Inadequate Sampling Rates Can Undermine the Reliability of Ecological Interaction Estimation



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
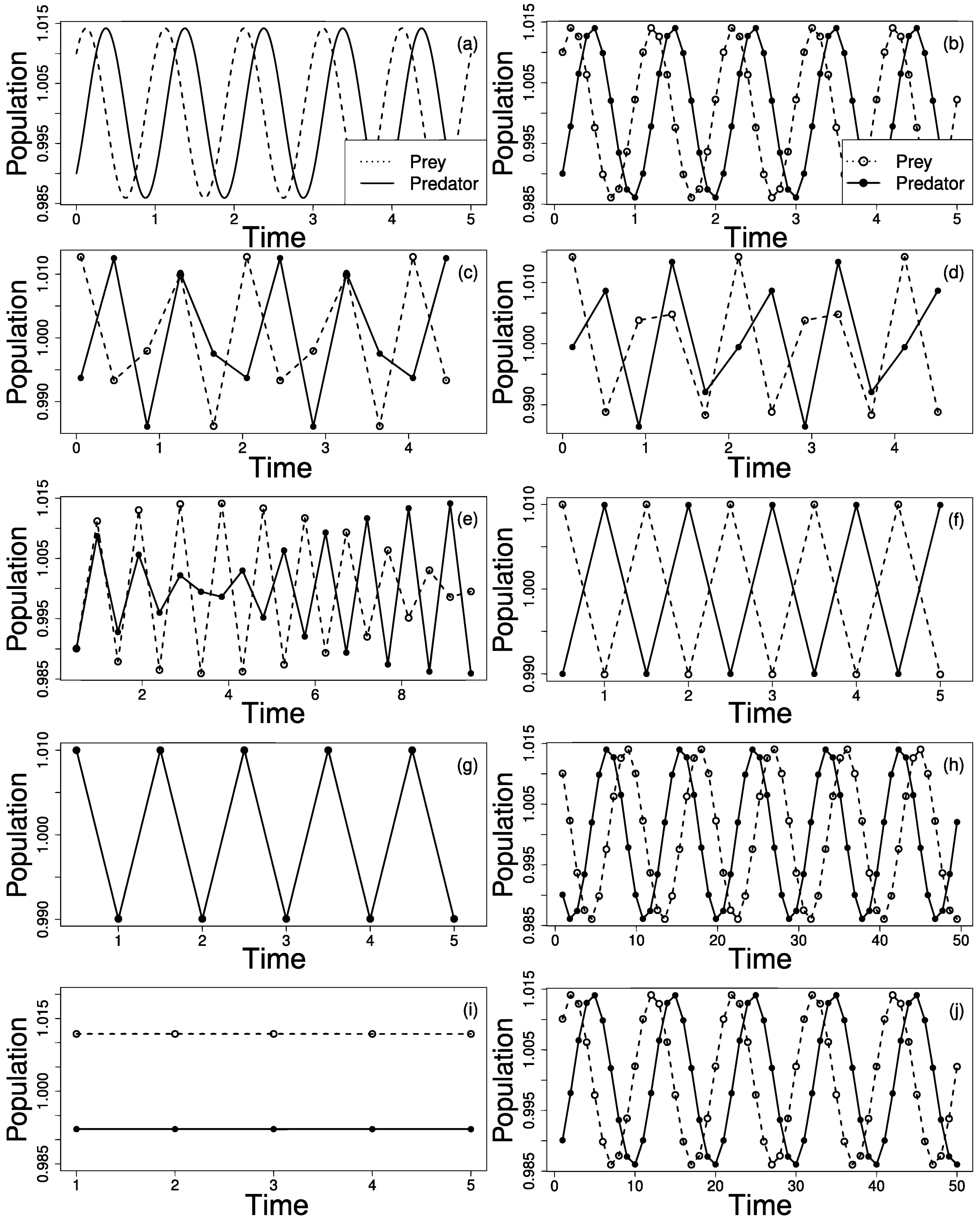
2. Aliasing the Lotka–Volterra Model
3. Sampling Effects
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LVM | Lotka Voltera model |
References
- Bulmer, M.G. A Statistical Analysis of the 10-Year Cycle in Canada. J. Anim. Ecol. 1974, 43, 701–718. [Google Scholar] [CrossRef]
- Barbosa, P.; Caldas, A.; Riechert, S.A. Species Abundance Distribution and Predator–Prey Interactions: Theoretical and Applied Consequences. In Ecology of Predator–Prey Interactions; Oxford University Press: Oxford, UK, 2005; pp. 344–368. [Google Scholar]
- Elton, C.S.; Nicholson, M. The ten year cycle in numbers of lynx in Canada. J. Anim. Ecol. 1942, 11, 215–244. [Google Scholar] [CrossRef]
- Odum, E.P. Fundamentals of Ecology; Saunders: Philadelphia, PA, USA, 1953. [Google Scholar]
- Rosenzweig, M.L.; MacArthur, R.H. Graphical representation and stability conditions of predator–prey interactions. Am. Nat. 1963, 97, 209–223. [Google Scholar] [CrossRef]
- Murray, J.D. Mathematical Biology: I. An Introduction; Springer: New York, NY, USA, 2012. [Google Scholar]
- Kery, M.; Dorazio, R.M.; Soldaat, L.; van Strien, A.; Zuiderwijk, A.; Royle, J.A. Trend estimation in populations with imperfect detection. J. Appl. Ecol. 2009, 46, 1163–1172. [Google Scholar] [CrossRef]
- Kishida, O.; Trussell, G.C.; Mougi, A.; Nishimura, K. Evolutionary ecology of inducible morphological plasticity in predator–prey interaction: Toward the practical links with population ecology. Popul. Ecol. 2010, 52, 37–46. [Google Scholar] [CrossRef]
- Oppenheim, A.V.; Schafer, R.W. Sampling of continuous-time signals. In Discrete-Time Signal Processing; Prentice Hall: Upper Saddle River, NJ, USA, 1999; pp. 140–150. [Google Scholar]
- Green, D.G. Time Series and Postglacial Forest Ecology. Quat. Res. 1981, 15, 265–277. [Google Scholar] [CrossRef]
- Ford, D.E.; Thornton, K.W. Time and length scales for the one-dimensional assumption and its relation to ecological models. Water Resour. Res. 1979, 15, 113–120. [Google Scholar] [CrossRef]
- Vandermeer, J. Coupled oscillations in food webs: Balancing competition and mutualism in simple ecological models. Am. Nat. 2004, 163, 857–867. [Google Scholar] [CrossRef] [PubMed]
- Benincà, E.; Huisman, J.; Heerkloss, R.; Jöhnk, K.D.; Branco, P.; van Nes, E.H.; Scheffer, M.; Ellner, S.P. Chaos in a long-term experiment with a plankton community. Nature 2008, 451, 822–825. [Google Scholar] [CrossRef] [PubMed]
- Benincà, E.; Jöhnk, K.D.; Heerkloss, R.; Huisman, J. Coupled predator–prey oscillations in a chaotic food web. Ecol. Lett. 2009, 12, 1367–1378. [Google Scholar] [CrossRef] [PubMed]
- Nyquist, H. Certain Topics in Telegraph Transmission Theory. Trans. Am. Inst. Electr. Eng. 1928, 47, 617–644. [Google Scholar] [CrossRef]
- Dormand, J.R.; Prince, P.J. A family of embedded Runge–Kutta formulae. J. Comput. Appl. Math. 1980, 6, 19–26. [Google Scholar] [CrossRef]
- Koren, I.; Feingold, G. Aerosol-cloud-precipitation system as a predator–prey problem. Proc. Natl. Acad. Sci. USA 2011, 108, 12227–12232. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Stenseth, N.C.; Krebs, C.J.; Zhang, Z. Linking climate change to population cycles of hares and lynx. Glob. Chang. Biol. 2013, 19, 3263–3271. [Google Scholar] [CrossRef]
- Stenseth, N.C. Canadian hare–lynx dynamics and climate variation: Need for further interdisciplinary work on the interface between ecology and climate. Clim. Res. 2007, 34, 91–92. [Google Scholar] [CrossRef]
- Snyder, W.E.; Chang, G.C.; Prasad, R.P. Conservation Biological Control: Biodiveristy Influences the Effectiveness of Predators. In Ecology of Predator–Prey Interactions; Oxford University Press: Oxford, UK, 2005; p. 324. [Google Scholar]
- Hunsicker, M.E.; Ciannelli, L.; Bailey, K.M.; Buckel, J.A.; Wilson White, J.; Link, J.S.; Essington, T.E.; Gaichas, S.; Anderson, T.W.; Brodeur, R.D.; et al. Functional responses and scaling in predator–prey interactions of marine fishes: Contemporary issues and emerging concepts. Ecol. Lett. 2011, 14, 1288–1299. [Google Scholar] [CrossRef] [PubMed]
- Weitz, J.S.; Levin, S.A. Size and scaling of predator–prey dynamics. Ecol. Lett. 2006, 9, 548–557. [Google Scholar] [CrossRef]
- Cortez, M.H. Comparing the qualitatively different effects rapidly evolving and rapidly induced defences on predator–prey Interactions. Ecol. Lett. 2011, 14, 202–209. [Google Scholar] [CrossRef] [PubMed]
- Kalinkat, G.; Schneider, F.D.; Digel, C.; Guill, C.; Rall, C.B.; Brose, U. Body masses, functional responses and predator–prey stability. Ecol. Lett. 2013, 16, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Schneider, F.D.; Scheu, S.; Brose, U. Body mass constraints on feeding rates determine the consequences of predator loss. Ecol. Lett. 2012, 15, 436–443. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabella, B.; Meloni, F.; Martinez, A.S. Inadequate Sampling Rates Can Undermine the Reliability of Ecological Interaction Estimation. Math. Comput. Appl. 2019, 24, 48. https://doi.org/10.3390/mca24020048
Cabella B, Meloni F, Martinez AS. Inadequate Sampling Rates Can Undermine the Reliability of Ecological Interaction Estimation. Mathematical and Computational Applications. 2019; 24(2):48. https://doi.org/10.3390/mca24020048
Chicago/Turabian StyleCabella, Brenno, Fernando Meloni, and Alexandre S. Martinez. 2019. "Inadequate Sampling Rates Can Undermine the Reliability of Ecological Interaction Estimation" Mathematical and Computational Applications 24, no. 2: 48. https://doi.org/10.3390/mca24020048
APA StyleCabella, B., Meloni, F., & Martinez, A. S. (2019). Inadequate Sampling Rates Can Undermine the Reliability of Ecological Interaction Estimation. Mathematical and Computational Applications, 24(2), 48. https://doi.org/10.3390/mca24020048