The following subsections explore and apply this judicial tree construction in several different ways. First, the tree associated to a handful of natural courts in the modern SCDB is depicted and discussed; I try to highlight the interesting features of these trees and use this as an opportunity to illustrate how I believe judicial trees can be interpreted (
Section 4.1). Next, I provide a tree-based alternative to the usual notion of a median justice based on MDS coordinates—one that I argue is more faithful to the idea of political centrality (
Section 4.2). I then split the judicial tree for a natural court into a forest, one tree for each issue area (civil rights, due process, etc.), to try to disentangle some of the factors that go into the tree’s overall shape (
Section 4.3). By computing trees for a natural court one case at a time, a dynamic evolution of the Court can be provided; this idea is illustrated by walking the reader through some of the key cases that shaped the 2010-2016 Roberts 4 Court (
Section 4.4). Finally, matrix completion is used to construct a massive judicial tree with every Supreme Court justice who has served since 1946 (
Section 4.5).
4.1. Snapshots of the Court
Here I look at, and comment on, a few particularly intriguing judicial trees to show the diversity of configurations that have occurred over the years and to practice interpreting them. In each case the method described in
Section 3 is used: the
majority variable in the SCDB is used to calculate voting disagreement rates for all pairs of justices in a given natural court (based on all cases heard jointly by each pair of justices, not just the cases during the given natural court) then the ordinary least squares minimizing phylogenetic tree is computed and plotted with the justices colored by the one-dimensional MDS coordinates coming from the same disagreement rates, and with the major axis (the path between the most frequently disagreeing justices) in bold. The most recent natural court in the SCDB (know as the Roberts 7 Court) is depicted in
Figure 2, so we’ll proceed reverse chronologically from there.
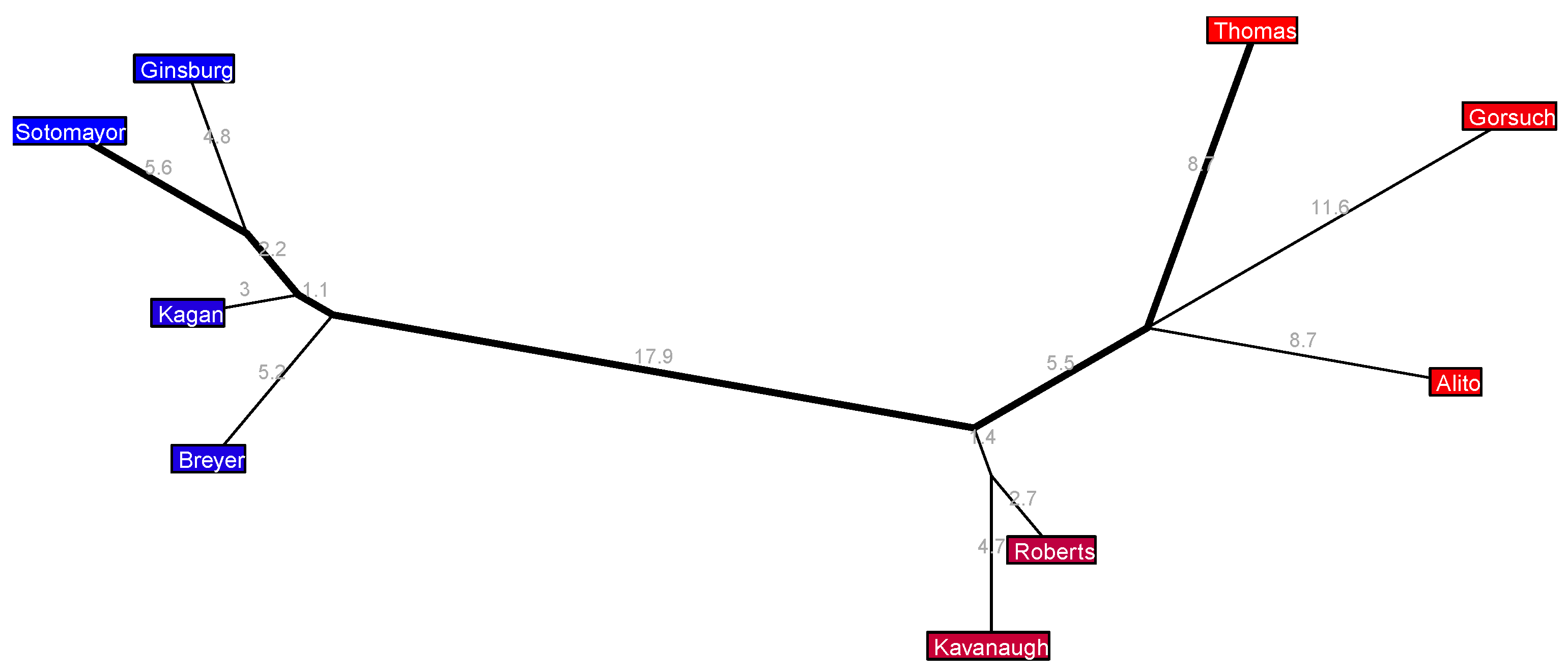
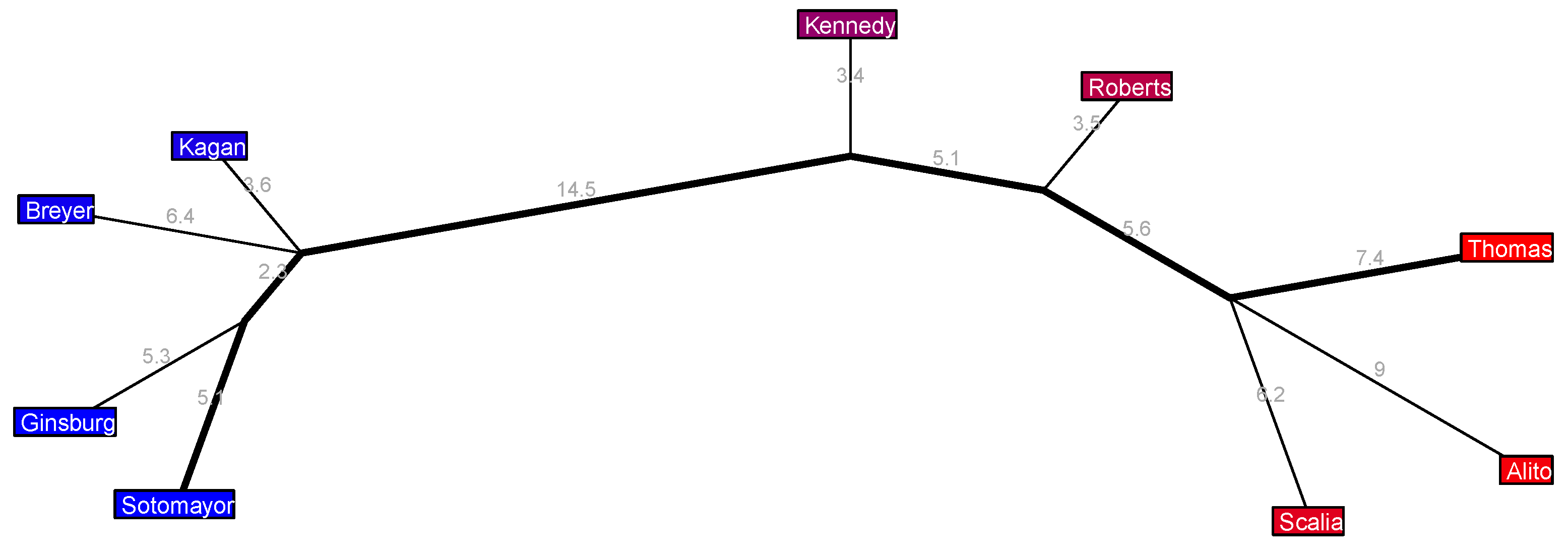
The long 2010–2016 Roberts 4 Court in
Figure 6 has a clear liberal wing with Sotomayor paired up with Ginsburg and Kagan paired up with Breyer (each pair only voting against each other about 10% of the time); a conservative wing with Thomas, Alito, and Scalia (with approximate disagreement rates among them ranging from 13.6% to 16.4%); Kennedy, a notorious swing vote during this era, sits near the center, veering ever so slightly to the right, with only a small deviation from the major axis, and a little further along to their right sits Roberts, who also only extends from the major axis by a small amount and who also is known for some important swing votes during these years—most notably, perhaps, when he surprised many pundits by siding with the liberals in the 5-to-4 case
National Federation of Independent Businesses v. Sebelius (2012) upholding the individual mandate of Obama’s Affordable Care Act on the grounds of the Constitution’s Taxing and Spending Clause.
The 1993–1994 Rehnquist 6 Court in
Figure 7 has a clear conservative coalition of five justices and a liberal coalition of four justices. There are two things that I find striking here. First, O’Connor, an influential swing voter [
39], extends from the major axis at the exact same center-right location as the swing voting Kennedy—yet there is a substantial distance between them (14.9%) and no common edge connecting them to the major axis as there was with Roberts and Kavanaugh in
Figure 2. This suggests that these two swing-voting justices are united in their political orientation but that their frequent disagreements with the rest of the conservative coalition tend to occur in distinct directions from each other, meaning they do not have much ideological common ground aside from their center-right political position. Second, the four-justice liberal coalition is remarkably spread out: these justices are all distant from the conservative justices, but additionally there are considerable distances between each pair of liberal justices—much more so than between the conservatives here and much more so than between the liberals in the two previous trees we have looked (
Figure 2 and
Figure 6). The liberal justices here seem to be frequently disagreeing with each other in ways more complicated than just degree of political extremity—suggested in the tree by the relatively large distances between them in contrast to the more modest separation between their political positions along the major axis.
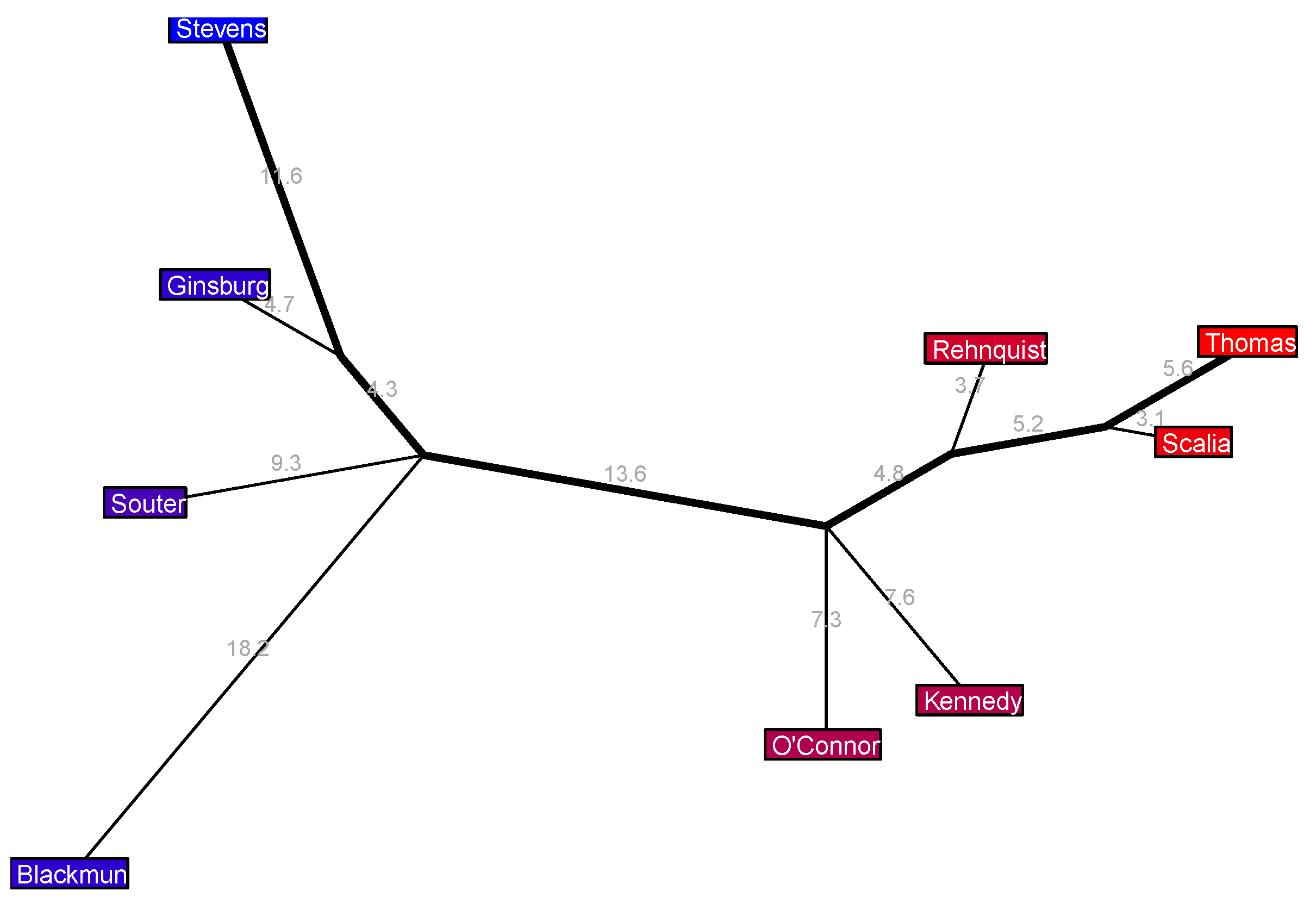
The 1975–1981 Burger 6 Court in
Figure 8 is quite remarkable: the Court has essentially fractured into a tripolar configuration. Marshall and Brennan are closely united on the liberal side, and joining them from a considerable distance at a center-left political position is Stevens—who not only is a sizable distance of 11.9% to the right of them, but he also extends a sizable 10.5% from the major axis, indicating disagreements with the liberals that occur in both the standard political direction and an additional unspecified direction. On the right we have a fairly standard configuration of three conservative justices, but quite unusually in the center of the Court we have a trio of justices—White, Blackmun, and Stewart—who all join the major axis at the same exact location yet the approximate disagreement rates between them are rather large, ranging from 24.7% to 25.6%. I think it would be fascinating to look further into the ideology and judicial behavior of these three centrist justices to see what sets them apart from the rest of the Court and from each other, but a deep dive into the cases they have voted on is beyond my expertise.
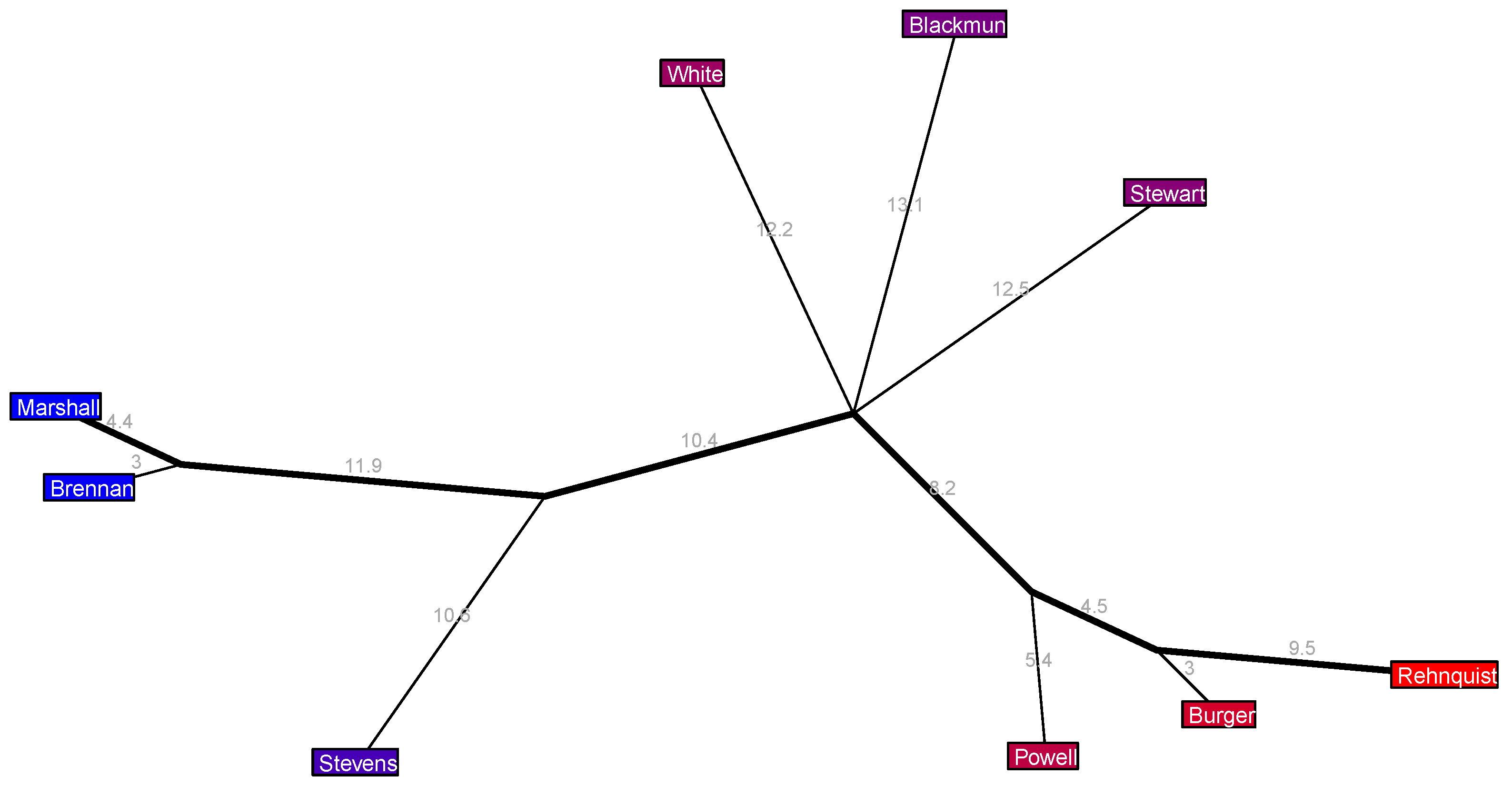
I am including the 1967–1969 Warren 10 Court in
Figure 9 as one of our snapshots mostly because it is so bizarre. The three conservative justices are easy to recognize, but beyond this things are quite scrambled. Douglas is the most liberal justice here (and indeed he was once called “the most doctrinaire and committed civil libertarian ever to sit on the court” [
40]), but one step to their right along the major axis is a node sprouting out five center-left justices. Two of these, Warren and Black, are each on their own edge leading directly to the major axis (and Black’s length 16.4% edge is sizable!), while the other three justices of this center-left quintet—Marshall, Brennan, and Fortas—form a curious trio who are all very close to each and separated from the major axis by a sizable length 8.8% edge. The interpretation here is that these five center-left justices all have the same political position and their frequent disagreements occur in three different directions orthogonal to this liberal-conservative axis: one direction for Warren, a separate direction for Black, and a further separate direction that is common to all of Marshall, Brennan, and Fortas. It is difficult to figure out what these directions could be, but if nothing else this is an example where there is clearly much more to the story than one-dimensional ideal point estimation can convey.
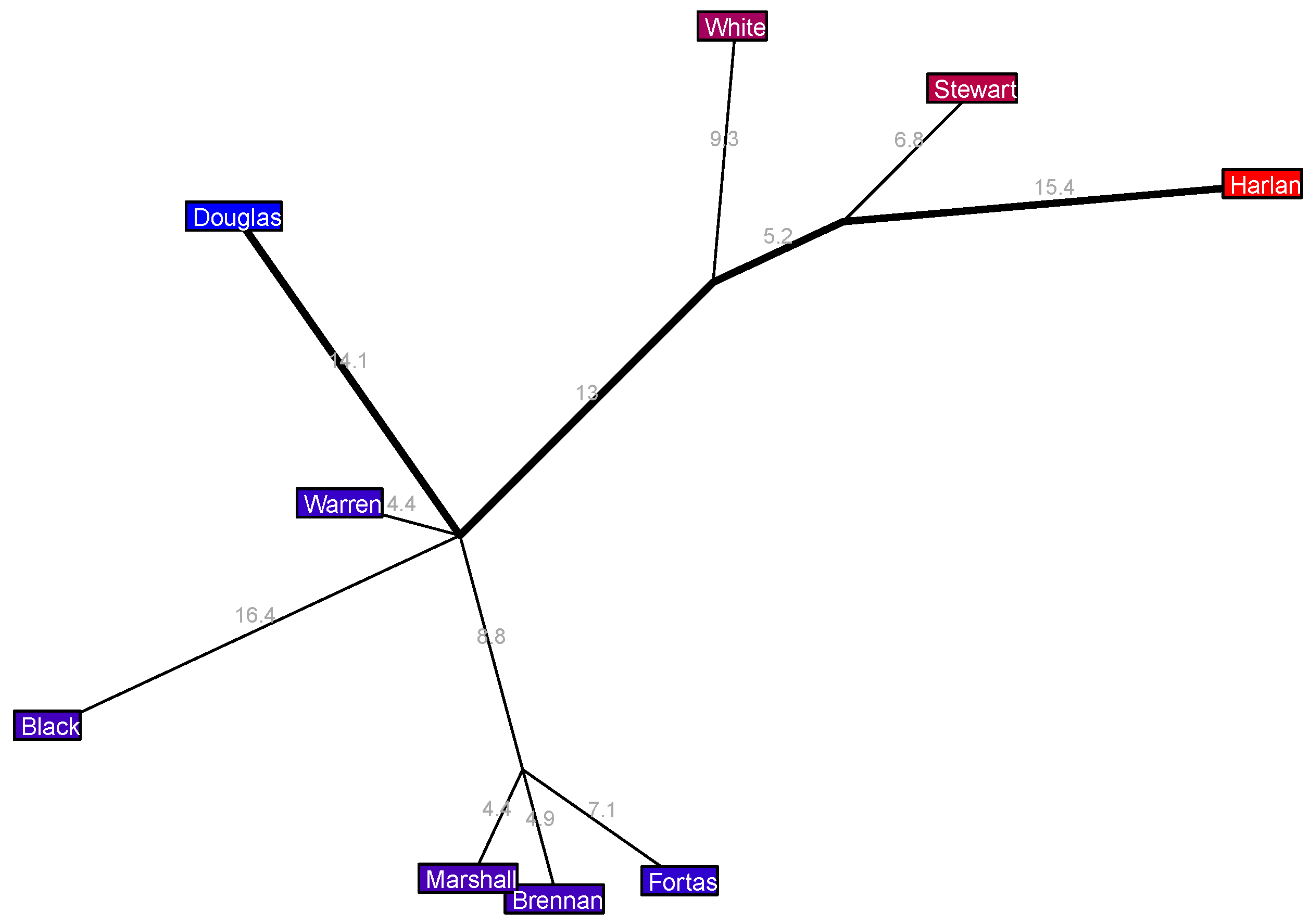
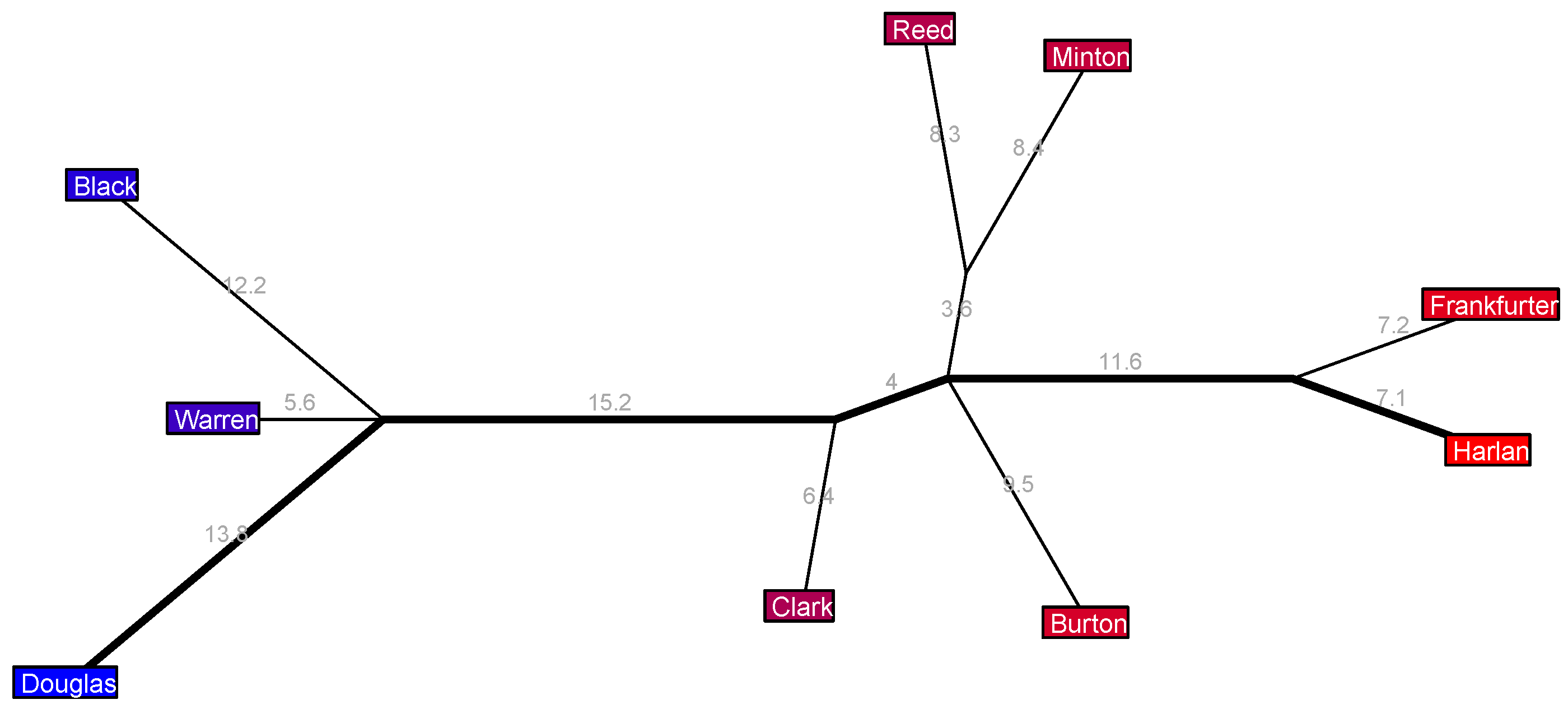
In the 1967–1969 Warren 10 Court in
Figure 10 we have a clear liberal coalition of three justices, a clear far-right pair of Frankfurter and Harlan, a centrist Clark, but then the center-right trio of Reed, Minton, and Burton is intriguing: all three of these justices have the same political position, but Reed and Minton are further united by a length 3.6% edge sprouting from the major axis. This suggests that these three justices all have similar center-right political orientations but that there is some ideological common ground uniting Reed and Minton that is not shared by Burton.
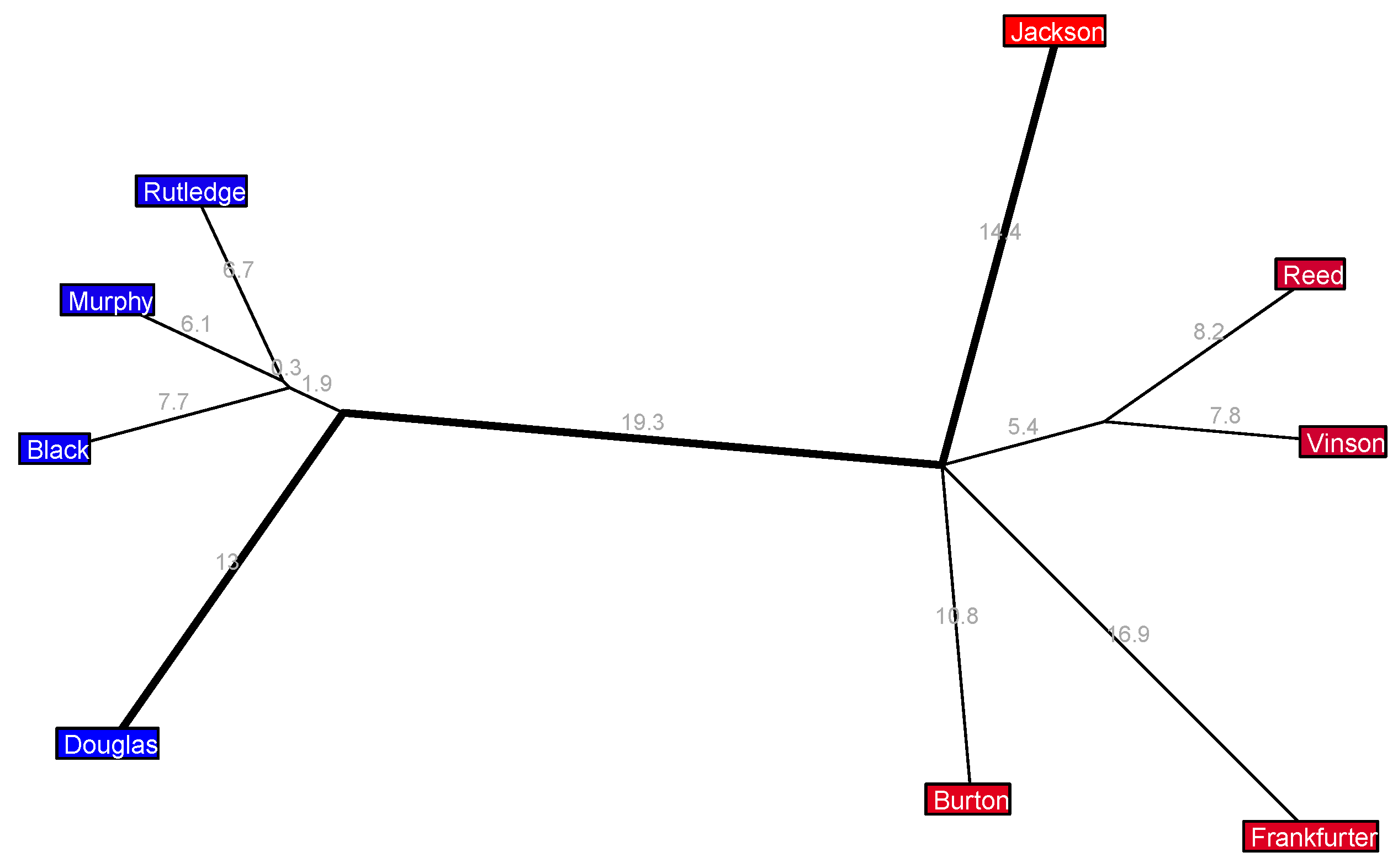
Finally, going all the way back to the 1946–1949 Vinson 1 Court, we see in
Figure 11 a very strongly bipolar layout more typical of recent times. There is a fairly cohesive four-justice liberal coalition and a less cohesive five-justice conservative coalition and a sizable gap between the two coalitions.
4.2. Central Justices
The related though meaningfully distinct notions of the “median justice,” the “power” of a justice, and a “swing vote” have interested scholars for many years. The field of empirical legal studies has produced a range of data-driven methods for quantifying and studying these concepts [
4,
5,
15,
16,
25,
32,
39,
41,
42]. The phylogenetic tree perspective of the Court developed in this paper yields a new network-theoretic way of describing which justices are most central to the Court. One particularly novel aspect of this approach is that it looks not just at whether there is an individual justice sitting at an influential central position but also whether there is a pair or even a trio of justices who collectively play the central role. It also attempts to extract political centrality from voting agreement rates, which as we have discussed earlier is not quite the same as measuring centrality directly from the one-dimensional MDS ideal point estimates. Let me start with the formal definition.
Definition 6. In the tree estimated for a given nine-justice natural court, if there is a justice j such that removing the node connecting j to the major axis results in two connected components not containing j, each with four justices, then j is central in this natural court. If there is a pair of justices, , that are connected to the major axis at the same node and removing this node results in two connected components not containing them, one with four justices and one with three justices, then the pair is central. If there is a triple of justices connected to the major axis all at the same node and removing this node results in two connected components not containing these justices, each with three justices, then the triple is central.
This is a lot to process, but it’s really just capturing whether there are justices who sit as the median of the Court as measured by political position along the major axis. Indeed, the definition can be rephrased as follows: j is central if four justices have political positions to the left and four justices have political positions to the right; are central if their political positions coincide and there are three justices with political positions on one side and four on the other; are central if their political positions coincide and there are three justices on either side of them in terms of political position. You might recall that I cautioned the reader from putting too much import on political position near the ends of the spectrum; fortunately this does not impact this notion of centrality which really only depends on the overall balance of justices on either side rather than the detailed order and position of the justices at the ends.
The usual definition of a median justice is that their one-dimensional ideal point estimate has four justices on either side, so this tree-based definition is replacing one-dimensional ideal point estimates with political position along the major axis (fitting with the theme of this paper that the major axis might better capture the liberal-conservative spectrum than the
x-coordinate in MDS) while simultaneously allowing for “ties” because the nature of trees means that justices often share political positions with each other and express their disagreements in other directions captured by branches extending from the major axis. Scholars have defined a “super median” justice by measuring the gap in one-dimensional ideal point estimates between the median justice and the nearest neighbors [
16]—a justice who sits in the middle of the Court with a large gap from them to the liberal and conservative coalitions tends to play an outsize role as the pivotal swing voter—and if desired one could immediately adapt this notion to tree-based centrality as well. Let us take a look at some examples by inspecting the judicial trees pictured so far.
In
Figure 2, Roberts and Kavanaugh form a central pair. I think of this as follows. If the court behaved in a purely one-dimensional political way, and if the major axis does reflect this political axis, then depending on the political focal point of the case we’d expect Roberts and Kavanaugh to either both side with the liberals or both side with the conservatives, and either way they are guaranteed to supply the majority as they do so. In other words, politically they should swing together. Of course, the court does not behave in a purely one-dimensional way, but in more than one dimension there is not really a notion of median justice (for instance, in [
5] medians are computed separately for the two axes), so my tree-based definition here is attempting to capture and convey how swing voters would behave if everything were driven by one-dimensional politics.
Figure 6 is simpler: Kennedy is the lone central justice. The strength of their position here, in the sense of super medians, is determined by their nearest neighbor, namely Roberts. This makes sense: if Roberts were more firmly allied with the other three conservative justices then Kennedy could freely swing the majority in political cases as he pleases, but Robert’s proximity to Kennedy means that there will still be cases where Roberts swings over to the liberal side (as we saw with the aforementioned case on the Affordable Care Act) and when this happens the liberals have a majority of five votes already so Kennedy is unable to swing the majority back over to the conservatives.
In
Figure 7, we have Kennedy and O’Connor as a central pair, which fits a common perception of them both as influential swing-voting justices [
39].
The curious tripolar judicial tree in
Figure 8 has White, Blackmun, and Stewart as a central trio—and a quite strong one due to the large gaps between their political position and that of the other justices on either side of them.
The scrambled tree in
Figure 9 has no central justice(s)—and indeed even by inspecting this tree manually it is difficult to tell where the balance of power lies in it. Note that by contrast one-dimensional ideal point estimation forces a median justice to always exist, which in my opinion does not make much sense here given how multidimensional and bizarrely balanced this Court is.
Figure 10 is far less scrambled and more straightforward than the previous one, yet it too has no central justice(s). Clark is not central because siding with the liberals is not enough to give them the majority, and the Reed-Minton-Burton trio is not central because they have two justices to their right and four to their left rather than three and three. I must admit that my definition of central here seems a bit questionable because this trio could grant the majority to either side by swinging all together (since two plus three reaches the crucial threshold of five) so perhaps one might want to enlarge the definition of a central trio to include these 4-3-2 configurations, but I will resist doing so because that just does not seem central enough to merit the name.
Figure 11 is actually the same tree topology as the previous figure (the only difference is the edge lengths), so here too there is no central justice(s).
The central justices for all the nine-justice natural courts in the modern SCDB are depicted below in
Table 1.
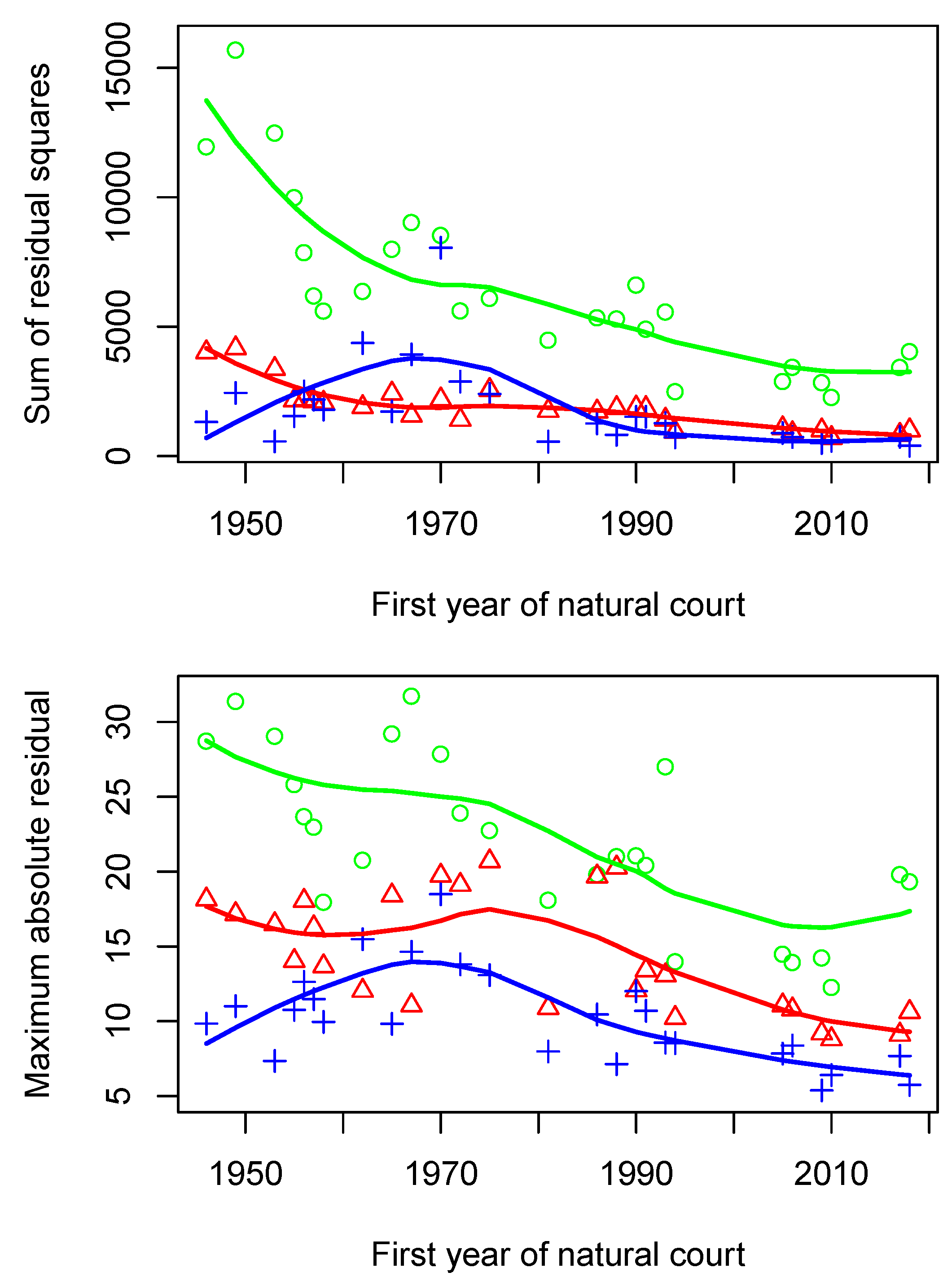
It is curious that all the natural courts after 1972 have at least one central justice, whereas only four of the prior eleven do. In general, the first half of these natural courts seem to exhibit more unusual balances of power and chaotic looking configurations whereas throughout the second half of these natural courts the structure seems to settle down more and more. This was seen in some of the snapshots from
Section 4.1, and it’s also seen in
Figure 5 where for instance all three methods (one-dimensional MDS, two-dimensional MDS, and tree estimation) have maximal absolute residues that steadily trend downward from the 1970s onward. This settling of the Court over the past fifty years is also one of the main topics I look into quantifying in the sequel paper [
9].
The names in this table of central justices have a lot of overlap with the other empirical methods suggested in the literature for measuring median and swing justices (it is interesting to compare in particular with (Table 4 in [
15]) where there is considerable though not complete agreement with the year-by-year analysis provided by Martin-Quinn scores). There is no way to say which method is “best” because there is no underlying truth to compare to; instead, each method gives a slightly different perspective on the issue. That said, it is reassuring that the different methods do not give wildly varying results.
4.3. Issue Areas
The SCDB includes a variable indicating the “issue area” of each case. The values of this variable are somewhat subjective and far from unequivocal—often there are multiple issues involved in a case and it is not clear how to choose just one of them, and sometimes the main issue in a case does not neatly align with one of the 14 possible values of this variable. Nonetheless, in the aggregate this provides a useful way to divide the cases. For a given natural court, one can produce a separate judicial tree for each different issue area, although for some issue areas there will not be enough cases to have rich disagreement rates among the justices, especially between the justices who did not overlap on the Court for very long. Let us take a look (see
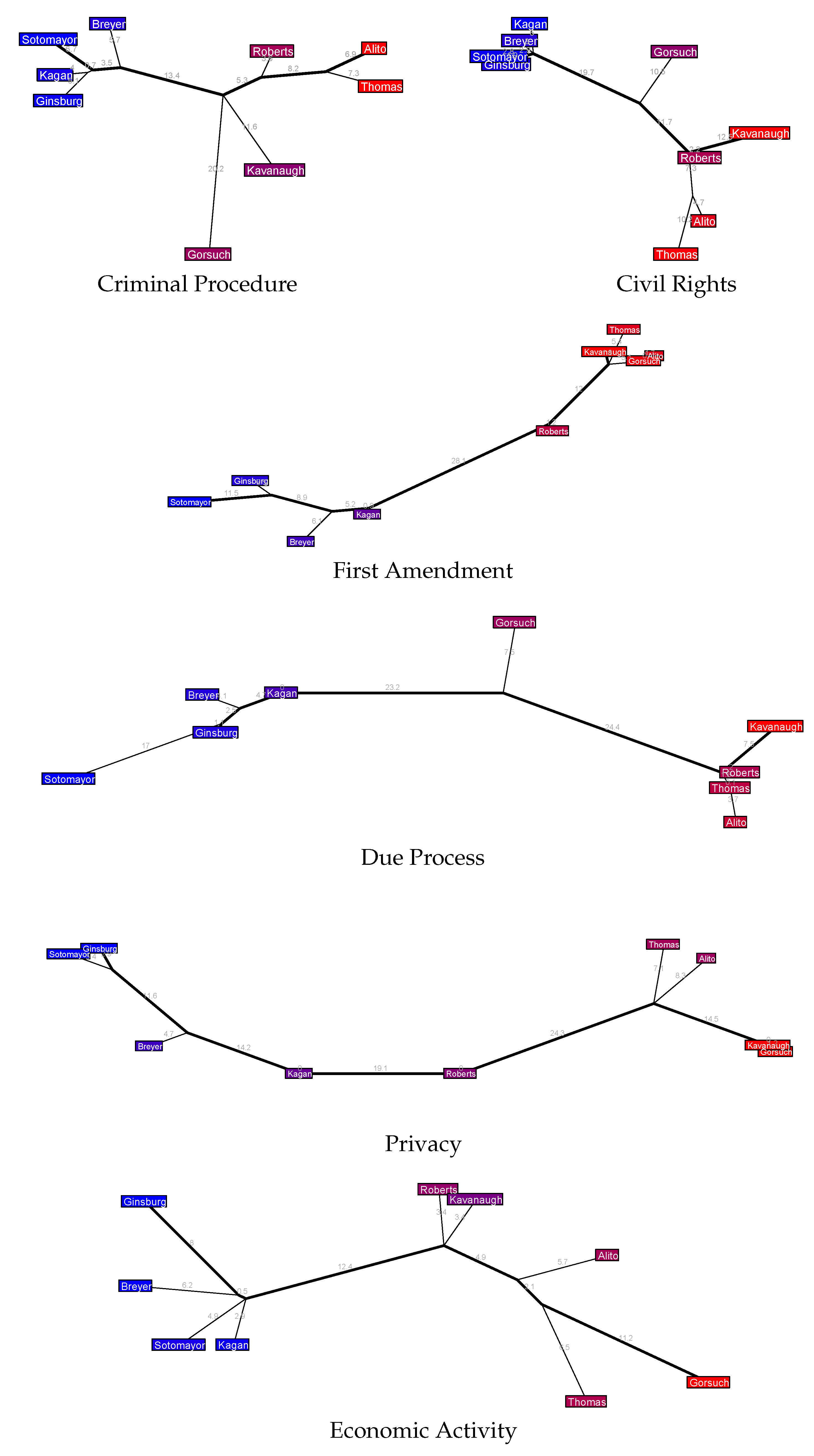
Figure 12 and
Figure 13) at some of the interesting trees in the issue area forest for the most recent natural court in the SCDB, the 2018–2020 Roberts 7 Court pictured in
Figure 2.
Criminal procedure cases look fairly similar to the full judicial tree in
Figure 2 except that one prominent difference is Gorsuch has moved from the Court’s far right to its center, where he extends a considerable distance from the major axis. Furthermore, Kavanaugh has moved from their center-right position with Roberts and now sits at the center with Gorsuch in terms of political position, though he shows no common ground with Gorsuch beyond this. Turning to Civil Rights cases, the liberals have not changed much but the conservatives have some intriguing positions: here Kavanaugh is a far-right justice whereas Gorsuch is centrist, and curiously Roberts is pretty far to the right in terms of political position but as their purple shading indicates he is still center-right in the one-dimensional MDS estimate.
What I find most striking about the First Amendment cases is that there is very little extension off the major axis anywhere in the tree. Based on the earlier discussions about what the major axis means, I interpret this as suggesting that these justices’ votes in First Amendment cases are very much captured by one-dimensional politics. This is mostly true of Due Process cases as well, though to a lesser extent. It is also interesting to note that in Due Process cases we have Gorsuch at the Court’s center, leaning just slightly to the right, whereas Kavanaugh is again the far right justice in these cases (as he was with Civil Rights and First Amendment cases), which is quite distinct from their overall center-right position in this natural court. Privacy cases are also mostly one-dimensional, the main small exception being some branching off the major axis by Thomas and Alito, suggesting that the small amount of higher-dimensional disagreement that occurs in these cases is among the different conservative viewpoints.
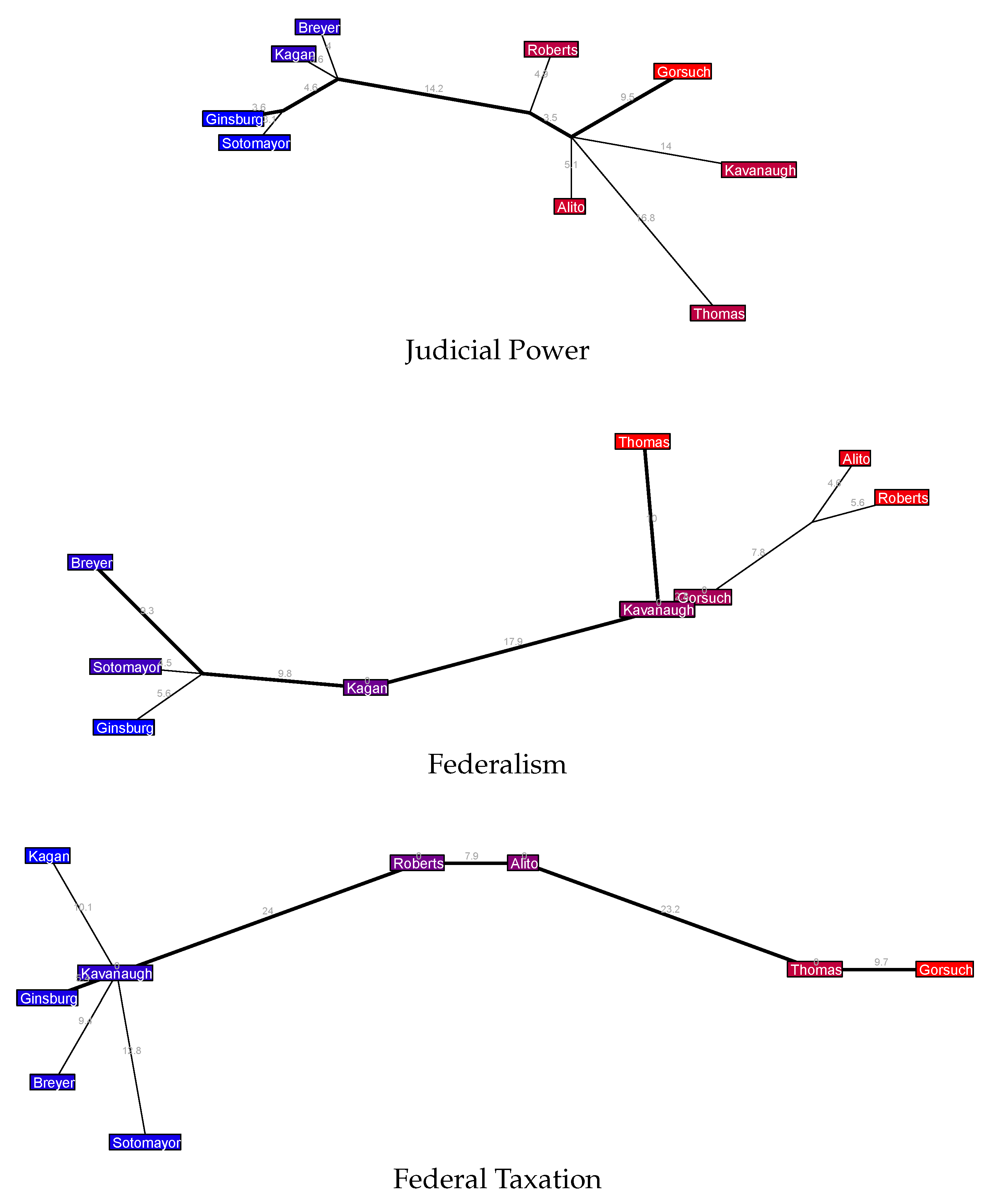
The tree for Economic Activity cases is very similar to the overall tree for this natural court, though curiously MDS seems to consider Alito and Thomas closer to the center of the Court here than the tree does (and also closer to the center than MDS does when combining all case issue areas). The tree for Judicial Power cases shows a fairly tight cohesion and little branching from the major axis among the liberal justices—but the conservative justices are fairly spread out, and mostly in non-political dimensions (by which, as usual, I mean dimensions beyond the primary liberal-conservative spectrum measured by the major axis). The Federalism tree also shows an interesting schism on the Court’s right side: Thomas, Alito, and Roberts are all colored staunchly conservative by the MDS estimate, but there is a sizable disagreement rate between Thomas and the pair of Alito-Roberts, who vote together much more often. The tree for Federal Taxation cases is quite striking: there is almost complete unidimensionality from the Court’s center justices (Roberts and Alito) to its right-most justices (Thomas and Gorsuch), whereas on the left we have a fractured liberal coalition that branches into many directions and which, perhaps most surprisingly, includes Kavanaugh at its center.
Can we use these issue area trees to help understand why Roberts and Kavanaugh have some “common ground” in the original
Figure 2 in the sense of the common edge that connects them to the major axis? I am not sure, to be honest, but this seems like a question worth pursuing in the future.
4.4. Evolution of the Court
Having seen the phylogenetic snapshots of various natural courts, a question that inevitably comes to mind is how this structure emerges—or, put another way, how natural courts evolve over time. One way to approach this is as follows. We can compute disagreement rates for the justices in a natural court one case at a time, using only cases heard during that natural court, and each time we update the disagreement rates we can compute a new judicial tree. At first the tree will vary wildly because each new case has a sizable impact on the disagreement rates, but over time the rates settle down and the tree converges to its mature state. This final state of the tree is not quite the same as the snapshots pictured earlier because those used disagreement rates based on all the cases heard by each pair of justices, not just the cases during the given natural court, but nonetheless this iterative approach gives a reasonable sense of how judicial trees take shape. This process is more interesting for the longer natural courts, so to illustrate it I will apply it here to the 2010–2016 Roberts 4 Court from
Figure 6.
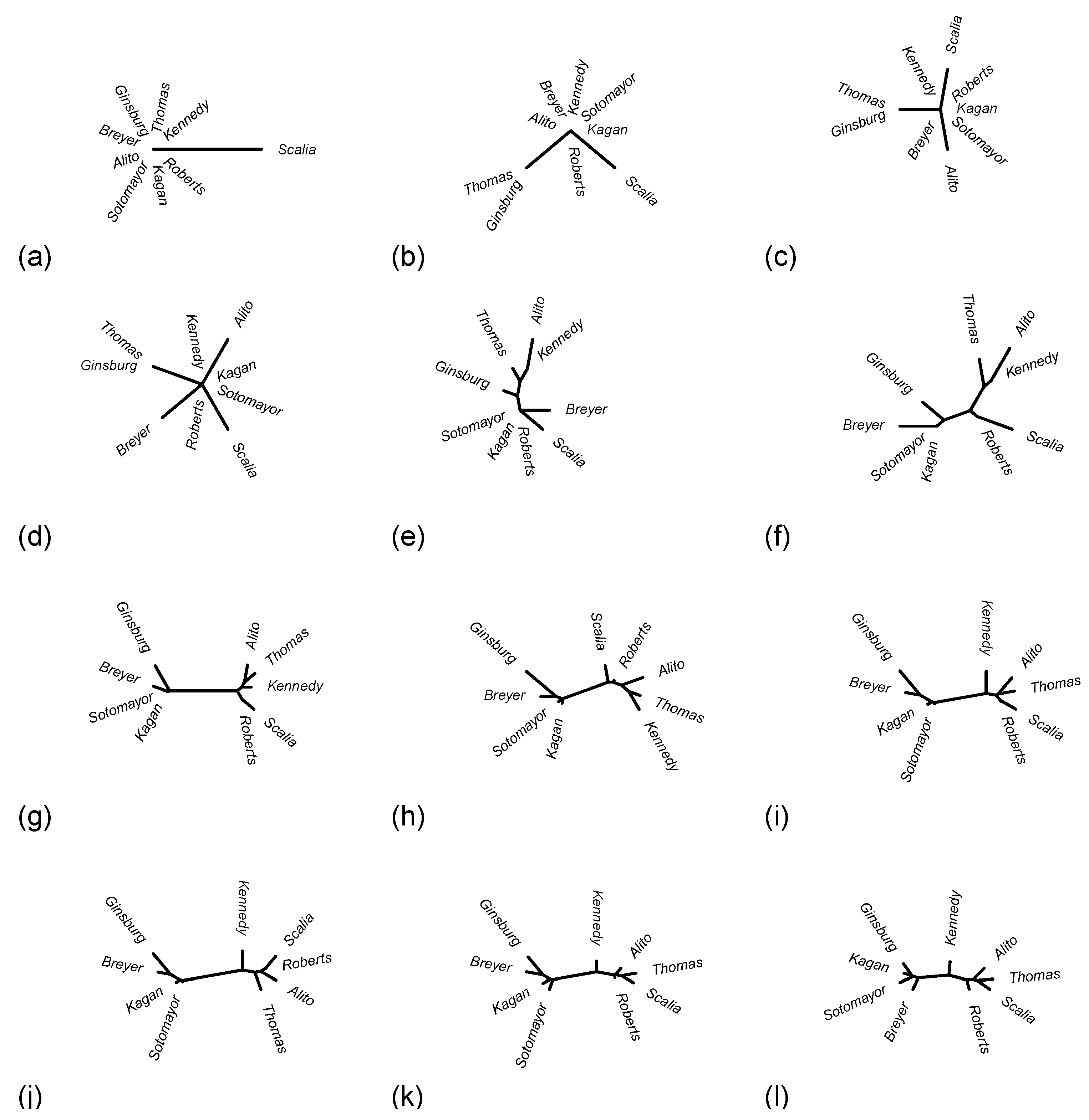
Often the judicial tree does not change substantively as additional cases are included one at a time, or justices may swap positions back and forth multiple times before settling down, so I will just highlight the cases where interesting changes occur to the tree that seem more than just temporary vacillations. Admittedly this is a subjective matter, but that’s ok; I view this process mostly as a tool for uncovering interesting cases that help tell the story of each natural court’s development. The highlighted judicial trees are depicted below in
Figure 14; they do not have all the additional annotations that I normally include because they would not be very readable on such small illustrations of these rather compact trees (and some of them, such as the MDS-coloring of the justice labels, do not really make sense early on). In discussing how the tree changes at each step of this process, I will draw from the legal summaries in Oyez [
43] to help describe the cases that led to these junctures in the tree’s evolution.
The case
Ransom v. FIA Card Services, N.A. (2011) resulted in an 8-to-1 decision, with Kagan writing her first opinion for the Court and Scalia providing the lone dissent. At issue here was whether the projected disposable income for a Chapter 13 bankruptcy plan could allow a vehicle ownership deduction only if the debtor was making payments on the vehicle. The majority opinion ruled that this was indeed so. Scalia’s dissent stemmed from a disagreement over the interpretation of a particular word in the Bankruptcy Abuse Prevention and Consumer Protection Act of 2005. He questioned the Court’s view that a word in a statutory text that does not seem to add any meaning should be construed in a manner so that it does, quipping: “The canon against superfluity is not a canon against verbosity.” This 8-to-1 vote, since it is the first non-unanimous case in the natural court under consideration (according to the order of the cases in the SCDB) results in the phylogenetic tree in
Figure 14a where a single edge connects the Scalia leaf to a leaf labelled by the other eight justices.
The next change, seen in
Figure 14b, is that Ginsburg and Thomas—an unlikely duo—branch off together from the node that was previously labelled by eight justices. This happened because of the 7-to-2 case
CSX Transportation, Inc. v. Alabama Department of Revenue (2011), which had a majority opinion by Kagan and a dissent by Thomas that was joined by Ginsburg. The railroad transportation company CSX had challenged the state of Alabama’s sales and use tax on diesel fuel, as a violation of the Railroad Revitalization and Regulatory Reform Act of 1976, but the suit was dismissed by a lower court. The Supreme Court ruled that a railroad company could indeed challenge such a tax that applies to rail carriers while exempting competitors in the transportation industry. The dissent agreed that Alabama’s tax was potentially discriminatory and that if it did single out railroads among general commercial and industrial taxpayers then the CSX suit would be valid, but in the view of Thomas and Ginsburg this particular tax did not satisfy this criterion.
Alito was the next to split off from the central node, as
Figure 14c illustrates, and he did so because of the 8-to-1 case
Snyder v. Phelps (2011). The family of a deceased soldier, Snyder, had been awarded USD 5 million in damages after members of the cult-like Westboro Baptist Church picketed Snyder’s funeral with extremely vulgar and offensive signs. The circuit courts held that this judgement violated the First Amendment’s protection of religious expression, and the Supreme Court’s majority opinion penned by Roberts agreed. Alito’s lone dissent asserted poignantly: “Our profound national commitment to free and open debate is not a license for the vicious verbal assault that occurred in this case.” Breyer subsequently split off from the central node, as
Figure 14d shows, due to the 8-to-1 vote in
Milner v. Department of the Navy (2011) where the Court supported a Freedom of Information Act request for internal Navy documents discussing the effects of explosions at several locations caused by the Navy. Breyer’s lone dissent ominously asserted: “I would let sleeping legal dogs lie.”
The next phylogenetic development that occurred, depicted in
Figure 14e, is more interesting. The 6-to-3 vote in
Skinner v. Switzer (2011) had a rather unusual majority of Breyer, Ginsburg, Kagan, Roberts, Scalia, and Sotomayor. This majority ruled that a convicted prisoner seeking access to biological evidence for DNA testing can assert that claim in a civil rights action under a federal statute on civil action for deprivation of rights. Thomas’ dissent, joined by Alito and Kennedy, stretched the phylogenetic tree out, so to speak, by pulling Kennedy and Thomas out along Alito’s branch and separating Thomas from Ginsburg. While in some sense the tree is really starting to take shape here—for instance, it now has four interior nodes instead of just one—the justices are quite scrambled as the non-unanimous cases leading to this phylogenetic tree have been, for whatever reason, heavily skewed towards oddball (in the sense of [
17]) majority and minority coalitions. In other words, this particular natural court evidently started rather chaotically before eventually settling down into voting patterns that align more closely with political ideology.
Turning now to
Figure 14f, we see a much more familiar state of this Court: a clear liberal branch with Breyer, Ginsburg, Kagan, and Sotomayor and conservative branch with Alito, Kennedy, Roberts, Scalia, and Thomas. The biggest difference between this and the final phylogenetic structure of the Court is that here Kennedy is deeply embedded in the conservative branch. Nevertheless, how did the Court suddenly reach such a mature configuration? The 5-to-4 vote in
Connick v. Thompson (2011) split right down the liberal-conservative divide, with Kennedy joining the conservative majority. It is somewhat striking that this was the 14th case decided by this natural court (again, according to the order in the SCDB) yet the first 5-to-4 decision—although the very next two cases,
Arizona Christian School Tuition Organization v. Winn (2011) and
Cullen v. Pinholster (2011), were both 5-to-4 with the exact same majority as
Connick. The politically divided vote in
Connick was enough to pull the phylogenetic tree apart into two very distinct and ideologically consistent branches, then the immediately following cases
Arizona and
Cullen both extended and solidified this transformation.
In
Figure 14g we have very little movement in the liberal branch, but in the conservative branch Kennedy has effectively swapped positions with Thomas, placing himself closer to the root of the conservative branch. This occurred because of the case
Brown v. Plata (2011), which had a majority consisting of Breyer, Ginsburg, Kagan, Kennedy, and Sotomayor. This was thus also a 5-to-4 case splitting along political lines, but this time Kennedy swung over to the liberals, and he did so while penning the majority opinion. The case concerned a class-action lawsuit alleging that California prisons violated the Eighth Amendment’s ban on cruel and unusual punishment. A special panel of three federal judges determined that serious overcrowding was the primary cause for such a violation and ordered the release of roughly forty-thousand inmates to decrease the inmate population to a number closer to the design capacity of these California prisons. The question before the Supreme Court was whether such a court order was in violation of the Prison Litigation Reform Act, and Kennedy’s majority opinion said no, thereby affirming the special panel in the face of Scalia’s strongly worded objection declaring its order as “perhaps the most radical injunction issued by a court in our Nation’s history.” Kennedy’s swing to the liberals here is reflected phylogenetically by their placement slightly closer to the center of the tree. Since at this point in the Court’s development he has sided with the conservatives in several 5-to-4 cases and only with the liberals in one 5-to-4 case, it makes sense that he is still placed squarely on the conservative branch.
The tree in
Figure 14h is in some sense a step backwards: there is again little movement in the liberal branch, but here Scalia has moved to the root of the conservative branch and Kennedy has moved out onto a more extreme branch with Thomas and Alito. There were several non-unanimous cases since the previously discussed one—namely, an Alito-Ginsburg minority in
Fowler v. United States (2011), a Kennedy-Thomas minority in
Camreta v. Greene (2011), a solitary Kennedy minority in
Global-Tech Appliances, Inc. v. SEB S.A. (2011), and a Breyer-Ginsburg minority in
Board of Trustees of the Leland Stanford Junior University v. Roche Molecular Systems, Inc. (2011)—but the phylogenetic movement here was driven primarily by
Sykes v. United States (2011), which had an oddball minority of Scalia, Ginsburg, and Kagan. The question underlying this case was whether resisting arrest counts as a violent felony under the Armed Career Criminal Act (ACCA). Kennedy wrote the majority opinion affirming this, and Thomas wrote a concurring opinion in which he mostly agreed with the Court but took issue with its interpretation of a particular legal test that does not appear in the ACCA. The oddball nature of the minority is largely explained by the fact that it took the form of one dissent by Scalia and a separate, strongly contrasting dissent by Kagan that was joined by Ginsburg. Scalia inveighed against the Court for repeatedly attempting to distinguish violent felonies from non-violent ones and succeeding only in issuing an “ad hoc judgment that will sow further confusion,” whereas Kagan and Ginsburg disagreed with the majority opinion because the particular petitioner in the case being heard was convicted only of simple vehicular flight rather than any flight offense involving aggressive or dangerous activity.
Next, coming to
Figure 14i, we have Kennedy moving strongly to their rightful place as the gatekeeper to the conservative branch of the Court. This was the result of
Janus Capital Group, Inc. v. First Derivative Traders (2011), which was 5-to-4 splitting down political lines with Kennedy siding with the conservatives,
Davis v. United States (2011), which had a Breyer-Ginsburg minority, and most importantly
J.D.B. v. North Carolina (2011), which was another 5-to-4 along political lines but with Kennedy siding with the liberals. This latter case, where the majority decided that courts should consider the age of a juvenile suspect in determining what “custody” means in the context of Miranda rights, helped solidify Kennedy’s role as a swing vote at the center-right of the Court—and he would remain there for the rest of this natural court’s development.
The trees in
Figure 14j,k,l show only minor jostling among the justices within each of the two main political branches of the Court. These finer movements are the result of a large number of cases and accumulate over the course of years. During this period Breyer establishes himself as the gatekeeper to the liberal branch, thus creating a smaller more strongly liberal branch of female justices, while Kennedy moves closer to the center of the Court and Roberts establishes himself as the gatekeeper to the conservative branch. One case that contributed to Ginsburg, Kagan, and Sotomayor branching off together was the 6-to-3 decision in
J. McIntyre Machinery, Ltd. v. Nicastro (2011) where these three justices formed the minority. The question before the Court was whether a consumer could sue a foreign manufacturer in state court over a product that the manufacturer marketed and sold through an exclusive distributor in the United States. The majority said no, on the grounds of due process rights of the defendant. The dissenting trio found the majority’s view too antiquated and believed it would make it too easy for manufacturers to unfairly protect themselves by strategically utilizing independent distributors.
An important case that pulled Roberts out of the deeply conservative branch populated by Alito, Scalia, and Thomas was the much-discussed
National Federation of Independent Businesses v. Sebelius (2012), mentioned briefly before in this paper, where Roberts joined the liberals—Breyer, Kagan, Ginsburg, and Sotomayor—to form the majority in this salient 5-to-4 case that upheld the individual mandate in the Affordable Care Act. In fact, it was at precisely this point in this natural court’s chronology that the phylogenetic tree reached its final shape shown in
Figure 14l. Indeed, from here onward the edge lengths in the tree adjusted slightly but no substantive changes to the tree occurred. To get a sense of how long exactly it took the phylogenetic tree to settle into its final shape, note that the first non-unanimous case in this natural court,
Ransom, was argued on 4 October 2010, and
National Federation was argued in 26–28 March 2012, whereas the last non-unanimous case argued before this Court,
Montanile v. Board of Trustees of the National Elevator Industrial Health Benefit Plan (2016), was argued on 9 November 2015.
While this phylogenetic view of the evolution of the Court likely does not provide insight into any individual cases, hopefully the reader sees that it can at least point to interesting individual cases where important shifts in the Court occurred—as well as providing an intriguing overview of the general movement of the justices relative to each other throughout the duration of the natural court being analyzed.
4.5. Across the Years
Having seen various uses of phylogenetic trees to study a particular natural court, a question that might come to mind is whether the interrelationships among justices across different natural courts can also be studied. Intuitively, if Justice A decided many cases with Justice B, and Justice B decided many cases with Justice C, then even if Justice A and Justice C never sat on the bench at the same time, it seems reasonable that an estimate of how frequently they would have voted together should be possible. Furthermore, indeed mathematicians have tools for doing exactly this. In this subsection I explore this task by seeking to create a phylogenetic tree with one leaf for each justice who has served since 1946, the first year of the modern SCDB. It should be noted, however, that the methods described below apply to any collection of justices, so scholars interested in seeing how smaller groups of justices are interrelated can certainly do so using the same techniques.
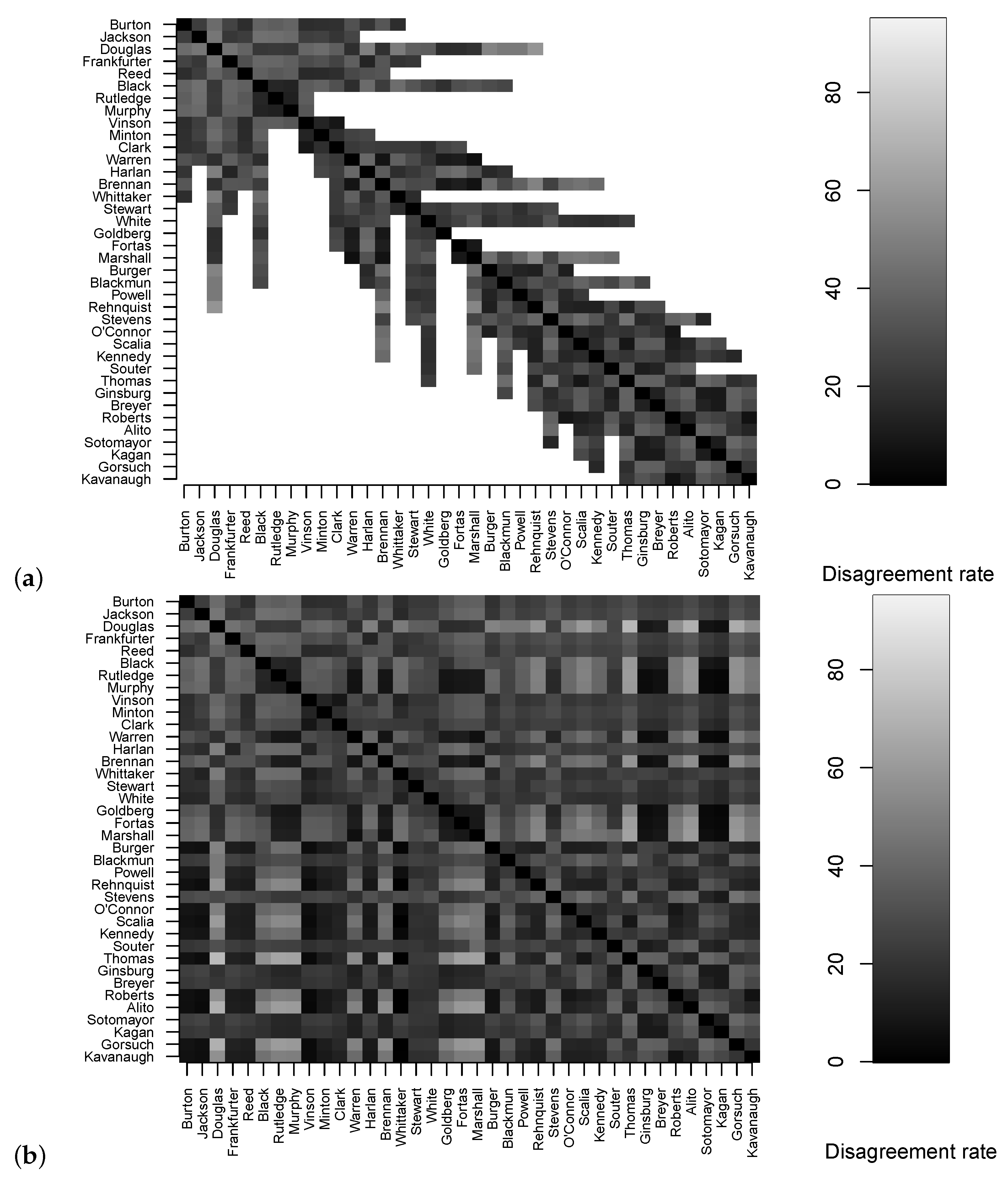
The disagreement rates
used to estimate judicial trees can be viewed as entries of a symmetric
matrix, where here
is the number of justices since 1946. This matrix, shown in
Figure 15a, has rows and columns indexed by the justices, and the
-entry is missing whenever justice
i and justice
j did not overlap for any cases. Matrix completion and statistical imputation refer to a variety of algorithms for filling in the missing entries of a matrix. Obviously there is no “correct” way to fill in missing entries—guessing how frequently non-overlapping justices would have voted against each other is a hypothetical gambit!—but each algorithm proceeds to fill in the missing entries based on some assumption about how the data is structured or some property of the incomplete matrix that should be preserved or some minimality property that distinguishes the completed matrix among all possible completions, etc. I will just pick a popular general purpose method that uses nuclear-norm regularization to find a low-rank matrix completion (as implemented in the R package
softImpute with the function
softImpute using the SVD algorithm type [
44]): see
Figure 15b. I make no claims that this is the right approach to matrix completion in the present setting, I simply want to use a popular method to see what it leads to.
Now that we have a completed matrix, we can view all the entries as voting disagreement rates (the original ones real, the filled in ones hypothetical) and apply tree estimation to produce a judicial tree as before, though now with 38 justices. Here the least squares tree estimate is computationally much harder to produce than for all the nine-justice trees in this paper, so instead of directly solving the optimization problem I first use the popular Neighbor-Joining algorithm to produce an approximate tree and then apply the heuristic algorithm
optim.phylo.ls in the R package
phytools [
45] to iteratively adjust this tree closer to a least squares minimum until it converges. This process is not guaranteed to result in the global least squares minimum, but for a tree of this modest (by biological standards) size it usually does so, and indeed I always got the same final result when perturbing the initial conditions here, so I suspect it is indeed the global minimum. The resulting judicial tree is pictured in
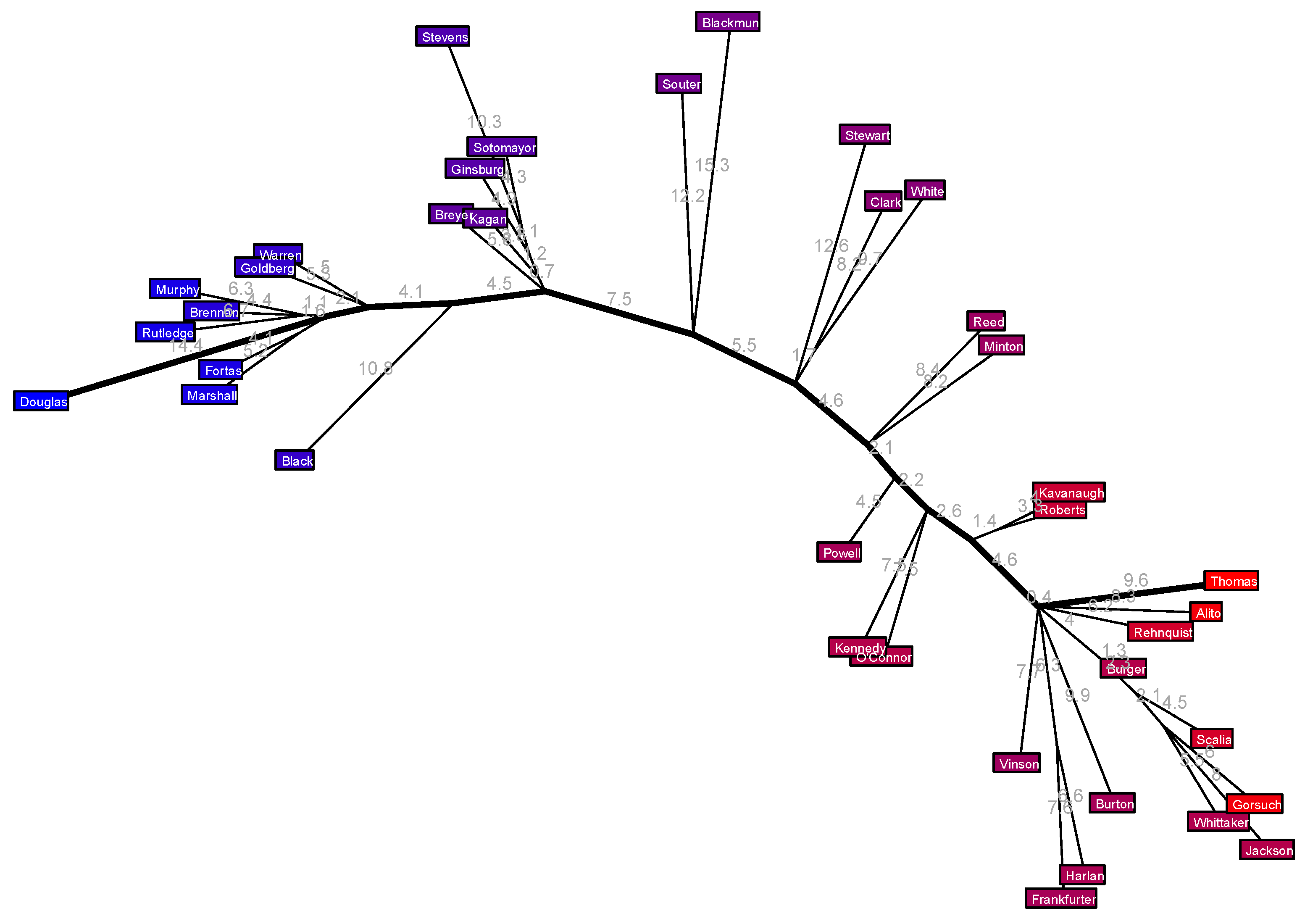
Figure 16.
The (hypothetical) most frequently disagreeing justices, as determined by the matrix completion, who therefore span the major axis in this tree are Douglas on the left and Thomas on the right. Recall that Douglas was earlier described as “the most doctrinaire and committed civil libertarian ever to sit on the court” [
40]. One striking feature of this tree is that the major axis appears quite prominent. There is a lot of extension off it by individual justices, but not many sizable branches carrying multiple justices. I am apprehensive to say that this means the Court’s aggregate behavior over the years has been driven largely by politics, because there is just too many tenuous components to that assertion, but if nothing else this suggests that the few branches we do see in this picture might be interesting and worth investigating. Let us walk through them one at at time. In what follows, I put the years each justice served on the Supreme Court in parentheses.
Starting from the left, we have a length 1.1% edge common to Murphy (1940–1949), Brennan (1956–1990), and Rutledge (1943–1949), and at the same political position along the major axis we have a length 1.6% edge common to Fortas (1965–1969) and Marshall (1967–1991). The two edges are so short that I do not read much into them. However, a bit further along the major axis there is another quintet of justices who share a political position but with a more interesting branching structure. Breyer (1994–) is attached directly at the major axis, then a length 0.7% edge leads to Kagan (2010–) splitting off, then after a length 1.2% edge Ginsburg (1993–2020) splits off, and finally a length 1.1% edge leads to a split between Sotomayor (2009–) and Stevens (1975–2010). If we view the branch point at the major axis as one end of a new axis (a “semimajor” axis, if you will) then Stevens is the opposite end of this axis, a full away, and the other four justices have the following coordinates along this axis: Breyer is 0, Kagan is 0.7, Ginsburg is 1.9, Sotomayor is 3. Is there an ideological interpretation to this axis—meaning something that unites these justices and is roughly quantified by their branch points along it? I do not know. Perhaps it would be an interesting question for a legal scholar to look into. That said, it is curious to note that these five justices almost all served on the Court simultaneously: the earliest departure was Stevens on 29 June 2010, and the latest arrival was Kagan on 7 August 2010. For ten years after that summer of 2010, and for one year prior to it, four of these five justices served concurrently.
The next branch from the major axis occurs where Stewart (1958–1981) extends directly from the major axis but at the same political position Clark (1949–1967) and White (1962–1993) split from each other after a length 1.7% edge. Further along, we have Roberts (2005–) and Kavanaugh (2018–) paired up, but then things get really interesting at the next junction. Ignoring a barely noticeable length 0.4% edge, all the remaining twelve justices split off from the same node. One of these is Thomas (1991–), whose edge is the final segment of the political axis, and four of them branch directly off from the major axis, so let me turn attention to the remaining seven justices. The seven-year overlapping Harlan (1955–1971) and Frankfurter (1939–1962) have paired off together with a considerable length 6.3% edge in common to the major axis. Lastly, there is a curious branch consisting of a length 4% edge after which Burger (1969–1986) splits off, then a length 2.3% edge after which Scalia (1986–2016) splits off, then a length 2.1% edge leading to a three-way split between the chronologically diverse trio of Gorsuch (2017–), Whittaker (1957–1962), and Jackson (1941–1954). Once again, we could think of the furthest justice among these latter five, namely Jackson, as the end of some kind of semimajor axis here in conservative territory, but what this quantifies, and what the sizable length 6.3% common edge to Harlan and Frankfurter represent, are beyond me.
Recall from
Section 2 that Lee in [
31] created an incomplete matrix of the pairwise correlations among the voting vectors for all 36 justices on the Court between 1946 and 2016 and then using methods from statistical mechanics constructed the maximum-entropy model compatible with these correlations. This model produces vote patterns quite similar to what has been observed historically, and in doing so it reveals many interesting phenomena about the Court, especially regarding consensus and formation of dissenting blocs. The author shows that their model, which has around 1000 parameters, fits the data better than a standard model in political science that has over 100,000 parameters. The judicial tree in
Figure 16 is a different kind of model: while it was created from the same data, it does not generate new data than can be compared with the original voting data. Instead, it is a model more in an intuitive sense of a visualization that helps the reader conceptualize the voting behavior of the court (it basically only has as many parameters as edges in the tree, which is 60, or perhaps thrice this if you separately count as parameters the length and two incident vertices for each edge). That said, it is interesting to note that Douglas is the far-left justice in the judicial tree here defining the liberal end of its major axis, and Lee found that Douglas also plays a special role in the maximum entropy model: there are only two negative correlations in the model, and both of them involve Douglas. (A negative correlation means that one justice voting to affirm in a case actually increases the odds of the other justice voting to reverse.) In Lee’s model, Douglas has a negative correlation with both Burger and Rehnquist, both of whom are deeply in conservative territory in my tree model but not especially distinguished in any way that I can see.
Before concluding this discussion of justices across the years, I would like to mention that a different network-theoretic visualization of all justices between the years 1956 and 2005, also based on voting disagreement rates but not involving trees, is shown in Chart 3 on page 253 of [
21].




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}