Multiple-stage Precursor Ion Separation and High Resolution Mass Spectrometry toward Structural Characterization of 2,3-Diacyltrehalose Family from Mycobacterium tuberculosis

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Sample Preparation
2.3. Mass Spectrometry
2.4. Nomenclature
3. Results and Discussion
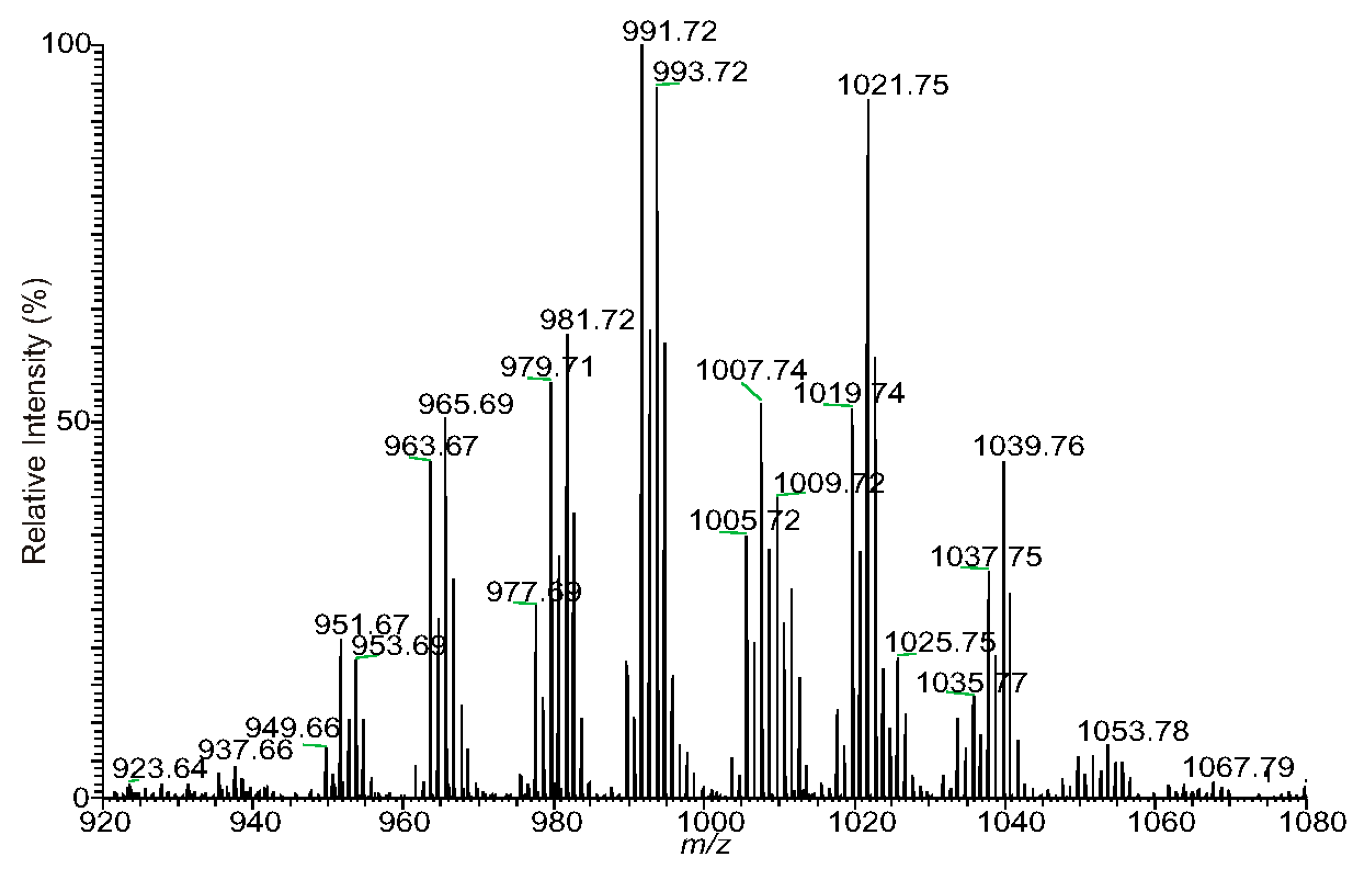
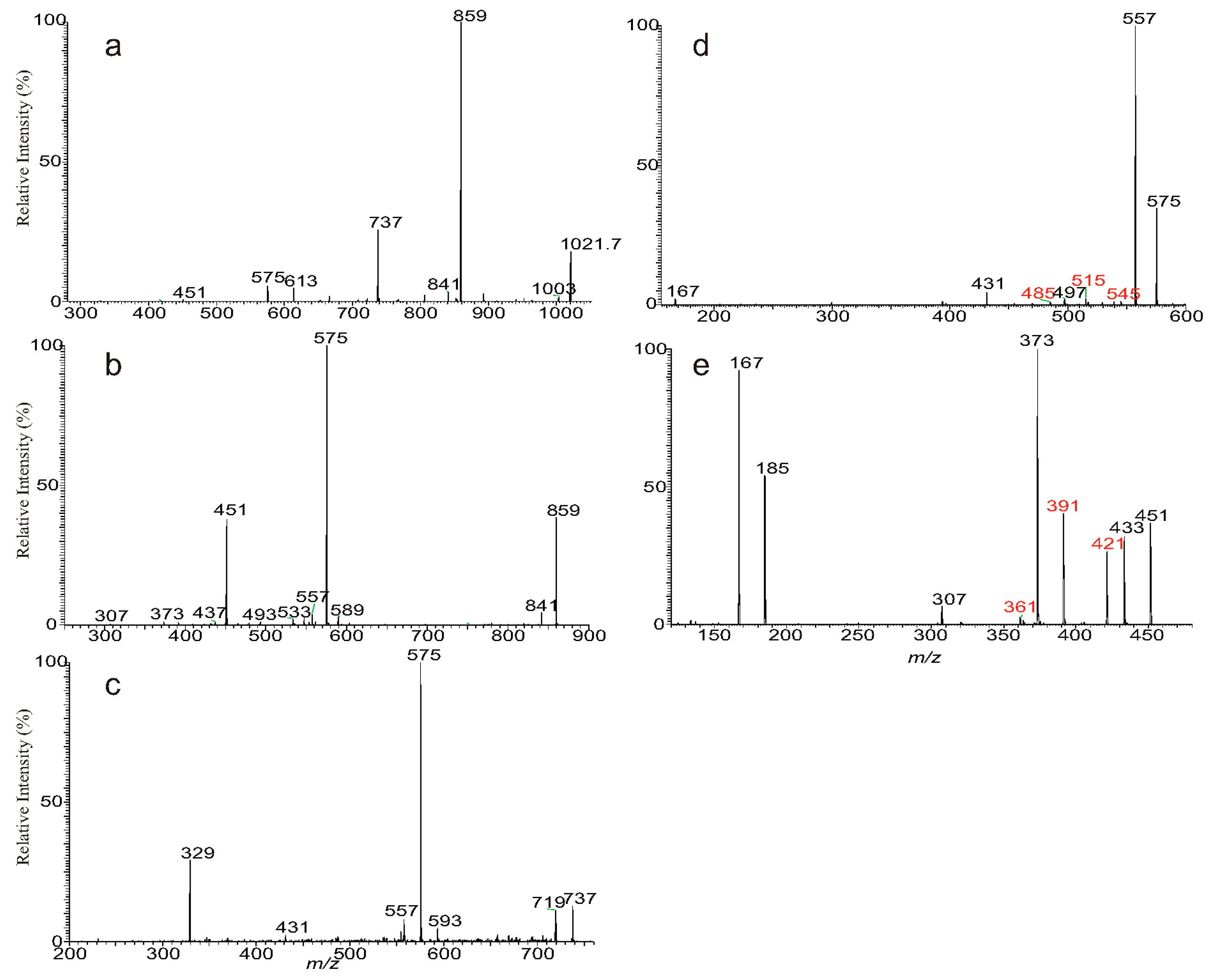
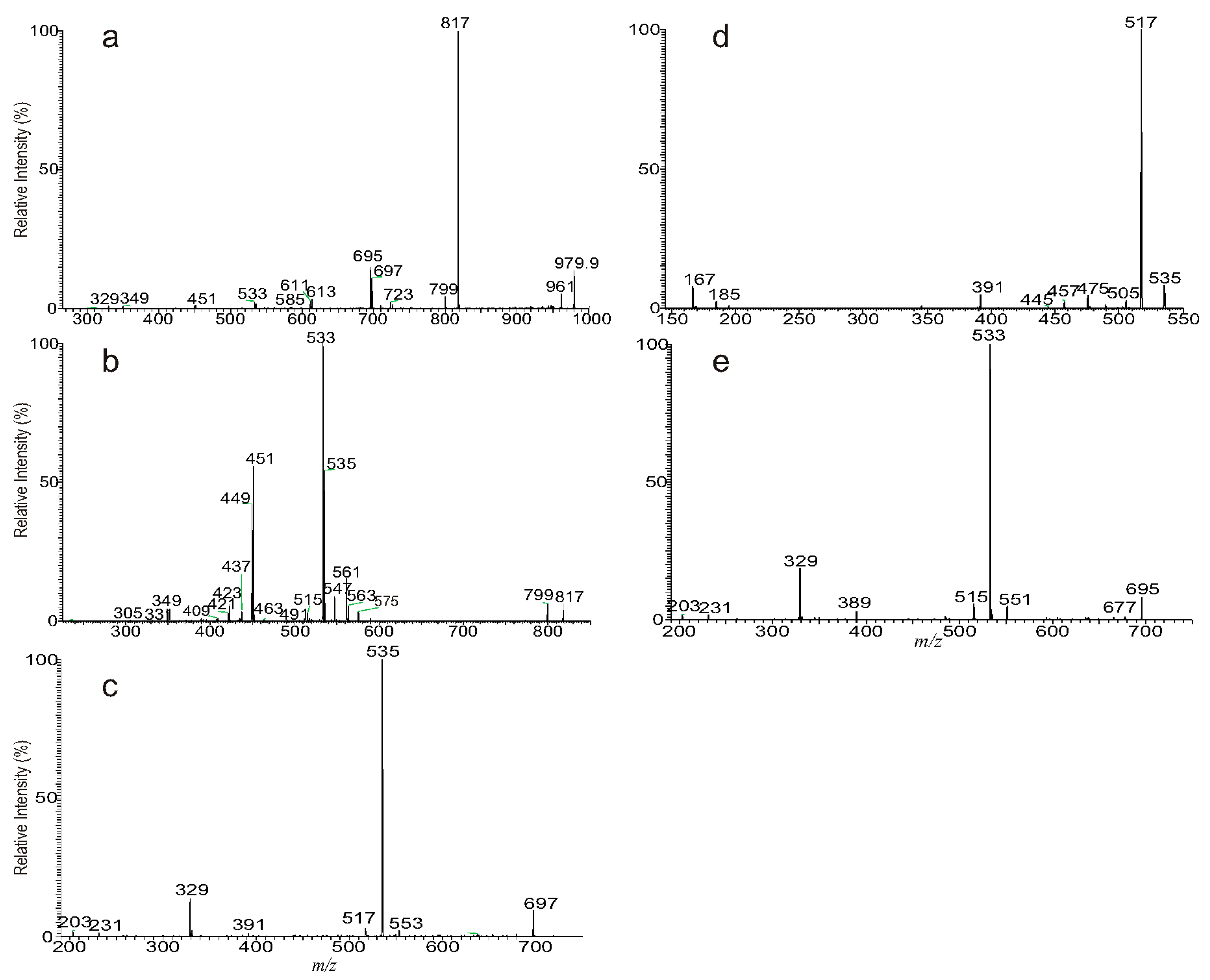
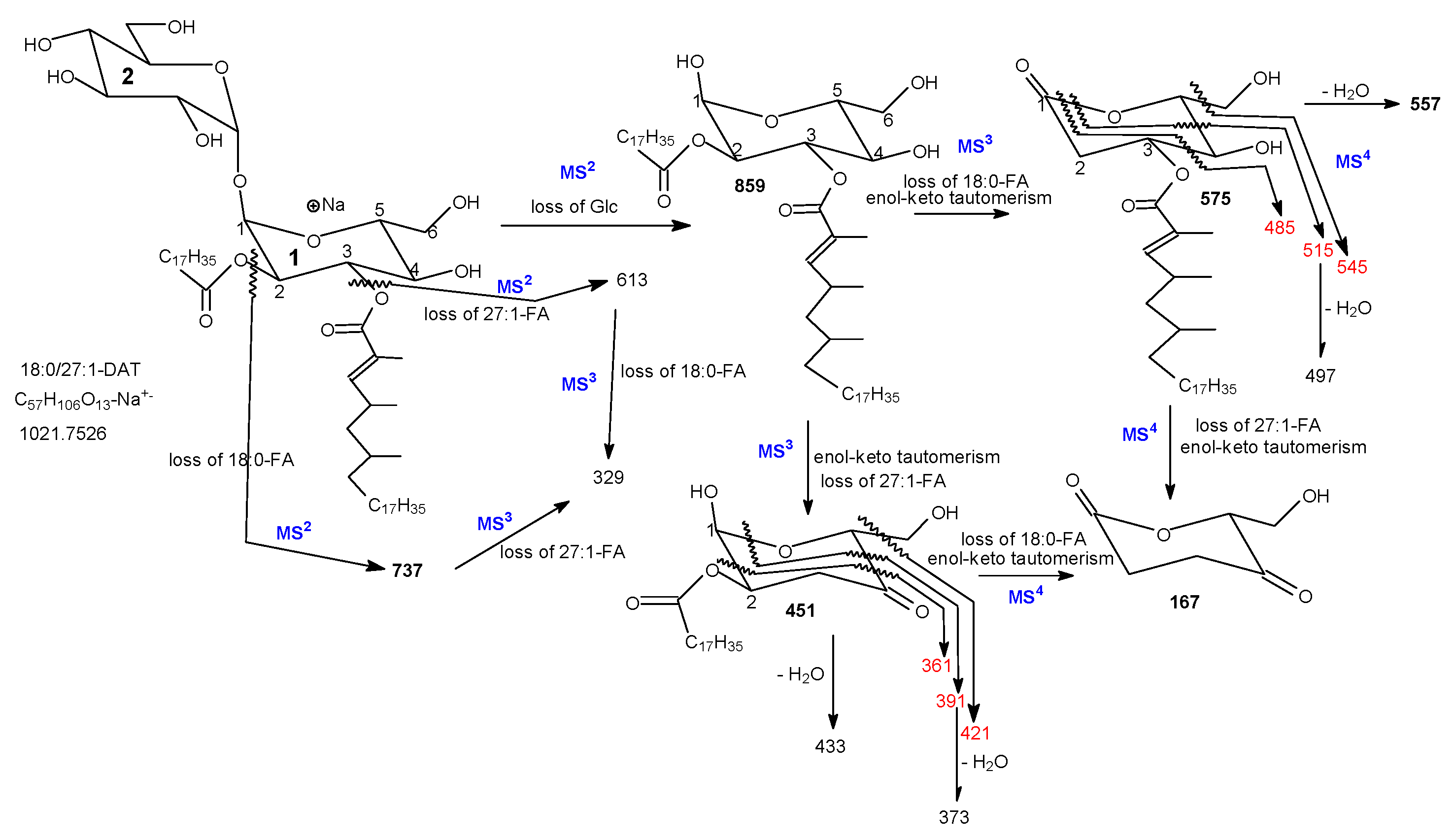
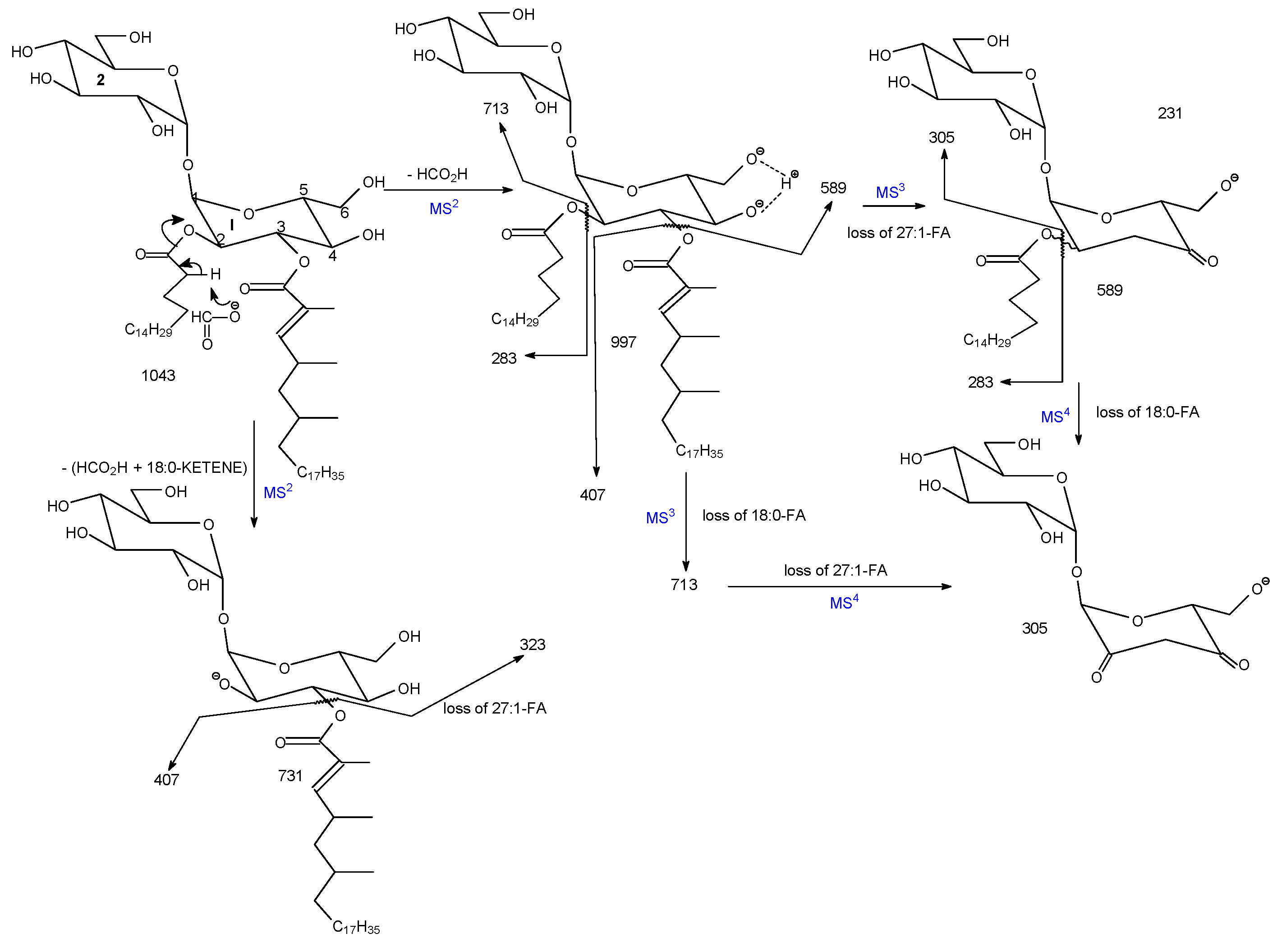
3.1. The Fragmentation Processes of the [M + Na]+ Ions of DAT Revealed by LIT MSn
3.2. LIT MSn on the [M + Na]+ Ions of DAT for Stereoisomer Recognition
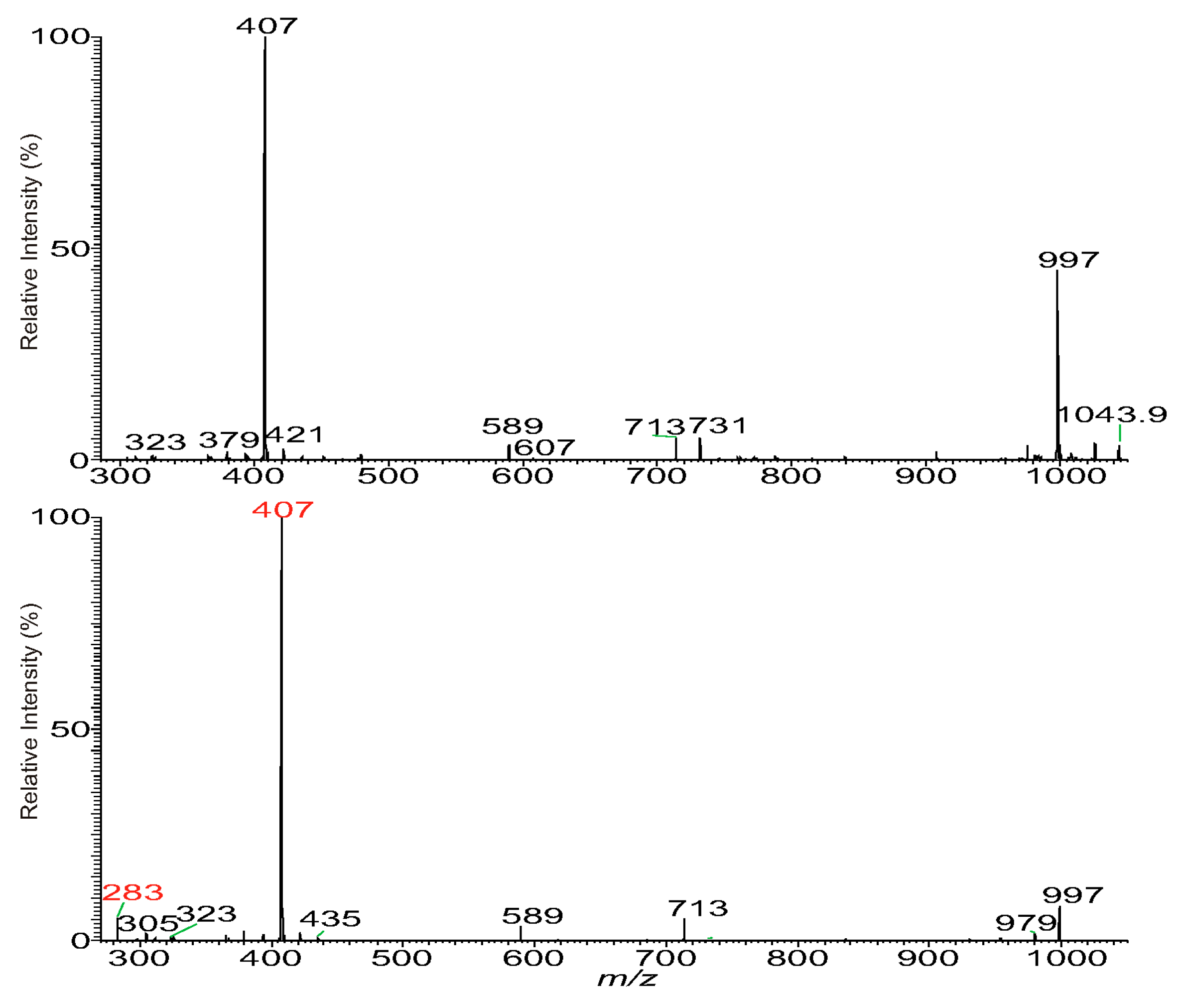
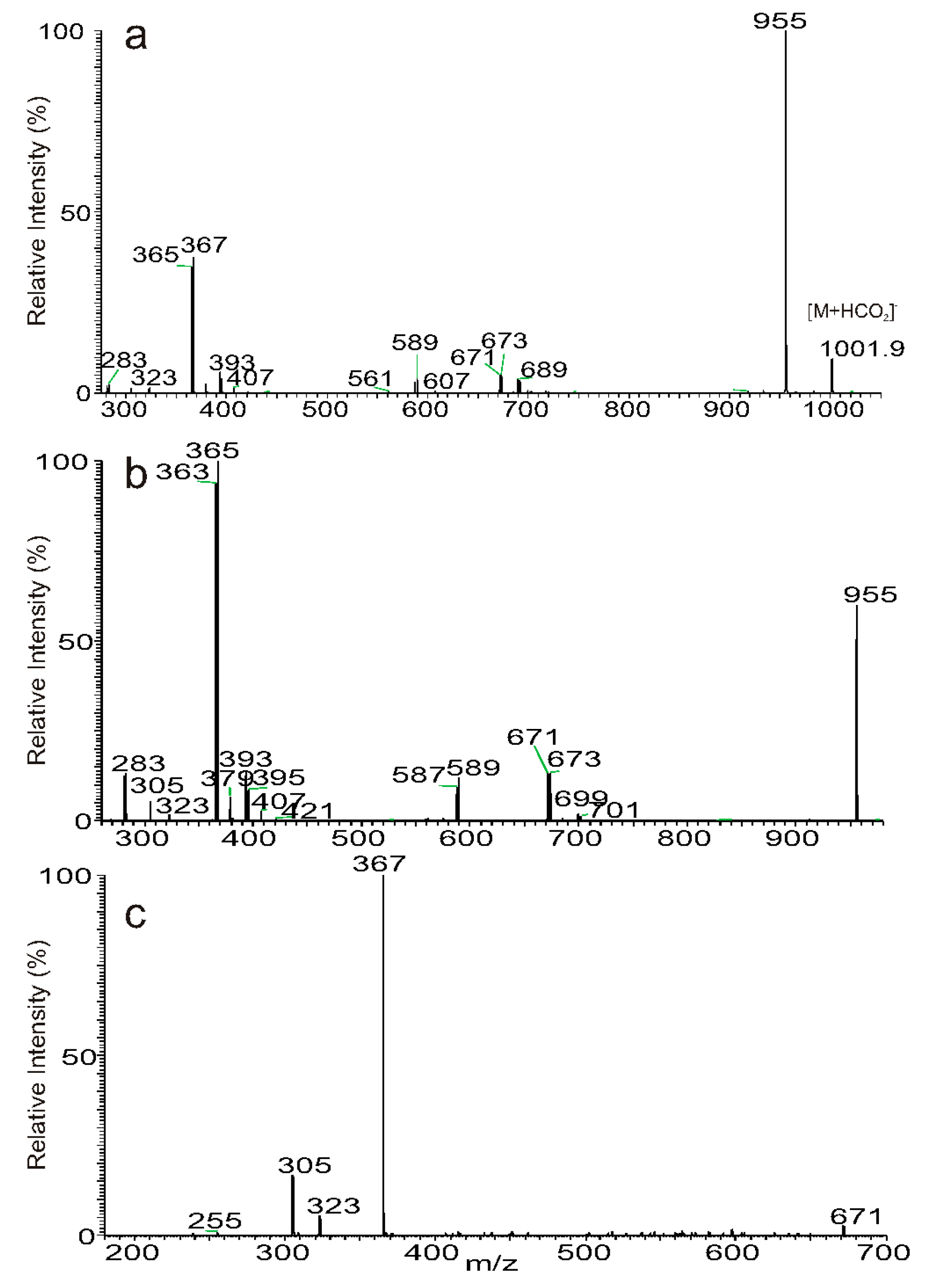
3.3. The Fragmentation Processes of the [M + HCO2]− Ions of DAT Revealed by LIT MSn
3.4. Recognition of Stereoisomers Applying LIT MSn on the [M + HCO2]− Ions
3.5. Characterization of Minor Species Applying LIT MSn on the [M + HCO2]− Ions
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Johnson, J.V.; Yost, R.A.; Kelley, P.E.; Bradford, D.C. Tandem-in-space and tandem-in-time mass spectrometry: triple quadrupoles and quadrupole ion traps. Anal. Chem. 1990, 62, 2162–2172. [Google Scholar] [CrossRef]
- Markarov, A.; Cousijn, E.; Cantebury, J.; Denisov, E.; Thoeing, C.; Lange, O.; Kreutzman, A.; Ayzikov, K.; Damoc, E.; Tabiwang, A.; et al. Extension of Orbitrap capabilities to enable new applications. In Proceedings of the 65th Conference on Mass Spectrometry and Allied Topics, Indianapolis, IN, USA, 4 June 2017. [Google Scholar]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perry, R.H.; Cooks, R.G.; Noll, R.J. Orbitrap mass spectrometry: Instrumentation, ion motion and applications. Mass Spectrom. Rev. 2008, 27, 661–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senyuva, H.Z.; Gökmen, V.; Sarikaya, E.A. Future perspectives in Orbitrap™-high-resolution mass spectrometry in food analysis: A review. Food Addit. Contam. A 2015, 32, 1568–1606. [Google Scholar] [CrossRef] [PubMed]
- Jabbour, R.E.; Snyder, A.P. Chap. 14—Mass spectrometry-based proteomics techniques for biological identification. In Biological Identification; Schaudies, R.P., Ed.; Woodhead Publishing: Sawston, UK, 2014; pp. 370–430. [Google Scholar]
- Eliuk, S.; Makarov, A. Evolution of Orbitrap Mass Spectrometry Instrumentation. Ann. Rev. Anal. Chem. 2015, 8, 61–80. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, E.R.; Streeter, C.; Turk, J.; Hsu, F.-F. Characterization of Sulfolipids of Mycobacterium tuberculosis H37Rv by Multiple-Stage Linear Ion-Trap High-Resolution Mass Spectrometry with Electrospray Ionization Reveals That the Family of Sulfolipid II Predominates. Biochemistry 2011, 50, 9135–9147. [Google Scholar] [CrossRef] [PubMed]
- Hsu, F.-F. Characterization of Hydroxyphthioceranoic and Phthioceranoic Acids by Charge-Switch Derivatization and CID Tandem Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2016, 27, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Hsu, F.F.; Turk, J.; Owens, R.M.; Rhoades, E.R.; Russell, D.G. Structural Characterization of Phosphatidyl-myo-Inositol Mannosides from Mycobacterium bovis Bacillus Calmette Guerin by Multiple-Stage Quadrupole Ion-Trap Mass Spectrometry with Electrospray Ionization. II. Monoacyl- and Diacyl-PIMs. J. Am. Soc. Mass Spectrom. 2007, 18, 479–492. [Google Scholar] [CrossRef]
- Hsu, F.F.; Turk, J.; Owens, R.M.; Rhoades, E.R.; Russell, D.G. Structural characterization of phosphatidyl-myo-inositol mannosides from Mycobacterium bovis Bacillus Calmette Guerin by multiple-stage quadrupole ion-trap mass spectrometry with electrospray ionization. I. PIMs and lyso-PIMs. J. Am. Soc. Mass Spectrom. 2007, 18, 466–478. [Google Scholar] [CrossRef]
- Flentie, K.N.; Stallings, C.L.; Turk, J.; Minnaard, A.J.; Hsu, F.-F. Characterization of phthiocerol and phthiodiolone dimycocerosate esters of M. tuberculosis by multiple-stage linear ion-trap MS. J. Lipid Res. 2016, 57, 142–155. [Google Scholar] [CrossRef]
- Hoppe, H.C.; de Wet, B.J.; Cywes, C.; Daffe, M.; Ehlers, M.R. Identification of phosphatidylinositol mannoside as a mycobacterial adhesin mediating both direct and opsonic binding to nonphagocytic mammalian cells. Infect. Immun. 1997, 65, 3896–3905. [Google Scholar] [PubMed]
- Howard, N.C.; Marin, N.D.; Ahmed, M.; Rosa, B.A.; Martin, J.; Bambouskova, M.; Sergushichev, A.; Loginicheva, E.; Kurepina, N.; Rangel-Moreno, J.; et al. Mycobacterium tuberculosis carrying a rifampicin drug resistance mutation reprograms macrophage metabolism through cell wall lipid changes. Nat. Microbiol. 2018, 3, 1099–1108. [Google Scholar] [CrossRef] [PubMed]
- DaffÉ, M.; Lacave, C.; LanÉElle, M.-A.; Gillois, M.; LanÉElle, G. Polyphthienoyl trehalose, glycolipids specific for virulent strains of the tubercle bacillus. Eur. J. Biochem. 1988, 172, 579–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minnikin, D.E.; Dobson, G.; Sesardic, D.; Ridell, M. Mycolipenates and Mycolipanolates of Trehalose from Mycobacterium tuberculosis. J. Gen. Microbiol. 1985, 131, 1369–1374. [Google Scholar] [CrossRef] [PubMed]
- Munoz, M.; Laneelle, M.A.; Luquin, M.; Torrelles, J.; Julian, E.; Ausina, V.; Daffe, M. Occurrence of an antigenic triacyl trehalose in clinical isolates and reference strains of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 1997, 157, 251–259. [Google Scholar] [CrossRef]
- Lemassu, A.; Laneelle, M.A.; Daffe, M. Revised structure of a trehalose-containing immunoreactive glycolipid of Mycobacterium tuberculosis. FEMS Microbiol. Lett. 1991, 62, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Ariza, M.A.; Martín-Luengo, F.; Valero-Guillén, P.L. A family of diacyltrehaloses isolated from Mycobacterium fortuitum. Microbiology 1994, 140, 1989–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, M.A.; Valero-Guillen, P.L. Delineation of molecular species of a family of diacyltrehaloses from Mycobacterium fortuitum by mass spectrometry. FEMS Microbiol. Lett. 1994, 119, 279–282. [Google Scholar] [CrossRef]
- Besra, G.S.; Bolton, R.C.; McNeil, M.R.; Ridell, M.; Simpson, K.E.; Glushka, J.; Van Halbeek, H.; Brennan, P.J.; Minnikin, D.E. Structural elucidation of a novel family of acyltrehaloses from Mycobacterium tuberculosis. Biochemistry 1992, 31, 9832–9837. [Google Scholar] [CrossRef]
- Gautier, N.; Marín, L.M.L.; Lanéelle, M.A.; Daffé, M. Structure of mycoside F, a family of trehalose-containing glycolipids of Mycobacterium fortuitum. FEMS Microbiol. Lett. 1992, 77, 81–87. [Google Scholar]
- Lee, K.-S.; Dubey, V.S.; Kolattukudy, P.E.; Song, C.-H.; Shin, A.R.; Jung, S.-B.; Yang, C.-S.; Kim, S.-Y.; Jo, E.-K.; Park, J.-K.; et al. Diacyltrehalose of Mycobacterium tuberculosis inhibits lipopolysaccharide- and mycobacteria-induced proinflammatory cytokine production in human monocytic cells. FEMS Microbiol. Lett. 2007, 267, 121–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saavedra, R.; Segura, E.; Leyva, R.; Esparza, L.A.; López-Marín, L.M. Mycobacterial Di-O-Acyl-Trehalose Inhibits Mitogen- and Antigen-Induced Proliferation of Murine T Cells In Vitro. Clin. Diagn. Lab. Immun. 2001, 8, 1081–1088. [Google Scholar] [CrossRef] [Green Version]
- Bailo, R.; Bhatt, A.; Ainsa, J.A. Lipid transport in Mycobacterium tuberculosis and its implications in virulence and drug development. Biochem. Pharmacol. 2015, 96, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Papa, F.; Cruaud, P.; David, H.L. Antigenicity and specificity of selected glycolipid fractions from Mycobacterium tuberculosis. Res. Microbiol. 1989, 140, 569–578. [Google Scholar] [CrossRef]
- Hamid, M.E.; Fraser, J.L.; Wallace, P.A.; Besra, G.; Goodfellow, M.; Minnikin, D.E. Antigenic glycolipids of Mycobacterium fortuitum based on trehalose acylated with 2-methyloctadec-2-enoic acid. Lett. Appl. Microbiol. Rev. 1993, 16, 132–135. [Google Scholar] [CrossRef]
- Ridell, M.; Wallerstr6m, G.; Minnikin, D.E.; Bolton, R.C.; Magnusson, M. A comparative serological study of antigenic glycolipids from Mycobacteriurn tuberculosis. Tubercle Lung Dis. 1992, 73, 71–75. [Google Scholar] [CrossRef]
- Tórtola, M.T.; Lanéelle, M.A.; Martín-Casabona, N. Comparison of two 2,3-diacyl trehalose antigens from Mycobacterium tuberculosis and Mycobacterium fortuitum for serology in tuberculosis patients. Clin. Diagn. Lab. Immun. 1996, 3, 563–566. [Google Scholar]
- Prabhakar, S.; Vivès, T.; Ferrières, V.; Benvegnu, T.; Legentil, L.; Lemiègre, L. A fully enzymatic esterification/transesterification sequence for the preparation of symmetrical and unsymmetrical trehalose diacyl conjugates. Green Chem. 2017, 19, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Botté, C.Y.; Deligny, M.; Roccia, A.; Bonneau, A.-L.; Saïdani, N.; Hardré, H.; Aci, S.; Yamaryo-Botté, Y.; Jouhet, J.; Dubots, E.; et al. Chemical inhibitors of monogalactosyldiacylglycerol synthases in Arabidopsis thaliana. Nat. Chem. Biol. 2011, 7, 834–842. [Google Scholar] [CrossRef]
- Olsen, J.V.; Macek, B.; Lange, O.; Makarov, A.; Horning, S.; Mann, M. Higher-energy C-trap dissociation for peptide modification analysis. Nat. Meth. 2007, 4, 709–712. [Google Scholar] [CrossRef]
- Hsu, F.F. Mass spectrometry-based shotgun lipidomics—A critical review from the technical point of view. Anal. Bioanal. Chem. 2018, 410, 6387–6409. [Google Scholar] [CrossRef] [PubMed]
- Frankfater, C.; Jiang, X.; Hsu, F.F. Characterization of Long-Chain Fatty Acid as N-(4-Aminomethylphenyl) Pyridinium Derivative by MALDI LIFT-TOF/TOF Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2018, 29, 1688–1699. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measured m/z | Theo. Mass m/z | Deviation mmu | Rel. Intensity % | Ele. Composition | Major structures | Minor structures |
|---|---|---|---|---|---|---|
| 935.6436 | 935.6430 | 0.55 | 3.42 | C51H92O13Na | 14:1/25:1 | 12:1/27:1 |
| 937.6594 | 937.6587 | 0.73 | 4.34 | C51H94O13Na | 14:0/25:1 | 12:0/27:1; 16:1/23:0; 15:1/24:0; 13:0/26:1 |
| 939.6747 | 939.6743 | 0.37 | 1.48 | C51H96O13Na | * | |
| 949.6585 | 949.6587 | −0.19 | 6.58 | C52H94O13Na | 16:1/24:1 | |
| 951.6742 | 951.6743 | −0.15 | 20.8 | C52H96O13Na | 16:1/24:0 | 16:0/24:1; 15:0/25:1; 17:1/23:0; 14:0/26:1; 13:0/27:1 |
| 953.6898 | 953.6900 | −0.12 | 18.49 | C52H98O13Na | 16:0/24:0 | |
| 961.6587 | 961.6587 | 0.01 | 4.31 | C53H94O13Na | * | |
| 963.6743 | 963.6743 | −0.05 | 44.16 | C53H96O13Na | 16:1/25:1; 14:1/27:1 | |
| 965.6898 | 965.6900 | −0.12 | 50.07 | C53H98O13Na | 16:0/25:1 | 14:0/27:1; 18:0/23:1 |
| 967.7054 | 967.7056 | −0.22 | 12.26 | C53H100O13Na | 18:0/23:0; 17:0/24:0 | 16:0/25:0 |
| 975.6740 | 975.6743 | −0.3 | 3.03 | C54H96O13Na | * | |
| 977.6899 | 977.6900 | −0.06 | 25.16 | C54H98O13Na | 16:1/26:1 | |
| 979.7056 | 979.7056 | −0.06 | 54.48 | C54H100O13Na | 18:0/24:1; 18:1/24:0 | 16:0/26:1; 17:0/25:1; 16:1/26:0; 15:0/27:1 |
| 981.7211 | 981.7213 | −0.15 | 62.62 | C54H102O13Na | 18:0/24:0 | 17:0/25:0; 16:0/26:0 |
| 989.6898 | 989.6900 | −0.17 | 18.03 | C55H98O13Na | 16:1/27:2 | |
| 991.7054 | 991.7056 | −0.2 | 100 | C55H100O13Na | 16:1/27:1 | 18:1/25:1; 16:0/27:2; 17:0/26:2 |
| 993.7210 | 993.7213 | −0.29 | 95.91 | C55H102O13Na | 16:0/27:1 | 18:0/25:1; 17:0/26:1 |
| 1005.7211 | 1005.7213 | −0.2 | 34.31 | C56H102O13Na | 16:1/28:1; 17:1/27:1 | 17:0/27:2; 18:1/26:1; 18:0/26:2 |
| 1007.7366 | 1007.7369 | −0.31 | 51.54 | C56H104O13Na | 17:0/27:1 | 16:0/28:1; 18:0/26:1; 19:0/25:1; 16:1/28:0 |
| 1009.7518 | 1009.7526 | −0.75 | 7.65 | C56H106O13Na | 18:0/26:0 | 20:0/24:0 |
| 1017.7209 | 1017.7213 | −0.38 | 11.79 | C57H102O13Na | 18:1/27:2 | |
| 1019.7367 | 1019.7369 | −0.25 | 52.39 | C57H104O13Na | 18:1/27:1 | 18:0/27:2; 17:1/28:1 |
| 1021.7522 | 1021.7526 | −0.37 | 92.1 | C57H106O13Na | 18:0/27:1 | 17:0/28:1 |
| 1033.7521 | 1033.7526 | −0.47 | 10.49 | C58H106O13Na | 18:1/28:1 | 19:0/27:2; 19:1/27:1; 18:0/28:2; 19:0/27:2 |
| 1035.7678 | 1035.7682 | −0.45 | 13.33 | C58H108O13Na | 19:0/27:1; 18:0/28:1 | 18:1/28:0; 17:0/29:1 |
| 1047.7677 | 1047.7682 | −0.5 | 2.56 | C59H108O13Na | 18:0/28:2 | |
| 1049.7835 | 1049.7839 | −0.31 | 5.47 | C59H110O13Na | 19:0/28:1 | 20:0/27:1; 18:0/29:1; 17:0/30:1 |
| 1061.7837 | 1061.7839 | −0.19 | 1.63 | C60H110O13Na | 18:1/30:1 | |
| 1063.7991 | 1063.7995 | −0.38 | 1.83 | C60H112O13Na | 18:0/30:1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frankfater, C.; Abramovitch, R.B.; Purdy, G.E.; Turk, J.; Legentil, L.; Lemiègre, L.; Hsu, F.-F. Multiple-stage Precursor Ion Separation and High Resolution Mass Spectrometry toward Structural Characterization of 2,3-Diacyltrehalose Family from Mycobacterium tuberculosis. Separations 2019, 6, 4. https://doi.org/10.3390/separations6010004
Frankfater C, Abramovitch RB, Purdy GE, Turk J, Legentil L, Lemiègre L, Hsu F-F. Multiple-stage Precursor Ion Separation and High Resolution Mass Spectrometry toward Structural Characterization of 2,3-Diacyltrehalose Family from Mycobacterium tuberculosis. Separations. 2019; 6(1):4. https://doi.org/10.3390/separations6010004
Chicago/Turabian StyleFrankfater, Cheryl, Robert B. Abramovitch, Georgiana E. Purdy, John Turk, Laurent Legentil, Loïc Lemiègre, and Fong-Fu Hsu. 2019. "Multiple-stage Precursor Ion Separation and High Resolution Mass Spectrometry toward Structural Characterization of 2,3-Diacyltrehalose Family from Mycobacterium tuberculosis" Separations 6, no. 1: 4. https://doi.org/10.3390/separations6010004
APA StyleFrankfater, C., Abramovitch, R. B., Purdy, G. E., Turk, J., Legentil, L., Lemiègre, L., & Hsu, F. -F. (2019). Multiple-stage Precursor Ion Separation and High Resolution Mass Spectrometry toward Structural Characterization of 2,3-Diacyltrehalose Family from Mycobacterium tuberculosis. Separations, 6(1), 4. https://doi.org/10.3390/separations6010004