F.G.; Ruoslahti, E. Integrin signaling. Science 1999, 285, 1028–1032. [Google Scholar] [CrossRef]
- Mokhtar, D.M.; Abdelhafez, E.A. An overview of the structural and functional aspects of immune cells in teleosts. Histol. Histopathol. 2021, 1, 18302. [Google Scholar] [CrossRef]
- Sperandio, M. Selectins and glycosyltransferases in leukocyte rolling in vivo. FEBS J. 2006, 273, 4377–4389. [Google Scholar] [CrossRef] [PubMed]
- Landgraf, M.A.; Martinez, L.L.; Rastelli, V.M.; Franco Mdo, C.; Soto-Suazo, M.; Tostes Rde, C.; Carvalho, M.H.; Nigro, D.; Fortes, Z.B. Intrauterine undernutrition in rats interferes with leukocyte migration, decreasing adhesion molecule expression in leukocytes and endothelial cells. J. Nutr. 2005, 135, 1480–1485. [Google Scholar] [CrossRef] [Green Version]
- Leeuwenberg, J.F.; Smeets, E.F.; Neefjes, J.J.; Shaffer, M.A.; Cinek, T.; Jeunhomme, T.M.; Ahern, T.J.; Buurman, W.A. E-selectin and intercellular adhesion molecule-1 are released by activated human endothelial cells in vitro. Immunology 1992, 77, 543–549. [Google Scholar] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueras-Aloy, J.; Gomez-Lopez, L.; Rodriguez-Miguelez, J.M.; Salvia-Roiges, M.D.; Jordan-Garcia, I.; Ferrer-Codina, I.; Carbonell-Estrany, X.; Jimenez-Gonzalez, R. Serum soluble ICAM-1, VCAM-1, L-selectin, and P-selectin levels as markers of infection and their relation to clinical severity in neonatal sepsis. Am. J. Perinatol. 2007, 24, 331–338. [Google Scholar] [CrossRef]
- Vrachnis, N.; Karavolos, S.; Iliodromiti, Z.; Sifakis, S.; Siristatidis, C.; Mastorakos, G.; Creatsas, G. Review: Impact of mediators present in amniotic fluid on preterm labour. In Vivo 2012, 26, 799–812. [Google Scholar]
- Docheva, N.; Romero, R.; Chaemsaithong, P.; Tarca, A.L.; Bhatti, G.; Pacora, P.; Panaitescu, B.; Chaiyasit, N.; Chaiworapongsa, T.; Maymon, E.; et al. The profiles of soluble adhesion molecules in the “great obstetrical syndromes”. J. Matern. Fetal Neonatal Med. 2019, 32, 2113–2136. [Google Scholar] [CrossRef]
- Coata, G.; Pennacchi, L.; Bini, V.; Liotta, L.; Di Renzo, G.C. Soluble adhesion molecules: Marker of pre-eclampsia and intrauterine growth restriction. J. Matern. Fetal Neonatal Med. 2002, 12, 28–34. [Google Scholar] [CrossRef]
- Huang, M.T.; Larbi, K.Y.; Scheiermann, C.; Woodfin, A.; Gerwin, N.; Haskard, D.O.; Nourshargh, S. ICAM-2 mediates neutrophil transmigration in vivo: Evidence for stimulus specificity and a role in PECAM-1-independent transmigration. Blood 2006, 107, 4721–4727. [Google Scholar] [CrossRef]
- Woodfin, A.; Voisin, M.B.; Nourshargh, S. PECAM-1: A multi-functional molecule in inflammation and vascular biology. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2514–2523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malamitsi-Puchner, A.; Vrachnis, N.; Samoli, E.; Baka, S.; Iliodromiti, Z.; Puchner, K.P.; Malligianis, P.; Hassiakos, D. Possible early prediction of preterm birth by determination of novel proinflammatory factors in midtrimester amniotic fluid. Ann. N. Y. Acad. Sci. 2006, 1092, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Vestweber, D.; Winderlich, M.; Cagna, G.; Nottebaum, A.F. Cell adhesion dynamics at endothelial junctions: VE-cadherin as a major player. Trends Cell Biol. 2009, 19, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.Y.; Harlos, K.; Bottomley, M.J.; Robinson, R.C.; Driscoll, P.C.; Edwards, R.M.; Clements, J.M.; Dudgeon, T.J.; Stuart, D.I. Crystal structure of an integrin-binding fragment of vascular cell adhesion molecule-1 at 1.8 A resolution. Nature 1995, 373, 539–544. [Google Scholar] [CrossRef]
- Park, E.J.; Myint, P.K.; Ito, A.; Appiah, M.G.; Darkwah, S.; Kawamoto, E.; Shimaoka, M. Integrin-Ligand Interactions in Inflammation, Cancer, and Metabolic Disease: Insights into the Multifaceted Roles of an Emerging Ligand Irisin. Front. Cell Dev. Biol. 2020, 8, 588066. [Google Scholar] [CrossRef]
- Cook-Mills, J.M. VCAM-1 signals during lymphocyte migration: Role of reactive oxygen species. Mol. Immunol. 2002, 39, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Privratsky, J.R.; Newman, D.K.; Newman, P.J. PECAM-1: Conflicts of interest in inflammation. Life Sci. 2010, 87, 69–82. [Google Scholar] [CrossRef] [Green Version]
- Raymond, S.L.; Mathias, B.J.; Murphy, T.J.; Rincon, J.C.; Lopez, M.C.; Ungaro, R.; Ellett, F.; Jorgensen, J.; Wynn, J.L.; Baker, H.V.; et al. Neutrophil chemotaxis and transcriptomics in term and preterm neonates. Transl. Res. 2017, 190, 4–15. [Google Scholar] [CrossRef]
- Anderson, D.C.; Rothlein, R.; Marlin, S.D.; Krater, S.S.; Smith, C.W. Impaired transendothelial migration by neonatal neutrophils: Abnormalities of Mac-1 (CD11b/CD18)-dependent adherence reactions. Blood 1990, 76, 2613–2621. [Google Scholar] [CrossRef] [Green Version]
- Torok, C.; Lundahl, J.; Hed, J.; Lagercrantz, H. Diversity in regulation of adhesion molecules (Mac-1 and L-selectin) in monocytes and neutrophils from neonates and adults. Arch. Dis. Child. 1993, 68, 561–565. [Google Scholar] [CrossRef]
- Abughali, N.; Berger, M.; Tosi, M.F. Deficient total cell content of CR3 (CD11b) in neonatal neutrophils. Blood 1994, 83, 1086–1092. [Google Scholar] [CrossRef] [Green Version]
- Nupponen, I.; Pesonen, E.; Andersson, S.; Makela, A.; Turunen, R.; Kautiainen, H.; Repo, H. Neutrophil activation in preterm infants who have respiratory distress syndrome. Pediatrics 2002, 110 (1 Pt 1), 36–41. [Google Scholar] [CrossRef]
- Storm, S.W.; Mariscalco, M.M.; Tosi, M.F. Postnatal maturation of total cell content and up-regulated surface expression of Mac-1 (CD11b/CD18) in polymorphonuclear leukocytes of human infants. J. Leukoc. Biol. 2008, 84, 477–479. [Google Scholar] [CrossRef] [PubMed]
- Weinschenk, N.P.; Farina, A.; Bianchi, D.W. Premature infants respond to early-onset and late-onset sepsis with leukocyte activation. J. Pediatr. 2000, 137, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Sarafidis, K.; Drossou-Agakidou, V.; Kanakoudi-Tsakalidou, F.; Taparkou, A.; Tsakalidis, C.; Tsandali, C.; Kremenopoulos, G. Evidence of early systemic activation and transendothelial migration of neutrophils in neonates with severe respiratory distress syndrome. Pediatr. Pulmonol. 2001, 31, 214–219. [Google Scholar] [CrossRef]
- McEvoy, L.T.; Zakem-Cloud, H.; Tosi, M.F. Total cell content of CR3 (CD11b/CD18) and LFA-1 (CD11a/CD18) in neonatal neutrophils: Relationship to gestational age. Blood 1996, 87, 3929–3933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buschmann, K.; Tschada, R.; Metzger, M.S.; Braach, N.; Kuss, N.; Hudalla, H.; Poeschl, J.; Frommhold, D. RAGE controls leukocyte adhesion in preterm and term infants. BMC Immunol. 2014, 15, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, A.L.; El-Bjeirami, W.; West, J.L.; McIntire, L.V.; Smith, C.W. Transendothelial migration enhances integrin-dependent human neutrophil chemokinesis. J. Leukoc. Biol. 2007, 81, 686–695. [Google Scholar] [CrossRef] [PubMed]
- Douglass, S.; Goyal, A.; Iozzo, R.V. The role of perlecan and endorepellin in the control of tumor angiogenesis and endothelial cell autophagy. Connect. Tissue Res. 2015, 56, 381–391. [Google Scholar] [CrossRef] [Green Version]
- O’Gorman, M.R.; Millard, D.D.; Lowder, J.N.; Yogev, R. Lymphocyte subpopulations in healthy 1-3-day-old infants. Cytometry 1998, 34, 235–241. [Google Scholar] [CrossRef]
- Pilarski, L.M.; Yacyshyn, B.R.; Jensen, G.S.; Pruski, E.; Pabst, H.F. Beta 1 integrin (CD29) expression on human postnatal T cell subsets defined by selective CD45 isoform expression. J. Immunol. 1991, 147, 830–837. [Google Scholar] [PubMed]
- Sato, K.; Kawasaki, H.; Nagayama, H.; Enomoto, M.; Morimoto, C.; Tadokoro, K.; Juji, T.; Takahashi, T. Chemokine receptor expressions and responsiveness of cord blood T cells. J. Immunol. 2001, 166, 1659–1666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenig, J.M.; Stegner, J.J.; Schmeck, A.C.; Saxonhouse, M.A.; Kenigsberg, L.E. Neonatal neutrophils with prolonged survival exhibit enhanced inflammatory and cytotoxic responsiveness. Pediatr. Res. 2005, 57, 424–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.K.; Keeney, S.E.; Alpard, S.K.; Schmalstieg, F.C. Comparison of L-selectin and CD11b on neutrophils of adults and neonates during the first month of life. Pediatr. Res. 2003, 53, 132–136. [Google Scholar] [CrossRef]
- Sundqvist, M.; Osla, V.; Jacobsson, B.; Rudin, A.; Savman, K.; Karlsson, A. Cord blood neutrophils display a galectin-3 responsive phenotype accentuated by vaginal delivery. BMC Pediatr. 2013, 13, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, S.M.; Corriden, R.; Nizet, V. Age-Appropriate Functions and Dysfunctions of the Neonatal Neutrophil. Front. Pediatr. 2017, 5, 23. [Google Scholar] [CrossRef]
- Buhrer, C.; Graulich, J.; Stibenz, D.; Dudenhausen, J.W.; Obladen, M. L-selectin is down-regulated in umbilical cord blood granulocytes and monocytes of newborn infants with acute bacterial infection. Pediatr. Res. 1994, 36, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Mariscalco, M.M.; Vergara, W.; Mei, J.; Smith, E.O.; Smith, C.W. Mechanisms of decreased leukocyte localization in the developing host. Am. J. Physiol. Heart Circ. Physiol. 2002, 282, H636–H644. [Google Scholar] [CrossRef] [Green Version]
- Orwoll, B.E.; Sapru, A. Biomarkers in Pediatric ARDS: Future Directions. Front. Pediatr. 2016, 4, 55. [Google Scholar] [CrossRef] [Green Version]
- Rajasekhar, D.; Kestin, A.S.; Bednarek, F.J.; Ellis, P.A.; Barnard, M.R.; Michelson, A.D. Neonatal platelets are less reactive than adult platelets to physiological agonists in whole blood. Thromb. Haemost. 1994, 72, 957–963. [Google Scholar] [CrossRef]
- Olutoye, O.O.; Zhu, X.; Cass, D.L.; Smith, C.W. Neutrophil recruitment by fetal porcine endothelial cells: Implications in scarless fetal wound healing. Pediatr. Res. 2005, 58, 1290–1294. [Google Scholar] [CrossRef] [Green Version]
- Nussbaum, C.; Gloning, A.; Pruenster, M.; Frommhold, D.; Bierschenk, S.; Genzel-Boroviczeny, O.; von Andrian, U.H.; Quackenbush, E.; Sperandio, M. Neutrophil and endothelial adhesive function during human fetal ontogeny. J. Leukoc. Biol. 2013, 93, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, P.L.; O’Brian Smith, E.; Hegemier, S.; Welty, S.E. Early clinical markers for the development of bronchopulmonary dysplasia: Soluble E-Selectin and ICAM-1. Pediatrics 1998, 102 Pt 1, 927–932. [Google Scholar] [CrossRef]
- Brou, L.; Almli, L.M.; Pearce, B.D.; Bhat, G.; Drobek, C.O.; Fortunato, S.; Menon, R. Dysregulated biomarkers induce distinct pathways in preterm birth. BJOG 2012, 119, 458–473. [Google Scholar] [CrossRef]
- Holm, M.; Morken, T.S.; Fichorova, R.N.; VanderVeen, D.K.; Allred, E.N.; Dammann, O.; Leviton, A. Neonatology Elgan Study Ophthalmology Committees. Systemic Inflammation-Associated Proteins and Retinopathy of Prematurity in Infants Born Before the 28th Week of Gestation. Investig. Ophthalmol. Vis. Sci. 2017, 58, 6419–6428. [Google Scholar] [CrossRef] [PubMed]
- Leviton, A.; Allred, E.N.; Fichorova, R.N.; Kuban, K.C.; Michael O’Shea, T.; Dammann, O.; Elgan Study Investigators. Systemic inflammation on postnatal days 21 and 28 and indicators of brain dysfunction 2 years later among children born before the 28th week of gestation. Early Hum. Dev. 2016, 93, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahni, M.; Yeboah, B.; Das, P.; Shah, D.; Ponnalagu, D.; Singh, H.; Nelin, L.D.; Bhandari, V. Novel biomarkers of bronchopulmonary dysplasia and bronchopulmonary dysplasia-associated pulmonary hypertension. J. Perinatol. 2020, 40, 1634–1643. [Google Scholar] [CrossRef] [PubMed]
- Huseynova, S.; Panakhova, N.; Orujova, P.; Hasanov, S.; Guliyev, M.; Orujov, A. Elevated levels of serum sICAM-1 in asphyxiated low birth weight newborns. Sci. Rep. 2014, 4, 6850. [Google Scholar] [CrossRef] [Green Version]
- Coalson, J.J.; Winter, V.T.; Siler-Khodr, T.; Yoder, B.A. Neonatal chronic lung disease in extremely immature baboons. Am. J. Respir. Crit. Care Med. 1999, 160, 1333–1346. [Google Scholar] [CrossRef] [Green Version]
- Rose, M.J.; Stenger, M.R.; Joshi, M.S.; Welty, S.E.; Bauer, J.A.; Nelin, L.D. Inhaled nitric oxide decreases leukocyte trafficking in the neonatal mouse lung during exposure to >95% oxygen. Pediatr. Res. 2010, 67, 244–249. [Google Scholar] [CrossRef] [Green Version]
- Thebaud, B.; Ladha, F.; Michelakis, E.D.; Sawicka, M.; Thurston, G.; Eaton, F.; Hashimoto, K.; Harry, G.; Haromy, A.; Korbutt, G.; et al. Vascular endothelial growth factor gene therapy increases survival, promotes lung angiogenesis, and prevents alveolar damage in hyperoxia-induced lung injury: Evidence that angiogenesis participates in alveolarization. Circulation 2005, 112, 2477–2486. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.C.; Schmalsteig, F.C.; Finegold, M.J.; Hughes, B.J.; Rothlein, R.; Miller, L.J.; Kohl, S.; Tosi, M.F.; Jacobs, R.L.; Waldrop, T.C.; et al. The severe and moderate phenotypes of heritable Mac-1, LFA-1 deficiency: Their quantitative definition and relation to leukocyte dysfunction and clinical features. J. Infect. Dis. 1985, 152, 668–689. [Google Scholar] [CrossRef]
- Rivera-Matos, I.R.; Rakita, R.M.; Mariscalco, M.M.; Elder, F.F.; Dreyer, S.A.; Cleary, T.G. Leukocyte adhesion deficiency mimicking Hirschsprung disease. J. Pediatr. 1995, 127, 755–757. [Google Scholar] [CrossRef]
- Sivathanu, S.; Sampath, S.; Sridhar, I. Case 1: Recurrent Omphalitis and Nonhealing Ulcers in a 7-month-old Girl. Pediatr. Rev. 2016, 37, 491–493. [Google Scholar] [CrossRef] [PubMed]
- Webber, E.C.; Church, J.; Rand, T.H.; Shah, A.J. Leukocyte adhesion deficiency in a female patient without delayed umbilical cord separation. J. Paediatr. Child Health 2007, 43, 406–408. [Google Scholar] [CrossRef]
- Mishra, A.; Gupta, M.; Dalvi, A.; Ghosh, K.; Madkaikar, M. Rapid Flow cytometric prenatal diagnosis of primary immunodeficiency (PID) disorders. J. Clin. Immunol. 2014, 34, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.M.; Brenchley, L.; Moutsopoulos, N.M. Primary immunodeficiencies reveal the essential role of tissue neutrophils in periodontitis. Immunol. Rev. 2019, 287, 226–235. [Google Scholar] [CrossRef]
- Kumar, A.; Gupta, A.; Rawat, A.; Ahuja, C.; Suri, D.; Singh, S. Brain Abscess in a Child with Leukocyte Adhesion Defect: An Unusual Association. J. Clin. Immunol. 2016, 36, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Hynes, R.O. Targeted mutations in cell adhesion genes: What have we learned from them? Dev. Biol. 1996, 180, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Etzioni, A.; Doerschuk, C.M.; Harlan, J.M. Of man and mouse: Leukocyte and endothelial adhesion molecule deficiencies. Blood 1999, 94, 3281–3288. [Google Scholar] [CrossRef]
- Shaw, J.M.; Al-Shamkhani, A.; Boxer, L.A.; Buckley, C.D.; Dodds, A.W.; Klein, N.; Nolan, S.M.; Roberts, I.; Roos, D.; Scarth, S.L.; et al. Characterization of four CD18 mutants in leucocyte adhesion deficient (LAD) patients with differential capacities to support expression and function of the CD11/CD18 integrins LFA-1, Mac-1 and p150,95. Clin. Exp. Immunol. 2001, 126, 311–318. [Google Scholar] [CrossRef]
- von Andrian, U.H.; Berger, E.M.; Ramezani, L.; Chambers, J.D.; Ochs, H.D.; Harlan, J.M.; Paulson, J.C.; Etzioni, A.; Arfors, K.E. In vivo behavior of neutrophils from two patients with distinct inherited leukocyte adhesion deficiency syndromes. J. Clin. Investig. 1993, 91, 2893–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etzioni, A.; Frydman, M.; Pollack, S.; Avidor, I.; Phillips, M.L.; Paulson, J.C.; Gershoni-Baruch, R. Brief report: Recurrent severe infections caused by a novel leukocyte adhesion deficiency. N. Engl. J. Med. 1992, 327, 1789–1792. [Google Scholar] [CrossRef] [PubMed]
- Crazzolara, R.; Maurer, K.; Schulze, H.; Zieger, B.; Zustin, J.; Schulz, A.S. A new mutation in the KINDLIN-3 gene ablates integrin-dependent leukocyte, platelet, and osteoclast function in a patient with leukocyte adhesion deficiency-III. Pediatr. Blood Cancer 2015, 62, 1677–1679. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.S.; Shigeoka, A.O.; Li, W.; Adams, R.H.; Prescott, S.M.; McIntyre, T.M.; Zimmerman, G.A.; Lorant, D.E. A novel syndrome of variant leukocyte adhesion deficiency involving defects in adhesion mediated by beta1 and beta2 integrins. Blood 2001, 97, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Fagerholm, E.D.; Moran, R.J.; Violante, I.R.; Leech, R.; Friston, K.J. Dynamic causal modelling of phase-amplitude interactions. NeuroImage 2020, 208, 116452. [Google Scholar] [CrossRef] [PubMed]
- Alon, R.; Aker, M.; Feigelson, S.; Sokolovsky-Eisenberg, M.; Staunton, D.E.; Cinamon, G.; Grabovsky, V.; Shamri, R.; Etzioni, A. A novel genetic leukocyte adhesion deficiency in subsecond triggering of integrin avidity by endothelial chemokines results in impaired leukocyte arrest on vascular endothelium under shear flow. Blood 2003, 101, 4437–4445. [Google Scholar] [CrossRef] [Green Version]
- Etzioni, A. Genetic etiologies of leukocyte adhesion defects. Curr. Opin. Immunol. 2009, 21, 481–486. [Google Scholar] [CrossRef]
- Fan, Z.; Ley, K. Leukocyte Adhesion Deficiency IV. Monocyte Integrin Activation Deficiency in Cystic Fibrosis. Am. J. Respir. Crit. Care Med. 2016, 193, 1075–1077. [Google Scholar] [CrossRef] [Green Version]
- Sorio, C.; Montresor, A.; Bolomini-Vittori, M.; Caldrer, S.; Rossi, B.; Dusi, S.; Angiari, S.; Johansson, J.E.; Vezzalini, M.; Leal, T.; et al. Mutations of Cystic Fibrosis Transmembrane Conductance Regulator Gene Cause a Monocyte-Selective Adhesion Deficiency. Am. J. Respir. Crit. Care Med. 2016, 193, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, X.; Wang, P.; Wang, H.; Jiang, W.; Sun, J.; Wang, X.; Zhai, X. Successful umbilical cord blood transplantation in children with leukocyte adhesion deficiency type I. Transl. Pediatr. 2020, 9, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Le Deist, F.; Cavazzana-Calvo, M.; Benkerrou, M.; Haddad, E.; Blanche, S.; Hartmann, W.; Friedrich, W.; Fischer, A. Results of allogeneic bone marrow transplantation in patients with leukocyte adhesion deficiency. Blood 1995, 86, 1629–1635. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki-Nakashimada, M.; Maravillas-Montero, J.L.; Berron-Ruiz, L.; Lopez-Ortega, O.; Ramirez-Alejo, N.; Acevedo-Ochoa, E.; Rivas-Larrauri, F.; Llamas-Guillen, B.; Blancas-Galicia, L.; Scheffler-Mendoza, S.; et al. Successful adjunctive immunoglobulin treatment in patients affected by leukocyte adhesion deficiency type 1 (LAD-1). Immunol. Res. 2015, 61, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Moutsopoulos, N.M.; Zerbe, C.S.; Wild, T.; Dutzan, N.; Brenchley, L.; DiPasquale, G.; Uzel, G.; Axelrod, K.C.; Lisco, A.; Notarangelo, L.D.; et al. Interleukin-12 and Interleukin-23 Blockade in Leukocyte Adhesion Deficiency Type 1. N. Engl. J. Med. 2017, 376, 1141–1146. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Lim, J.H.; Kajikawa, T.; Li, X.; Vallance, B.A.; Moutsopoulos, N.M.; Chavakis, T.; Hajishengallis, G. Macrophage beta2-Integrins Regulate IL-22 by ILC3s and Protect from Lethal Citrobacter rodentium-Induced Colitis. Cell Rep. 2019, 26, 1614–1626.e5. [Google Scholar] [CrossRef] [Green Version]
- Marsili, M.; Lougaris, V.; Lucantoni, M.; Di Marzio, D.; Baronio, M.; Vitali, M.; Lombardi, G.; Chiarelli, F.; Breda, L. Successful anti-TNF-alpha treatment in a girl with LAD-1 disease and autoimmune manifestations. J. Clin. Immunol. 2014, 34, 788–791. [Google Scholar] [CrossRef]
- Nakagawa, M.; Bondy, G.P.; Waisman, D.; Minshall, D.; Hogg, J.C.; van Eeden, S.F. The effect of glucocorticoids on the expression of L-selectin on polymorphonuclear leukocyte. Blood 1999, 93, 2730–2737. [Google Scholar] [CrossRef]
- Ballabh, P.; Kumari, J.; Krauss, A.N.; Shin, J.J.; Jain, A.; Auld, P.A.; Lesser, M.L.; Cunningham-Rundles, S. Soluble E-selectin, soluble L-selectin and soluble ICAM-1 in bronchopulmonary dysplasia, and changes with dexamethasone. Pediatrics 2003, 111, 461–468. [Google Scholar] [CrossRef]
- Koehne, P.S.; Wagner, M.H.; Willam, C.; Sonntag, J.; Buhrer, C.; Obladen, M. Soluble intercellular cell adhesion molecule-1 and L-selectin plasma concentrations and response to surfactant in preterm infants. Pediatr. Crit. Care Med. 2002, 3, 23–28. [Google Scholar] [CrossRef]
- Lorant, D.E.; Li, W.; Tabatabaei, N.; Garver, M.K.; Albertine, K.H. P-selectin expression by endothelial cells is decreased in neonatal rats and human premature infants. Blood 1999, 94, 600–609. [Google Scholar] [CrossRef]
- Pieh, C.; Kruger, M.; Lagreze, W.A.; Gimpel, C.; Buschbeck, C.; Zirrgiebel, U.; Agostini, H.T. Plasma sE-selectin in premature infants: A possible surrogate marker of retinopathy of prematurity. Investig. Ophthalmol. Vis. Sci. 2010, 51, 3709–3713. [Google Scholar] [CrossRef]
- Zielinska, K.A.; Van Moortel, L.; Opdenakker, G.; De Bosscher, K.; Van den Steen, P.E. Endothelial Response to Glucocorticoids in Inflammatory Diseases. Front. Immunol. 2016, 7, 592. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, R.; Stack, T.; Yi, S.; Mathew, B.; Shull, K.R.; Scott, E.A.; Mathew, M.T.; Bijukumar, D.R. High Density Display of an Anti-Angiogenic Peptide on Micelle Surfaces Enhances Their Inhibition of alphavbeta3 Integrin-Mediated Neovascularization In Vitro. Nanomaterials 2020, 10, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, K.; Rivera-Nieves, J.; Sandborn, W.J.; Shattil, S. Integrin-based therapeutics: Biological basis, clinical use and new drugs. Nat. Rev. Drug Discov. 2016, 15, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frenette, P.S. Locking a leukocyte integrin with statins. N. Engl. J. Med. 2001, 345, 1419–1421. [Google Scholar] [CrossRef]
- Turunen, R.; Nupponen, I.; Siitonen, S.; Repo, H.; Andersson, S. Onset of mechanical ventilation is associated with rapid activation of circulating phagocytes in preterm infants. Pediatrics 2006, 117, 448–454. [Google Scholar] [CrossRef]
- Desai, L.P.; Sinclair, S.E.; Chapman, K.E.; Hassid, A.; Waters, C.M. High tidal volume mechanical ventilation with hyperoxia alters alveolar type II cell adhesion. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 293, L769–L778. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vrachnis, N.; Zygouris, D.; Vrachnis, D.; Roussos, N.; Loukas, N.; Antonakopoulos, N.; Paltoglou, G.; Barbounaki, S.; Valsamakis, G.; Iliodromiti, Z. Perinatal Inflammation: Could Partial Blocking of Cell Adhesion Molecule Function Be a Solution? Children 2021, 8, 380. https://doi.org/10.3390/children8050380
Vrachnis N, Zygouris D, Vrachnis D, Roussos N, Loukas N, Antonakopoulos N, Paltoglou G, Barbounaki S, Valsamakis G, Iliodromiti Z. Perinatal Inflammation: Could Partial Blocking of Cell Adhesion Molecule Function Be a Solution? Children. 2021; 8(5):380. https://doi.org/10.3390/children8050380
Chicago/Turabian StyleVrachnis, Nikolaos, Dimitrios Zygouris, Dionysios Vrachnis, Nikolaos Roussos, Nikolaos Loukas, Nikolaos Antonakopoulos, Georgios Paltoglou, Stavroula Barbounaki, Georgios Valsamakis, and Zoi Iliodromiti. 2021. "Perinatal Inflammation: Could Partial Blocking of Cell Adhesion Molecule Function Be a Solution?" Children 8, no. 5: 380. https://doi.org/10.3390/children8050380
APA StyleVrachnis, N., Zygouris, D., Vrachnis, D., Roussos, N., Loukas, N., Antonakopoulos, N., Paltoglou, G., Barbounaki, S., Valsamakis, G., & Iliodromiti, Z. (2021). Perinatal Inflammation: Could Partial Blocking of Cell Adhesion Molecule Function Be a Solution? Children, 8(5), 380. https://doi.org/10.3390/children8050380