Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients

 ,
, 
 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Raw Materials
2.3. Proximate Composition
2.4. Fatty Acid Profile and α-Tocopherol Content
2.4.1. Fatty Acid Profile
2.4.2. α-Tocopherol Content
2.5. Phenolic Extracts Preparation
2.5.1. Extraction of Free Phenolic Compounds (FP)
2.5.2. Extraction of Bound Phenolic Compounds (BP)
2.6. Total Phenols (TP) Quantification
2.7. Determination of Free and Bound Phenolic Compounds Using HPLC-DAD-ESI/MS and HPLC-DAD
2.8. Total Antioxidant Capacity (TAC)
2.8.1. DPPH Assay
2.8.2. ORAC Assay
2.8.3. ABTS Assay
2.8.4. Relative Antioxidant Capacity Index (RACI)
2.9. Glycemic Index (GI)
2.10. Color Analysis
2.11. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition
3.2. Fatty Acid Profile (FA) and α-Tocopherol Content
3.3. Determination of Free and Bound Phenolic Content and Composition
3.4. Total Antioxidant Capacity (TAC)
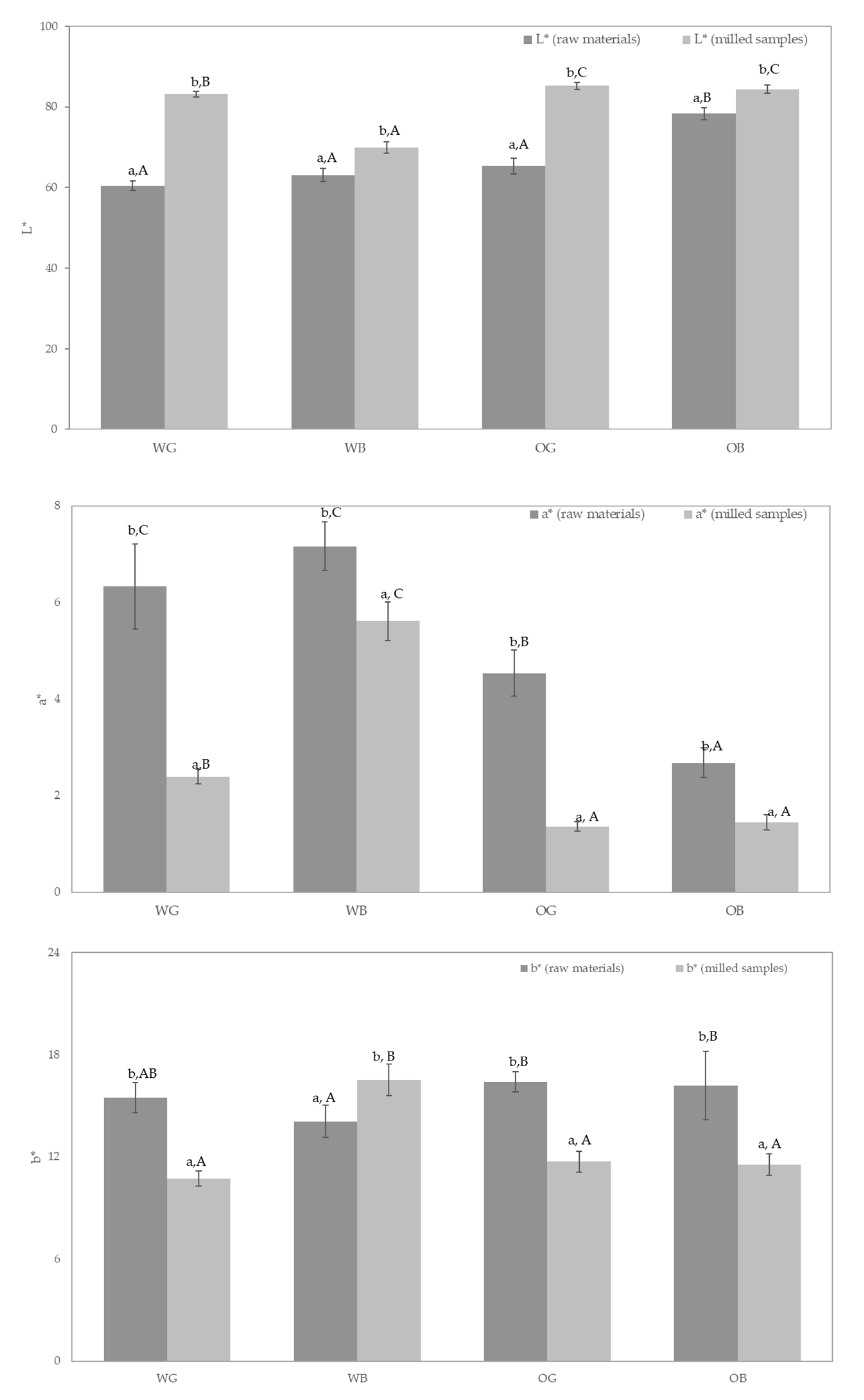
3.5. Color Analysis
3.6. Glycemic Index
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vanegas, S.M.; Meydani, M.; Barnett, J.B.; Goldin, B.; Kane, A.; Rasmussen, H.; Brown, C.; Vangay, P.; Knights, D.; Jonnalagadda, S.; et al. Substituting whole grains for refined grains in a 6-wk randomized trial has a modest effect on gut microbiota and immune and inflammatory markers of healthy adults. Am. J. Clin. Nutr. 2017, 105, 635–650. [Google Scholar] [CrossRef] [Green Version]
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björck, I.; Östman, E.; Kristensen, M.; Mateo Anson, N.; Price, R.K.; Haenen, G.R.M.M.; Havenaar, R.; Bach Knudsen, K.E.; Frid, A.; Mykkänen, H.; et al. Cereal grains for nutrition and health benefits: Overview of results from in vitro, animal and human studies in the HEALTHGRAIN project. Trends Food Sci. Technol. 2012, 25, 87–100. [Google Scholar] [CrossRef]
- He, M.; van Dam, R.M.; Rimm, E.; Hu, F.B.; Qi, L. Whole-Grain, Cereal Fiber, Bran, and Germ Intake and the Risks of All-Cause and Cardiovascular Disease-Specific Mortality Among Women With Type 2 Diabetes Mellitus. Circulation 2010, 121, 2162–2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fardet, A.; Rock, E.; Rémésy, C. Is the in vitro antioxidant potential of whole-grain cereals and cereal products well reflected in vivo? J. Cereal Sci. 2008, 48, 258–276. [Google Scholar] [CrossRef]
- Damsgaard, C.T.; Biltoft-Jensen, A.; Tetens, I.; Michaelsen, K.F.; Lind, M.V.; Astrup, A.; Landberg, R. Whole-Grain Intake, Reflected by Dietary Records and Biomarkers, Is Inversely Associated with Circulating Insulin and Other Cardiometabolic Markers in 8- to 11-Year-Old Children. J. Nutr. 2017, 147, 816–824. [Google Scholar] [CrossRef]
- Lillioja, S.; Neal, A.L.; Tapsell, L.; Jacobs, D.R. Whole Grains, type 2 diabetes, coronary heart disease, and hypertension: Links to the aleurone preferred over indigestible fiber. BioFactors 2013, 39, 242–258. [Google Scholar] [CrossRef] [Green Version]
- Murtaugh, M.A.; Jacobs, D.R.; Jacob, B.; Steffen, L.M.; Marquart, L. Epidemiological support for the protection of whole grains against diabetes. Proc. Nutr. Soc. 2003, 62, 143–149. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Scientific Opinion on the substantiation of health claims related to wheat bran fibre and increase in faecal bulk (ID3066) reduction in intestinal transit time (ID 828, 839, 3067, 4699) and contribution to the maintenance or achievement of a normal body weight (ID 829) pursuant to Article 13 (1) of Regulation (EC) No 1924/20061. EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). EFSA J. 2010, 8, 1817. [Google Scholar] [CrossRef]
- Jacobs, D.R.; Steffen, L.M. Nutrients, foods, and dietary patterns as exposures in research: A framework for food synergy. Am. J. Clin. Nutr. 2003, 78, 508S–513S. [Google Scholar] [CrossRef]
- Koh-Banerjee, P.; Franz, M.; Sampson, L.; Liu, S.; Jacobs, D.R.; Spiegelman, D.; Willett, W.; Rimm, E. Changes in whole-grain, bran, and cereal fiber consumption in relation to 8 − y weight gain among men. Am. J. Clin. Nutr. 2004, 80, 1237–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, D.R.; Meyer, K.A.; Kushi, L.H.; Folsom, A.R. Is whole grain intake associated with reduced total and cause-specific death rates in older women? The Iowa Women’s Health Study. Am. J. Public Health 1999, 89, 322–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Stampfer, M.J.; Hu, F.B.; Giovannucci, E.; Rimm, E.; Manson, J.E.; Hennekens, C.H.; Willett, W.C. Whole-grain consumption and risk of coronary heart disease: Results from the Nurses’ Health Study. Am. J. Clin. Nutr. 1999, 70, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, E.Q.; Chacko, S.A.; Chou, E.L.; Kugizaki, M.; Liu, S. Greater Whole-Grain Intake Is Associated with Lower Risk of Type 2 Diabetes, Cardiovascular Disease, and Weight Gain. J. Nutr. 2012, 142, 1304–1313. [Google Scholar] [CrossRef]
- De Munter, J.S.L.; Hu, F.B.; Spiegelman, D.; Franz, M.; van Dam, R.M. Whole Grain, Bran, and Germ Intake and Risk of Type 2 Diabetes: A Prospective Cohort Study and Systematic Review. PLoS Med. 2007, 4, e261. [Google Scholar] [CrossRef]
- Adams, J.F.; Engstrom, A.A. Dietary intake of whole grain vs. recommendations. Cereal Foods World 2000, 45, 75–78. [Google Scholar]
- Sheperd, R.; Dean, M.; Lampila, P.; Arvola, A.; Saba, A.; Vassallo, M.; Claupein, E.; Winkelmann, M.; Lähteenmäki, L. Communicating the benefits of wholegrain and functional grain products to European consumers. Trends Food Sci. Technol. 2012, 25, 63–69. [Google Scholar] [CrossRef]
- Javed, M.M.; Zahoor, S.; Shafaat, S.; Mehmooda, I.; Gul, A.; Rashee, H.; Bukhari, S.A.I.; Aftab, M.N.; Ikram-ul-Haq. Wheat bran as a brown gold: Nutritious value and its biotechnological applications. Afr. J. Microbiol. Res. 2012, 6, 724–733. [Google Scholar]
- Food and Agriculture Organization of United Nations (FAO). FAO Cereal Supply and Demand Brief. Available online: http://www.fao.org/worldfoodsituation/csdb/en/ (accessed on 27 May 2019).
- Katileviciute, A.; Plakys, G.; Budreviciute, A.; Onder, K.; Damiati, S.; Kodzius, R. A Sight to Wheat Bran: High Value-Added Products. Biomolecules 2019, 9, 887. [Google Scholar] [CrossRef] [Green Version]
- Heuzé, V.; Tran, G.; Baumont, R.; Noblet, J.; Renaudeau, D.; Lessire, M.; Lebas, F. Wheat bran. Feedipedia, a programme by INRAE, CIRAD, AFZ and FAO. 2005. Available online: https://www.feedipedia.org/node/726 (accessed on 14 October 2020).
- Prückler, M.; Siebenhandl-Ehn, S.; Apprich, S.; Höltinger, S.; Haas, C.; Schmid, E.; Kneifel, W. Wheat bran-based biorefinery 1: Composition of wheat bran and strategies of functionalization. LWT—Food Sci. Technol. 2014, 56, 211–221. [Google Scholar] [CrossRef]
- Cornell, H.; Hoveling, A.W. Wheat: Chemistry and Utilization; CRC Press Taylor & Francis Group: New York, NY, USA, 2020. [Google Scholar] [CrossRef]
- Betschart, A.A. Nutritional quality of wheat and wheat products. In Wheat, Chemistry and Technology, 3rd ed.; Pomeranz, Y., Ed.; American Association of Cereal Chemists: Saint Paul, MN, USA, 1988; pp. 91–129. [Google Scholar]
- Lawton, C.L.; Walton, J.; Hoyland, A.; Howarth, E.; Allan, P.; Chesters, D.; Dye, L. Short term (14 days) consumption of insoluble wheat bran fibre-containing breakfast cereals improves subjective digestive feelings, general wellbeing and bowel function in a dose dependent manner. Nutrients 2013, 5, 1436–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slavin, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandil, A.; Li, J.; Vasanthan, T.; Bressler, D.C. Phenolic acids in Some Cereal Grains and Their Inhibitory Effect on Starch Liquefaction and Saccharification. J. Agric. Food Chem. 2012, 60, 8444–8449. [Google Scholar] [CrossRef] [PubMed]
- Arranz, S.; Silvan, J.M.; Saura-Calixto, F. Nonextractable polyphenols, usually ignored, are the major part of dietary polyphenols: A study on the Spanish diet. Mol. Nutr. Food Res. 2010, 54, 1646–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gani, A.; Wani, S.M.; Masoodi, F.A.; Hameed, G. Whole-Grain Cereal Bioactive Compounds and Their Health Benefits: A Review. J. Food Process. Technol. 2012, 3, 146. [Google Scholar] [CrossRef] [Green Version]
- Holtekjølen, A.K.; Bævre, A.B.; Rødbotten, M.; Berg, H.; Knutsen, S.H. Antioxidant properties and sensory profiles of breads containing barley flour. Food Chem. 2008, 110, 414–421. [Google Scholar] [CrossRef]
- Kim, M.-J.; Hyun, J.-N.; Kim, J.-A.; Park, J.-C.; Kim, M.-Y.; Kim, J.-G.; Lee, S.-J.; Chun, S.-C.; Chung, I.-M. Relation between Phenolic Compounds, Anthocyanins Content and Antioxidant Activity in Colored Barley Germplasm. J. Agric. Food Chem. 2007, 55, 4802–4809. [Google Scholar] [CrossRef]
- Apea-Bah, F.; Beta, T. Bound Phenolic Constituents as Co-passengers of Dietary Fibre. In Cereal Grain-Based Functional Foods: Carbohydrate and Phytochemical Components; Beta, T., Camire, M.E., Eds.; The Royal Society of Chemistry: London, UK, 2019; pp. 278–304. [Google Scholar]
- Ndolo, V.U.; Beta, T.; Fulcher, R.G. Ferulic acid fluorescence intensity profiles and concentration measured by HPLC in pigmented and non-pigmented cereals. Food Res. Int. 2013, 52, 109–118. [Google Scholar] [CrossRef]
- Lempereur, I.; Rouau, J.; Abecassis, J. Genetic and Agronomic Variation in Arabinoxylan and Ferulic Acid Contents of Durum Wheat (Triticum durum L.) Grain and Its Milling Fractions. J. Cereal Sci. 1997, 25, 103–110. [Google Scholar] [CrossRef]
- Krygier, K.; Sosulski, F.; Hogge, L. Free, esterified, and insoluble-bound phenolic acids. 1. Extraction and purification procedure. J. Agric. Food Chem. 1982, 30, 330–334. [Google Scholar] [CrossRef]
- Fazary, A.E.; Ju, Y.-H. Feruloyl Esterases as Biotechnological Tools: Current and Future Perspectives. Acta Biochim. Biophys. Sin. 2007, 39, 811–828. [Google Scholar] [CrossRef] [PubMed]
- Bunzel, M.; Ralph, J.; Marita, J.M.; Hatfield, R.D.; Steinhar, H. Diferulates as structural components in soluble and insoluble cereal dietary fibre. J. Sci. Food Agric. 2011, 81, 653–660. [Google Scholar] [CrossRef]
- Andreasan, M.F.; Kroon, P.A.; Williamson, G.; Garcia-Conesa, M.T. Intestinal release and uptake of phenolic antioxidant diferulic acids. Free Radic. Biol. Med. 2001, 31, 304–314. [Google Scholar] [CrossRef]
- AOAC. Methods 990.03, 2003.05, 985.29 & 923.03. In Official Methods of Analysis of AOAC International, 18th ed.; AOAC: Gaithersburg, MD, USA, 2005. [Google Scholar]
- Goñi, I.; Díaz-Rubio, M.E.; Pérez-Jiménez, J.; Saura-Calixto, F. Towards an updated methodology for measurement of dietary fiber, including associated polyphenols, in food and beverages. Food Res. Int. 2009, 42, 840–846. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- AOCS. Official Method Ce 8-89. In Determination of Tocopherols and Tocotrienols 314 in Vegetable Oil and Fat by HPLC; AOCS: Champaign, IL, USA, 1992. [Google Scholar]
- Slinkard, K.; Singleton, V.L. Total Phenol Analyses: Automation and Comparison with Manual Methods. Am. J. Enol. Vitic. 1977, 28, 49–55. [Google Scholar]
- Martin-Diana, A.B.; Izquierdo, N.; Albertos, I.; Sanchez, M.S.; Herrero, A.; Sanz, M.A.; Rico, D. Valorization of Carob’s Germ and Seed Peel as Natural Antioxidant Ingredients in Gluten-Free Crackers. J. Food Process. Preserv. 2017, 41, e12770. [Google Scholar] [CrossRef]
- García-Villalba, R.; Espín, J.C.; Aaby, K.; Alasalvar, C.; Heinonen, M.; Jacobs, G.; Voorspoels, S.; Koivumäki, T.; Kroon, P.A.; Pelvan, E.; et al. Validated Method for the Characterization and Quantification of Extractable and Nonextractable Ellagitannins after Acid Hydrolysis in Pomegranate Fruits, Juices, and Extracts. J. Agric. Food Chem. 2015, 63, 6555–6566. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and Validation of an Improved Oxygen Radical Absorbance Capacity Assay Using Fluorescein as the Fluorescent Probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Pereira, J.A.; Oliveira, I.; Sousa, A.; Ferreira, I.C.F.R.; Bento, A.; Estevinho, L. Bioactive properties and chemical composition of six walnut (Juglans regia L.) cultivars. Food Chem. Toxicol. 2008, 46, 2103–2111. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Tanumihardjo, S.A. An Integrated Approach to Evaluate Food Antioxidant Capacity. J. Food Sci. 2007, 72, R159–R165. [Google Scholar] [CrossRef]
- Gularte, M.A.; Rosell, C.M. Physicochemical properties and enzymatic hydrolysis of different starches in the presence of hydrocolloids. Carbohydr. Polym. 2011, 85, 237–244. [Google Scholar] [CrossRef] [Green Version]
- Granfeldt, Y.; Björck, I.; Drews, A.; Tovar, J. An in vitro procedure based on chewing to predict metabolic response to starch in cereal and legume products. Eur. J. Clin. Nutr. 1992, 46, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Šterna, V.; Zute, S.; Brunava, L. Oat Grain Composition and its Nutrition Benefice. Agric. Agric. Sci. Procedia 2016, 8, 252–256. [Google Scholar] [CrossRef] [Green Version]
- Šrámková, Z.; Gregová, E.; Šturdík, E. Chemical composition and nutritional quality of wheat grain. Acta Chim. Slovaca 2009, 2, 115–138. [Google Scholar]
- Jung, H.G.; Allen, M.S. Characteristics of plant cell walls affecting intake and digestibility of forages by ruminants. J. Anim. Sci. 1995, 73, 2774–2790. [Google Scholar] [CrossRef]
- Sontag-Strohm, T.; Lehtinen, P.; Kaukovirta-Norja, A. Oat products and their current status in the celiac diet. In Gluten-Free Cereal Products and Beverages; Arendt, E.K., Dal Bello, F., Eds.; Academic Press: San Diego, CA, USA, 2008; pp. 192–202. [Google Scholar] [CrossRef]
- Dhinda, F.; Jyothi, L.A.; Prakash, J.; Dasappa, I. Effect of Ingredients on Rheological, Nutritional and Quality Characteristics of High Protein, High Fibre and Low Carbohydrate Bread. Food Bioprocess Technol. 2012, 5, 2998–3006. [Google Scholar] [CrossRef]
- Halima, N.B.; Saad, R.B.; Khemakhem, B.; Fendri, I.; Abdelkafi, S. Oat (Avena sativa L.): Oil and Nutriment Compounds Valorization for Potential Use in Industrial Applications. J. Oleo Sci. 2015, 64, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Narducci, V.; Finotti, E.; Galli, V.; Carcea, M. Lipids and Fatty Acids in Italian Durum Wheat (Triticum durum Desf.) Cultivars. Foods 2019, 8, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burton, G.W.; Traber, M.G. Vitamin E: Antioxidant activity, biokinetics, and bioavailability. Annu. Rev. Nutr. 1999, 10, 357–382. [Google Scholar] [CrossRef] [PubMed]
- Ciudad-Mulero, M.; Barros, L.; Fernandes, A.; Ferreira, I.C.F.R.; Callejo, M.; Matallana-González, M.; Fernández-Ruiz, V.; Morales, P.; Carrillo, J.M. Potential Health Claims of Durum and Bread Wheat Flours as Functional Ingredients. Nutrients 2020, 12, 504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidalgo, A.; Brandolini, A. Protein, ash, lutein and tocols distribution in einkorn (Triticum monococcum L. subsp. monococcum) seed fractions. Food Chem. 2008, 107, 444–448. [Google Scholar] [CrossRef]
- Pasha, I.; Ahmad, F.; Siddique, Z.; Iqbal, F. Probing the effect of physical modifications on cereal bran chemistry and antioxidant potential. J. Food Meas. Charact. 2020, 14, 1909–1918. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F. Importance of Insoluble-Bound Phenolics to Antioxidant Properties of Wheat. J. Agric. Food Chem. 2006, 54, 1256–1264. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Segura-Carretero, A.; Di Silvestro, R.; Marotti, I.; Arráez-Román, D.; Benedettelli, S.; Ghiselli, L.; Fernández-Gutiérrez, A. Profiles of phenolic compounds in modern and old common wheat varieties determined by liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7670–7681. [Google Scholar] [CrossRef]
- Peterson, D.M. Oat Antioxidants. J. Cereal Sci. 2001, 33, 115–129. [Google Scholar] [CrossRef]
- Călinoiu, L.F.; Vodnar, D.C. Thermal Processing for the Release of Phenolic Compounds from Wheat and Oat Bran. Biomolecules 2020, 10, 21. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Piironen, V.; Lampi, A.-M.; Nyström, L.; Li, L.; Rakszegi, M.; Fraś, A.; Boros, D.; Gebruers, K.; Courtin, C.M.; et al. Phytochemical and Fiber Components in Oat Varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9777–9784. [Google Scholar] [CrossRef]
- Verma, B.; Hucl, P.; Chibbar, R.N. Phenolic Content and Antioxidant Properties of Bran in 51 Wheat Cultivars. Cereal Chem. 2008, 85, 544–549. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E. Ferulic Acid: An Antioxidant Found Naturally in Plant Cell Walls and Feruloyl Esterases Involved in its Release and Their Applications. Crit. Rev. Biotechnol. 2004, 24, 59–83. [Google Scholar] [CrossRef] [PubMed]
- Price, R.K.; Wallace, J.M.W.; Hamill, L.L.; Keaveney, E.M.; Strain, J.J.; Parker, M.J.; Welch, R.W. Evaluation of the effect of wheat aleurone-rich foods on markers of antioxidant status, inflammation and endothelial function in apparently healthy men and women. Br. J. Nutr. 2012, 108, 1644–1651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, R.K.; Welch, R.W.; Lee-Manion, A.M.; Bradbury, I.; Strain, J.J. Total Phenolics and Antioxidant Potential in Plasma and Urine of Humans After Consumption of Wheat Bran. Cereal Chem. 2008, 85, 152–157. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Deemer, E.K. Development and validation of oxygen radical absorbance capacity assay for lipophilic antioxidants using randomly methylated cyclodextrin as the solubility enhancer. J. Agric. Food Chem. 2002, 50, 1815–1821. [Google Scholar] [CrossRef]
- Halliwell, B. Antioxidants: The basics—What they are and how to evaluate them. Adv. Pharmacol. 1997, 38, 3–20. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Soong, Y.Y.; Tan, S.P.; Leong, L.P.; Henry, J.K. Total antioxidant capacity and starch digestibility of muffins baked with rice, wheat, oat, corn and barley flour. Food Chem. 2014, 164, 462–469. [Google Scholar] [CrossRef]
- Beta, T.; Nam, S.; Dexter, J.E.; Sapirstein, H.D. Phenolic Content and Antioxidant Activity of Pearled Wheat and Roller-Milled Fractions. Cereal Chem. 2005, 82, 390–393. [Google Scholar] [CrossRef]
- Yu, L.; Nanguet, A.-L.; Beta, T. Comparison of Antioxidant Properties of Refined and Whole Wheat Flour and Bread. Antioxidants 2013, 2, 370–383. [Google Scholar] [CrossRef] [Green Version]
- Ragaee, S.; Seetharaman, K.; Abdel-Aal, E.-S.M. The impact of milling and thermal processing on phenolic compounds in cereal grains. Crit. Rev. Food Sci. Nutr. 2014, 54, 837–849. [Google Scholar] [CrossRef]
- Yang, Z.; Han, Y.; Gu, Z.; Fan, G.; Chen, Z. Thermal degradation kinetics of aqueous anthocyanins and visual color of purple corn (Zea mays L.) cob. Innov. Food Sci. Emer. Technol. 2008, 9, 341–347. [Google Scholar] [CrossRef]
- Sandhu, K.S.; Singh, N.; Malhi, N.S. Some properties of corn grains and their flours. I: Physicochemical, functional and chapati-making properties of flours. Food Chem. 2007, 101, 938–946. [Google Scholar] [CrossRef]
- Wolever, T.M.S. The Glycaemic Index: A Physiological Classification of Dietary Carbohydrate; CABI: Wallingford, UK, 2006; pp. 1–227. [Google Scholar] [CrossRef]
- Brennan, C.; Tudorica, C. Evaluation of potential mechanisms by which dietary fiber additions reduce the predicted glycemic index of fresh pasta. Int. J. Food Sci. Technol. 2008, 43, 2151–2162. [Google Scholar] [CrossRef]
- Onipe, O.O.; Beswa, D.; Jideani, A.I.O. In vitro starch digestibility and glycaemic index of fried dough and batter enriched with wheat and oat bran. Foods 2020, 9, 1374. [Google Scholar] [CrossRef]
- Xie, X.; Cui, S.; Li, W.; Tsao, R. Isolation and characterization of wheat bran starch. Food Res. Int. 2008, 41, 882–887. [Google Scholar] [CrossRef]
- Murugadass, G.; Dipnaik, K. Preliminary study of ratio of amylose and amylopectin as indicators of glycemic index and in vitro enzymatic hydrolysis of rice and wheat starches. Int. J. Res. Med. Sci. 2018, 6, 3095. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WG | WB | OG | OB | ||
|---|---|---|---|---|---|
| Ash | 1.78 ± 0.03 b | 7.31 ± 0.04 d | 1.69 ± 0.02 a | 3.03 ± 0.01 c | |
| Fat | 2.18 ± 0.01 a | 2.73 ± 0.46 b | 7.27 ± 0.26 c | 9.20 ± 0.78 d | |
| Moisture | 9.45 ± 0.32 b | 13.64 ± 0.20 d | 8.92 ± 0.02 a | 10.02 ± 0.08 c | |
| Protein | 10.66 ± 0.04 a | 13.19 ± 0.27 c | 11.50 ± 0.27 b | 17.63 ± 0.00 d | |
| Carbohydrates | 75.94 ± 0.24 c | 63.14 ± 0.49 b | 70.63 ± 0.58 c | 60.12 ± 0.69 a | |
| Soluble Fiber | NSPs | 1.35 ± 0.003 a | 1.55 ± 0.29 ab | 3.06 ± 0.34 b | 4.50 ± 0.33 c |
| Insoluble Fiber | NSPs | 11.56 ± 1.10 b | 43.18 ± 3.08 c | 7.17 ± 0.50 a | 7.71 ± 0.62 a |
| Klason Lignin | 2.01 ± 0.44 b | 9.16 ± 1.27 c | 1.49 ± 0.44 a | 11.97 ± 1.46 d | |
| Total Fiber | 15.10 ± 1.18 b | 53.89 ± 3.34 d | 11.72 ± 0.74 a | 24.18 ± 1.62 c | |
| WG | WB | OG | OB | |
|---|---|---|---|---|
| SOLUBLE FRACTION | ||||
| Ferulic acid | 25.40 ± 1.10 a | 51.93 ± 2.69 b | N.D. | N.D. |
| Caffeic Acid-4-Glucoside | 17.69 ± 3.80 a | 67.33 ± 1.71 d | 22.39 ± 0.11 b | 30.45 ± 1.19 c |
| Isovitexin 8-C-β-glucoside (Vicenin-2) | 0.00 ± 0.00 a | 36.92 ± 0.68 d | 6.99 ± 0.23 b | 7.81 ± 0.10 c |
| Apigenin-6/8-C-pentoside-8/6-C-hexoside I | 19.96 ± 7.00 a | 15.56 ± 0.85 a | 11.54 ± 8.36 a | 15.55 ± 6.53 a |
| Apigenin-6/8-C-pentoside-8/6-C-hexoside II | 12.66 ± 6.06 c | 7.24 ± 0.00 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Apigenin-6/8-C-pentoside-8/6-C-hexoside III | 12.09 ± 1.09 c | 0.00 ± 0.00 a | 5.29 ± 0.74 b | 4.26 ± 1.51 b |
| Apigenin-8-C-glucoside (Vitexin) | 115.97 ± 0.06 c | 551.05 ± 28.50 d | 24.74 ± 1.90 a | 49.63 ± 1.59 b |
| 1-Acetoxy pinoresinol | 0.00 ± 0.00 a | 40.42 ± 1.88 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Daidzein | 0.00 ± 0.00 a | 27.39 ± 1.54 a | 29.17 ± 0.11 a | 28.72 ± 1.33 a |
| Apigenin-6-C-arabinoside-8-C-hexoside I | 24.95 ± 2.62 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Apigenin-6-C-arabinoside-8-C-hexoside II | 39.52 ± 0.82 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Avenanthramide K | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 6.15 ± 0.19 b | 8.16 ± 0.59 c |
| Glycosilated/Acetylated-3′,4′,5′-trihydroxy-3,7-dimethylflavone | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 11.47 ± 0.62 b | 8.16 ± 0.26 a |
| Apigenin-6-C-galactosyl-8-C-glucosyl-O-glucuropyranoside | 15.63 ± 0.65 b | 56.11 ± 5.29 c | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Avenanthramide 2c | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 17.28 ± 0.24 c | 13.67 ± 0.01 b |
| Avenanthramide 2p | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 32.28 ± 0.32 c | 17.23 ± 0.86 b |
| Avenanthramide 2f | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 27.29 ± 0.00 b | 24.59 ± 1.60 a |
| Total | 283.88 ± 0.50 c | 853.94 ± 39.08 d | 194.60 ± 10.53 a | 208.23 ± 15.56 b |
| INSOLUBLE FRACTION | ||||
| Medioresinol | 233.48 ± 0.69 c | 891.66 ± 54.60 d | 126.23 ± 4.22 b | 76.22 ± 7.18 a |
| Ferulic acid | 3776.53 ± 4.74 b | 16,936.18 ± 1055.39 c | 1165.71 ± 5.63 a | 1769.94 ± 181.73 b |
| Dihydroferulic Acid I | 47.35 ± 0.36 c | 0.00 ± 0.00 a | 24.64 ± 5.44 a | 36.86 ± 1.98 b |
| Dihydroferulic Acid II | 188.41 ± 1.48 b | 333.62 ± 36.69 c | 86.65 ± 42.89 a | 88.08 ± 3.63 a |
| Dihydroferulic Acid III | 444.07 ± 0.07 c | 1215.03 ± 162.10 d | 190.58 ± 5.15 b | 182.94 ± 6.36 a |
| Total | 4689.84 ± 19.33 c | 19,376.57 ± 911.20 d | 1593.81 ± 52.07 a | 2154.04 ± 200.88 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Diana, A.B.; García-Casas, M.J.; Martínez-Villaluenga, C.; Frías, J.; Peñas, E.; Rico, D. Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients. Foods 2021, 10, 115. https://doi.org/10.3390/foods10010115
Martín-Diana AB, García-Casas MJ, Martínez-Villaluenga C, Frías J, Peñas E, Rico D. Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients. Foods. 2021; 10(1):115. https://doi.org/10.3390/foods10010115
Chicago/Turabian StyleMartín-Diana, Ana Belén, María Jesús García-Casas, Cristina Martínez-Villaluenga, Juana Frías, Elena Peñas, and Daniel Rico. 2021. "Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients" Foods 10, no. 1: 115. https://doi.org/10.3390/foods10010115
APA StyleMartín-Diana, A. B., García-Casas, M. J., Martínez-Villaluenga, C., Frías, J., Peñas, E., & Rico, D. (2021). Wheat and Oat Brans as Sources of Polyphenol Compounds for Development of Antioxidant Nutraceutical Ingredients. Foods, 10(1), 115. https://doi.org/10.3390/foods10010115