
Antimicrobial Activity and Action Mechanism of Thymoquinone against Bacillus cereus and Its Spores

Abstract
:1. Introduction
2. Materials and Methods
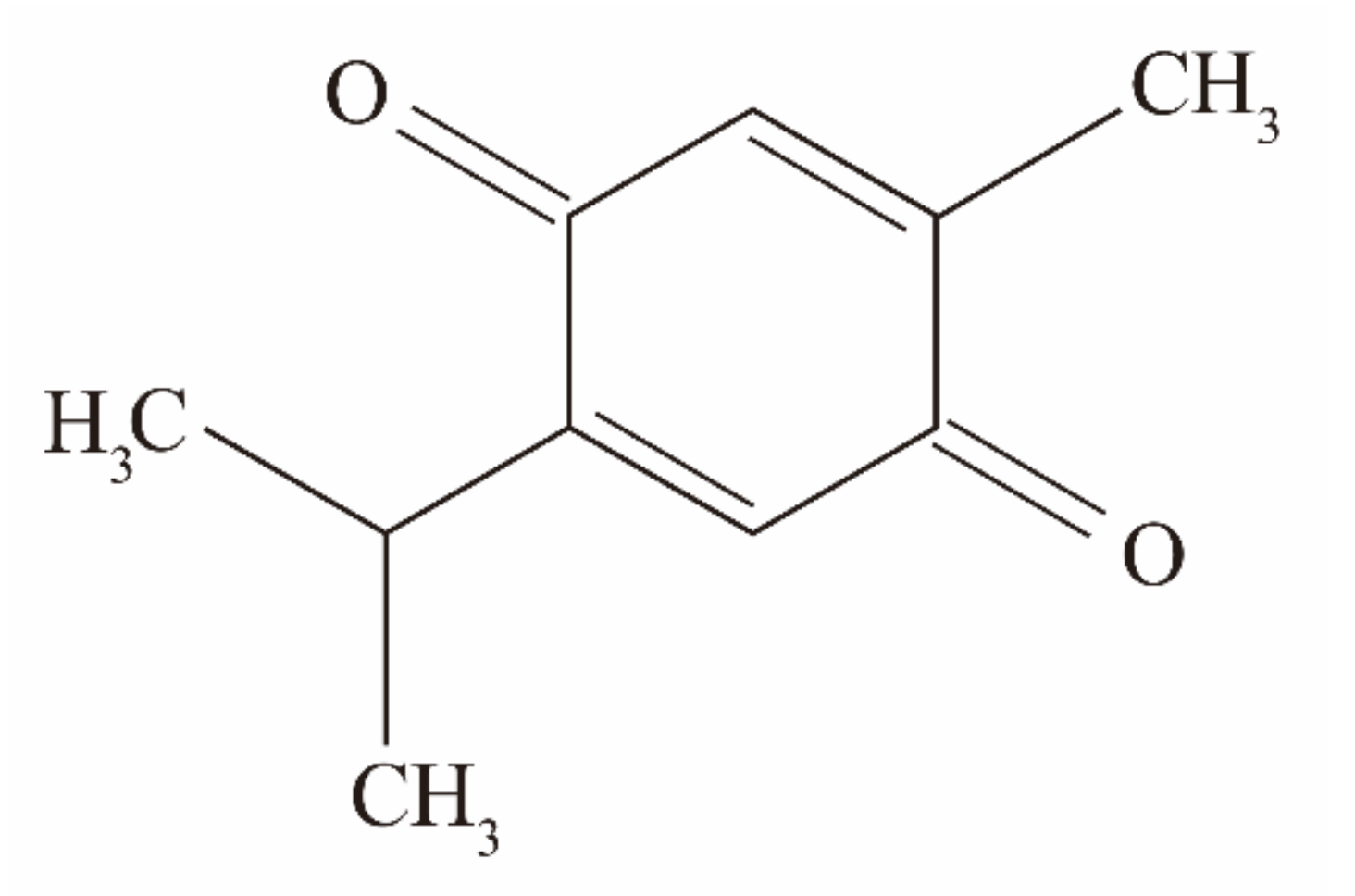
2.1. Reagents
2.2. Bacterial Strain, Growth Conditions, and Spore Preparation
2.3. Determinations of the MIC and MBC of TQ against B. cereus
2.4. Mechanism of Action against B. cereus
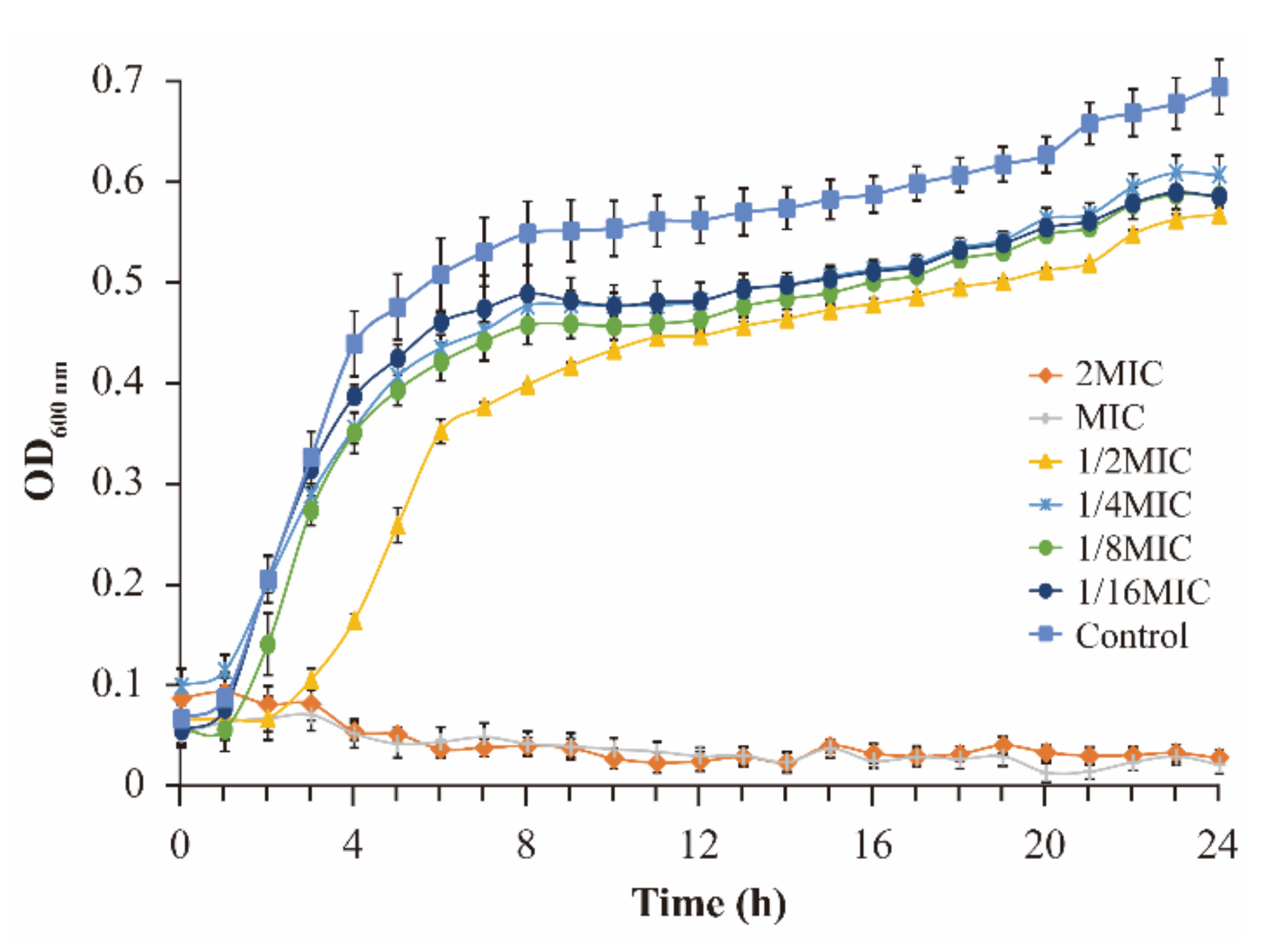
2.4.1. Growth Curve
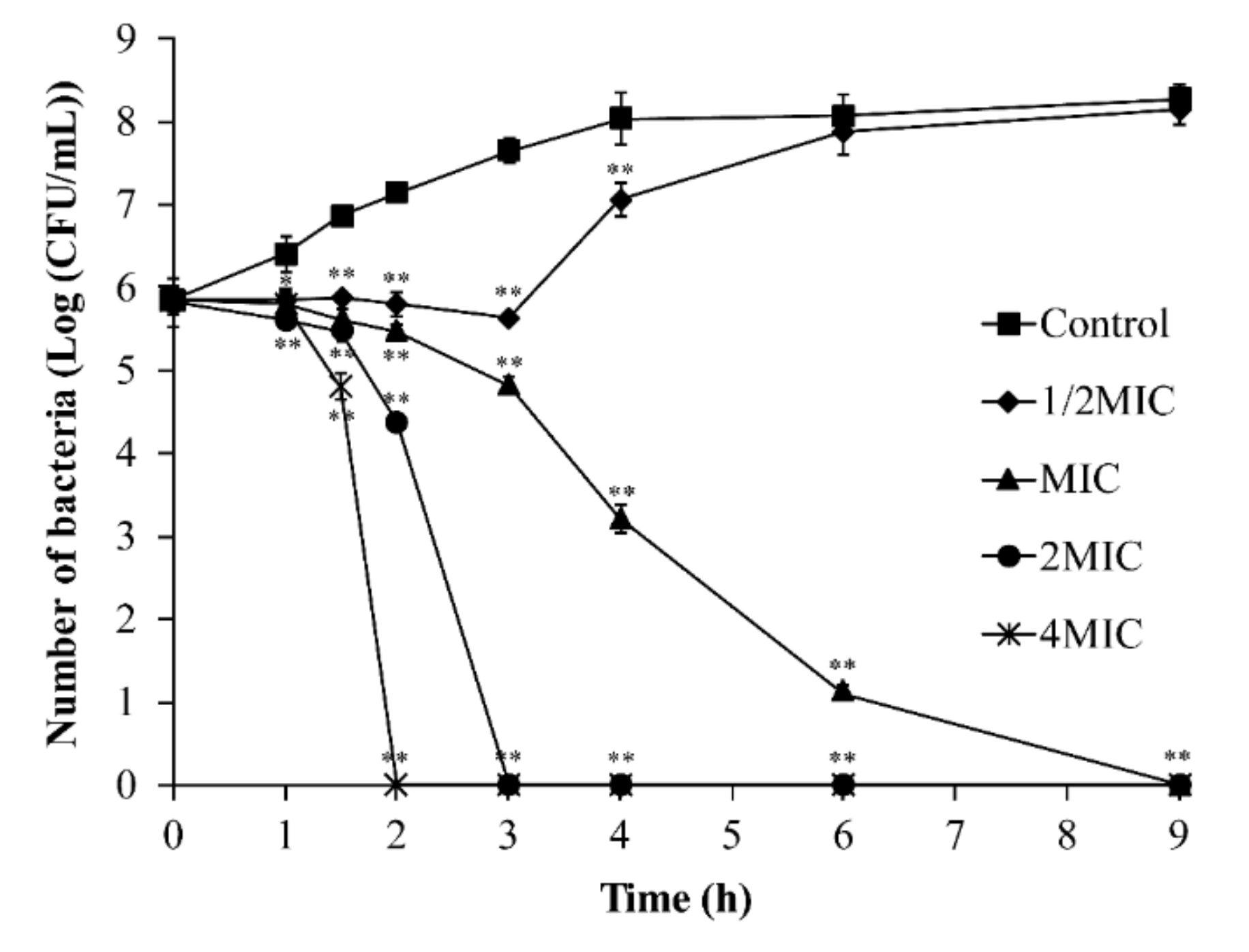
2.4.2. Inactivation of B. cereus by TQ in LB Broth
2.4.3. Membrane Potential
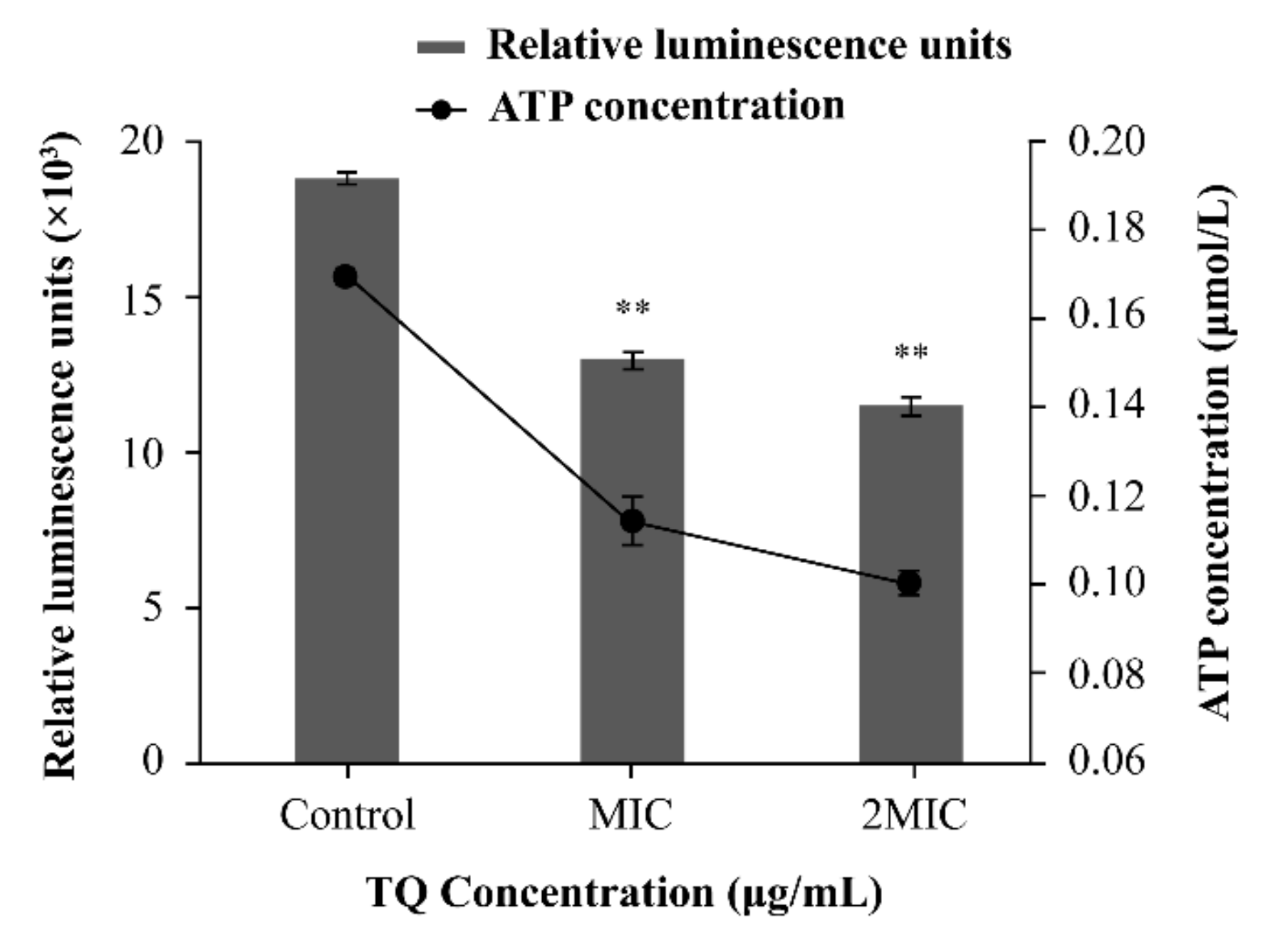
2.4.4. Determination of Intracellular ATP Concentrations
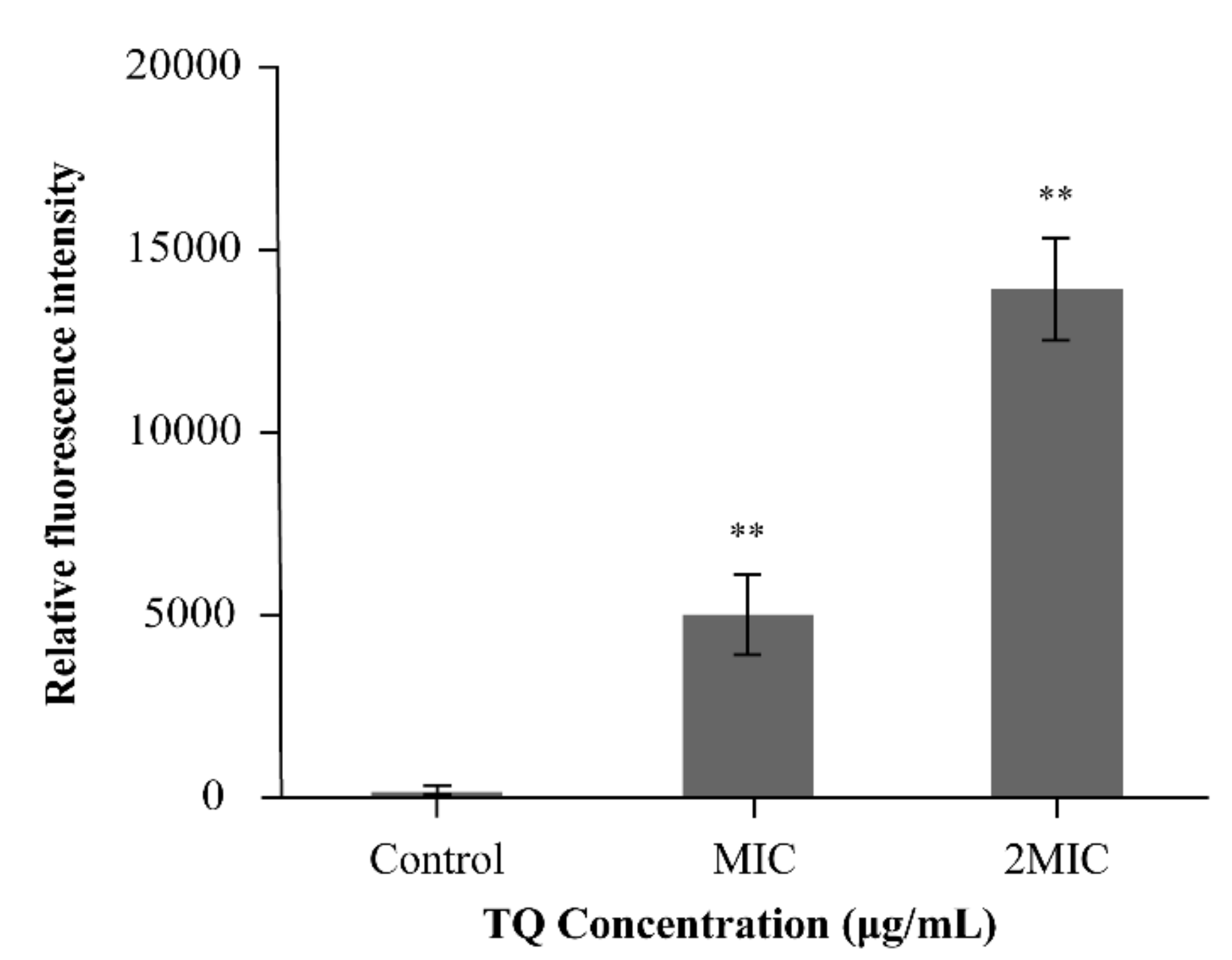
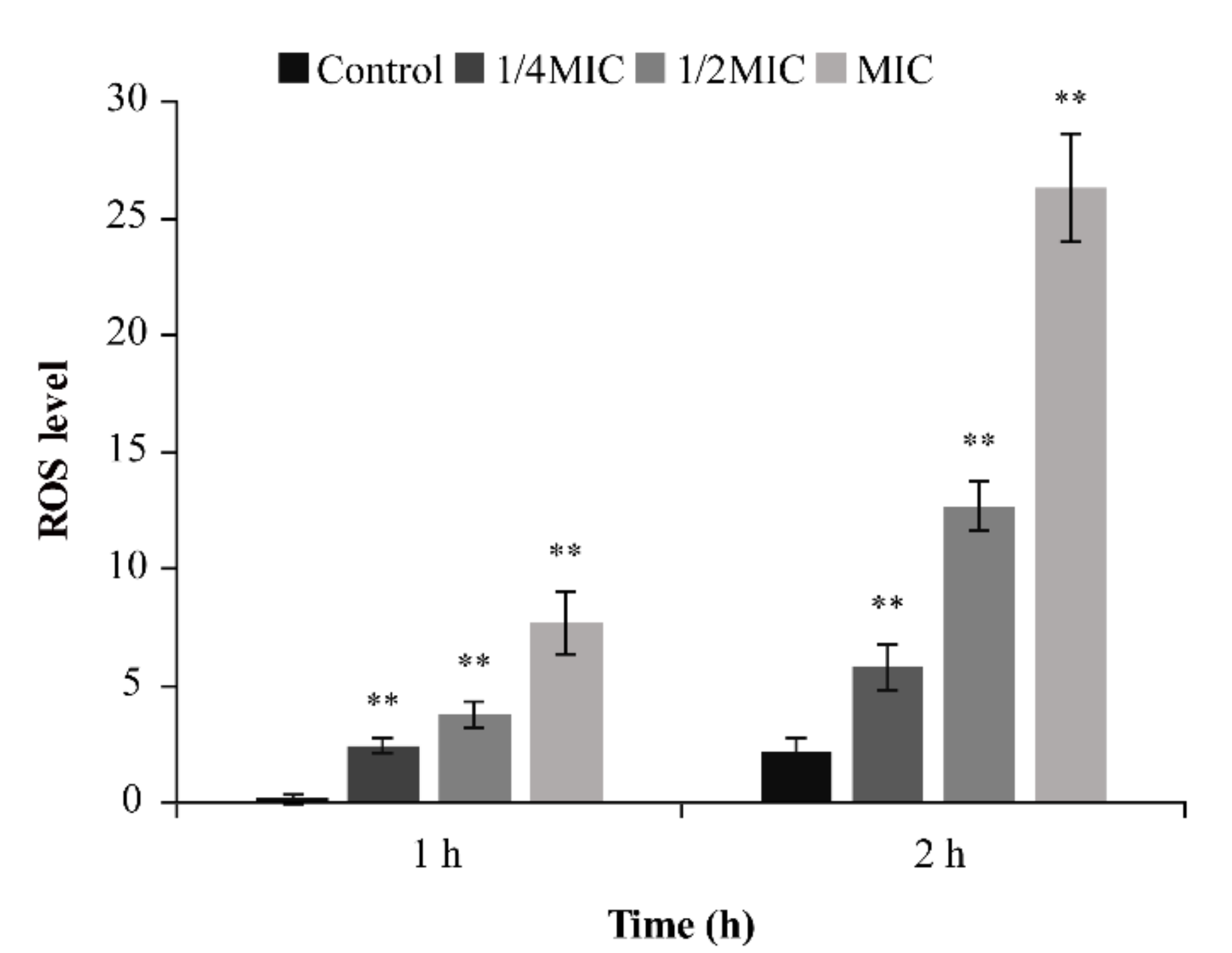
2.4.5. ROS Determination
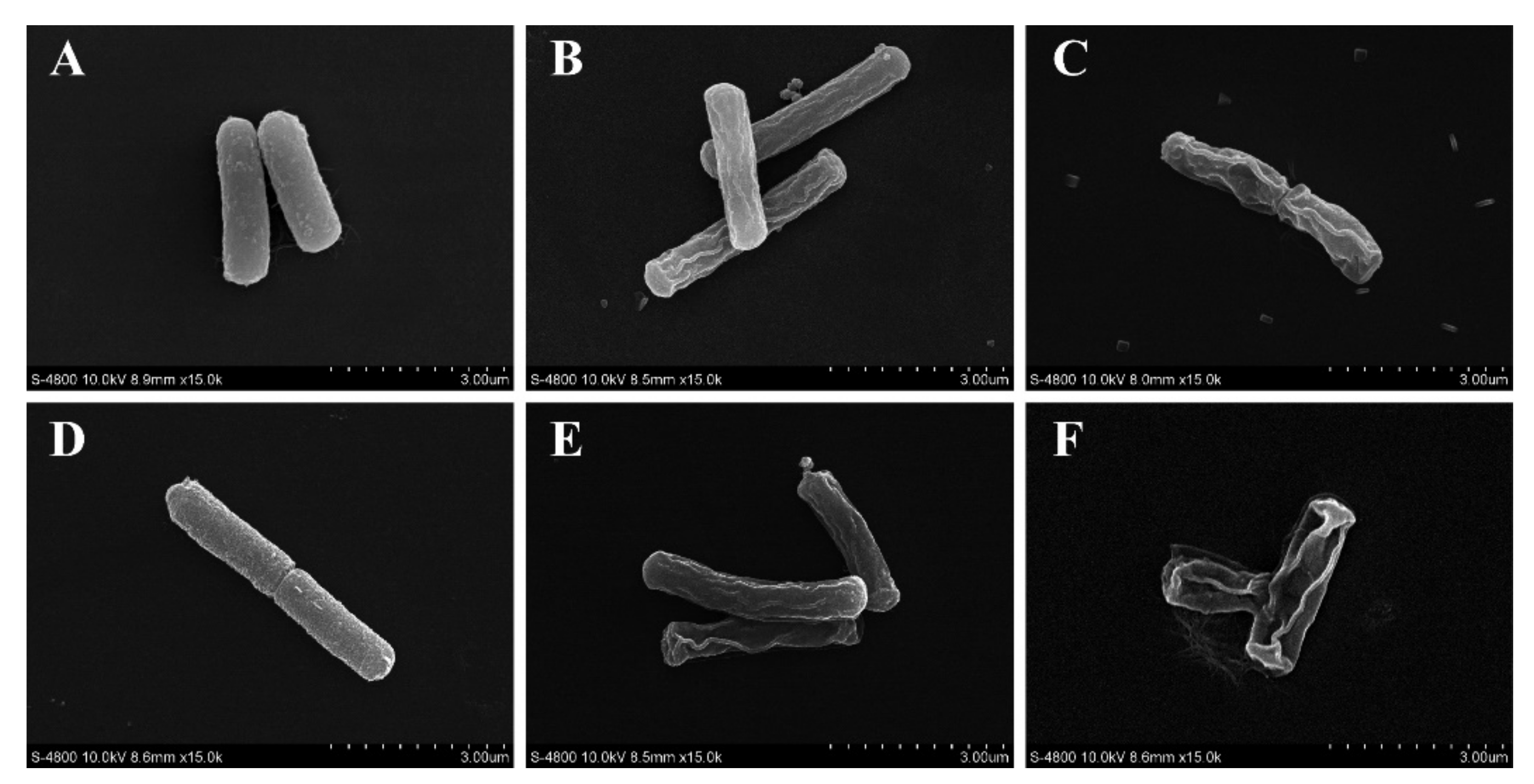
2.4.6. Bacterial Morphology
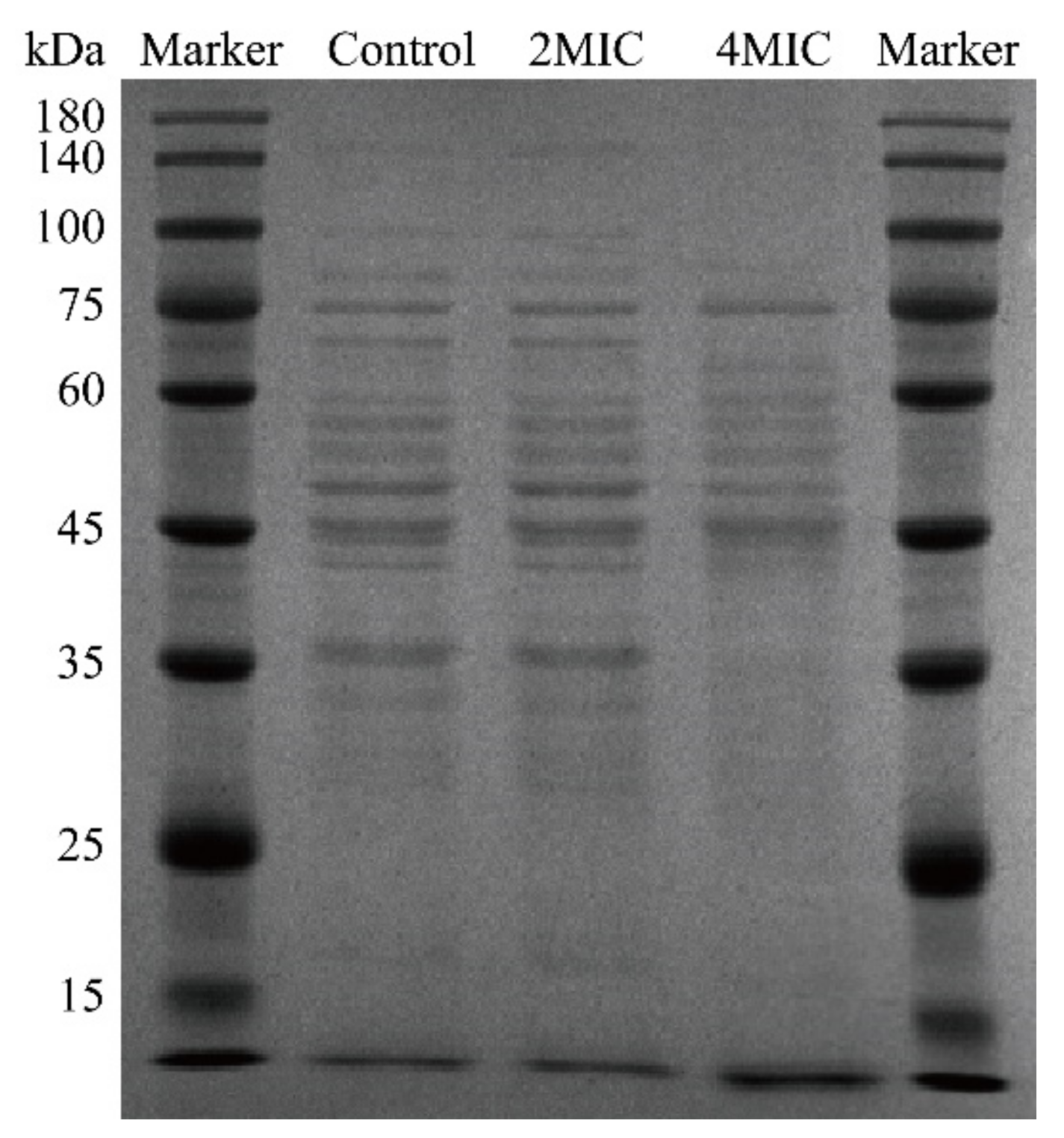
2.4.7. Sodium Dodecyl Sulfate–Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.4.8. Reverse Transcription–Quantitative PCR (RT-qPCR)
2.5. Determination of the Antibacterial Activity of TQ in Reconstituted Infant Formula (RIF)
2.6. Inhibitory Effect of TQ on B. cereus Spores
2.6.1. Effect of TQ on Spore Germination Rates
2.6.2. Confocal Laser-Scanning Microscopy Observations
2.7. Statistical Analysis
3. Results
3.1. MICs and MBCs
3.2. Growth Curve
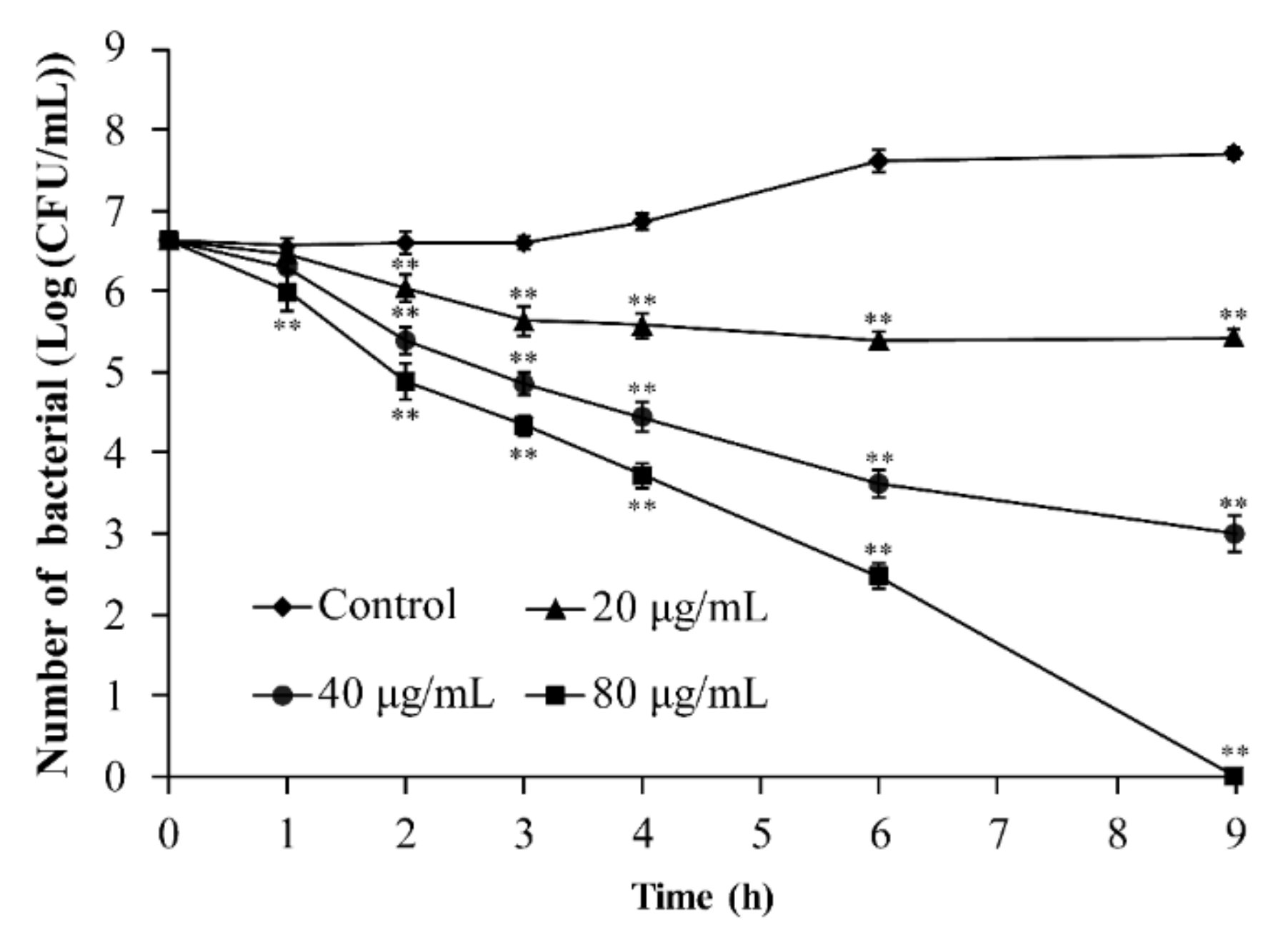
3.3. Inactivation of B. cereus by TQ in LB Broth
3.4. Antibacterial Mechanism of TQ against B. cereus
3.4.1. Membrane Potential
3.4.2. Changes in Intracellular ATP Concentrations
3.4.3. ROS Determination
3.4.4. Morphology of B. cereus
3.4.5. SDS-PAGE
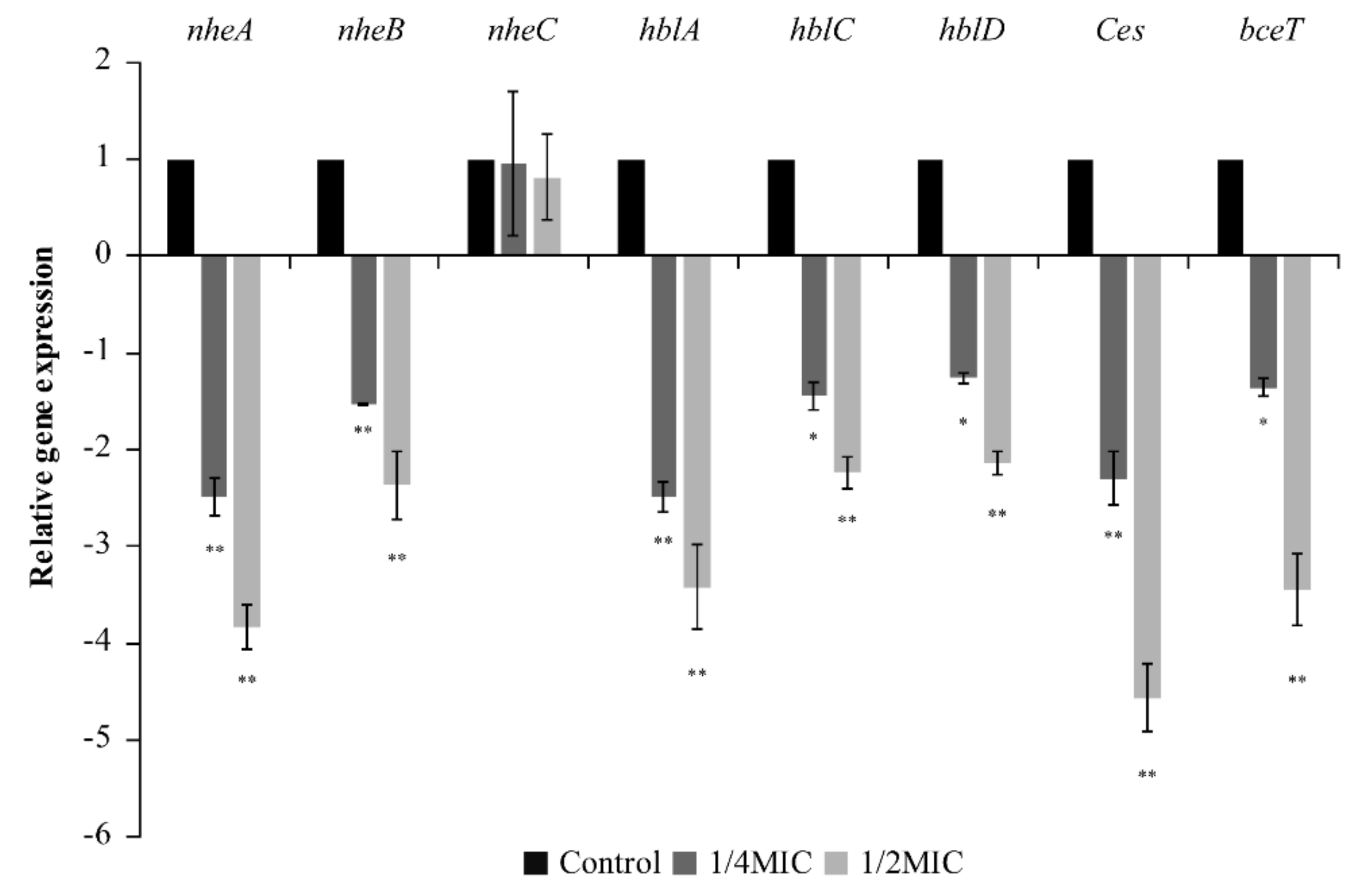
3.5. RT-qPCR Analysis
3.6. Determination of the Antibacterial Activity of TQ in RIF
3.7. Inhibitory Effect of TQ on B. cereus Spores
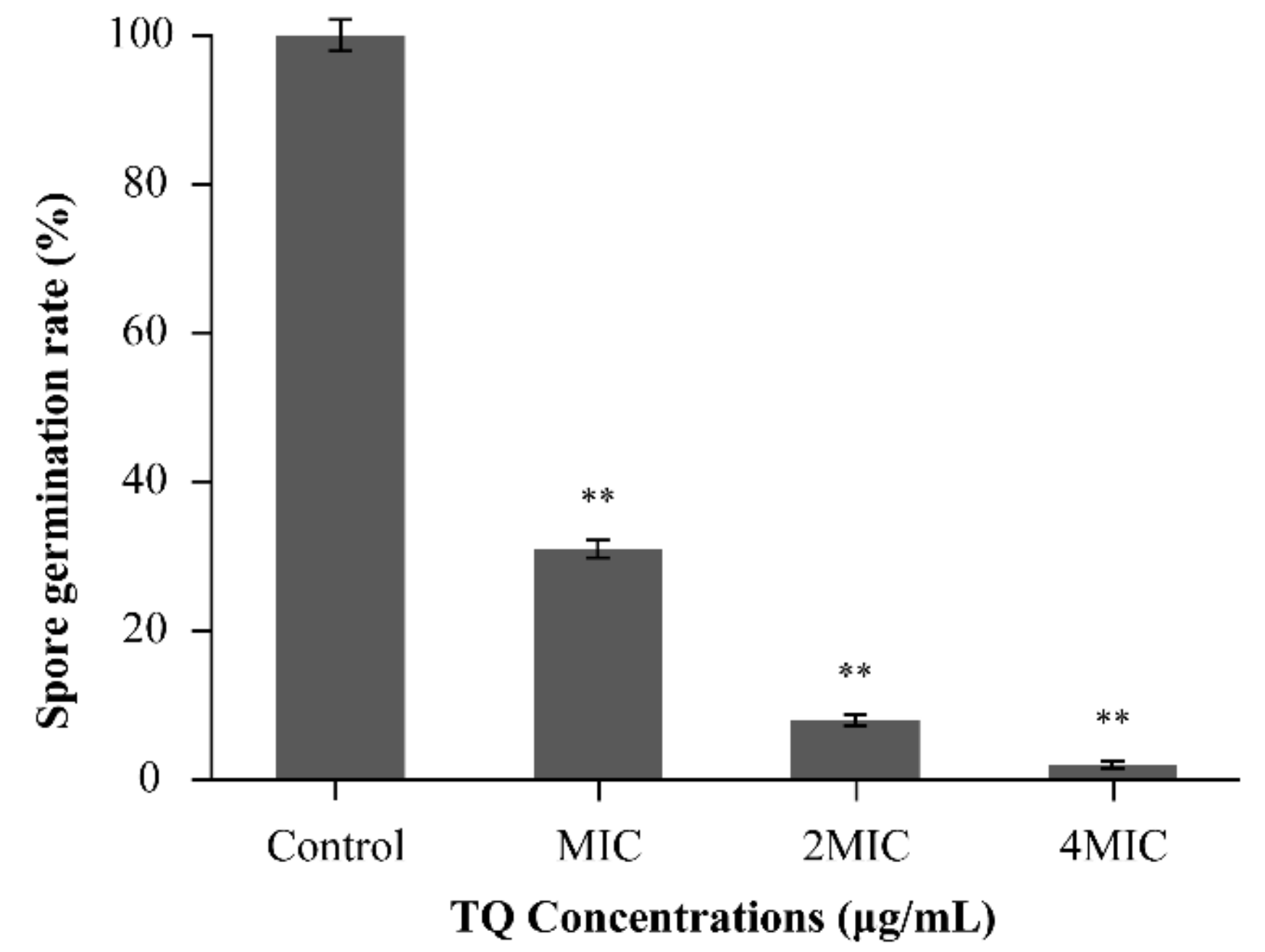
3.7.1. Inhibition of Spore Germination by TQ
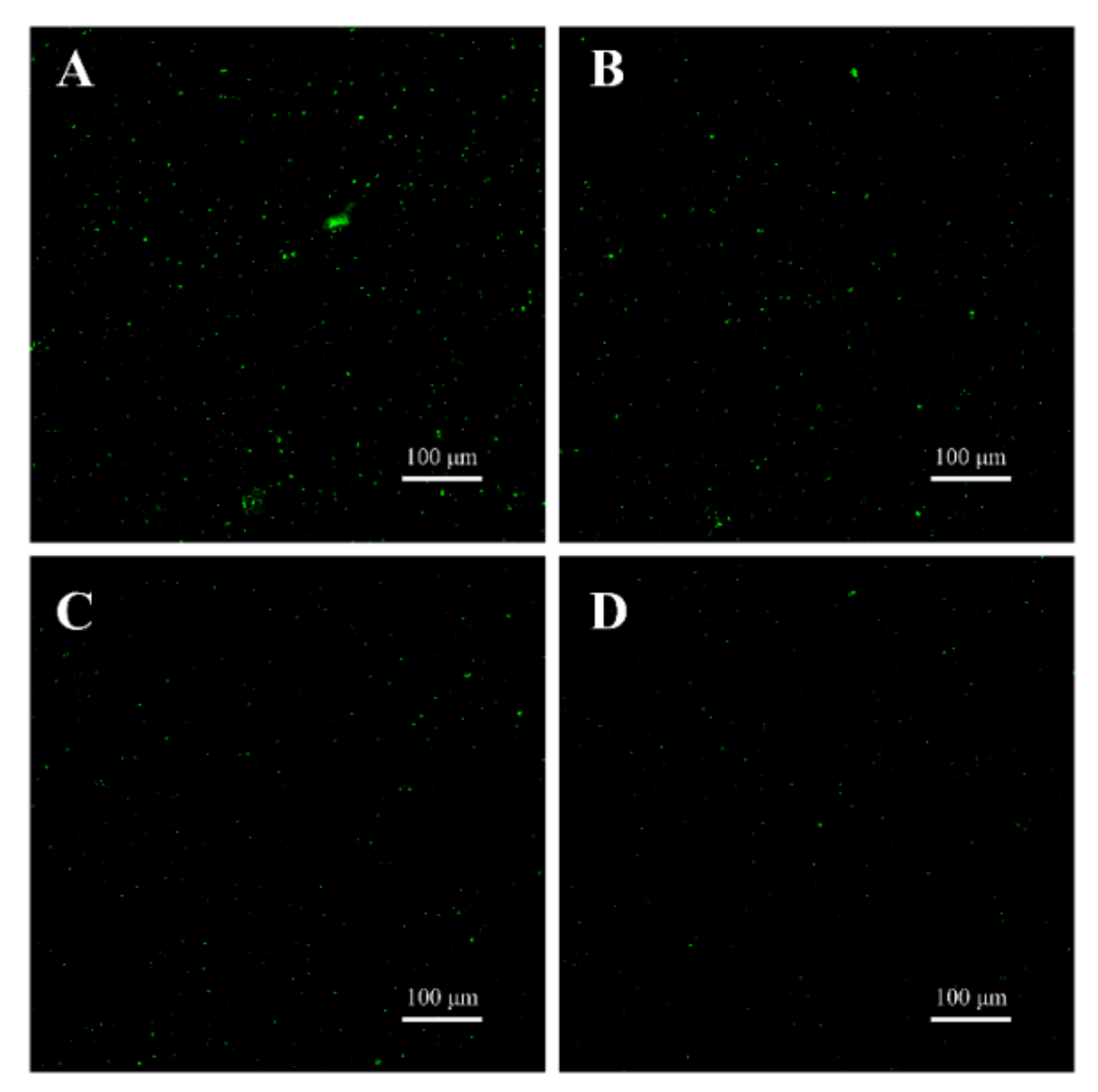
3.7.2. CLSM Observations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Osimani, A.; Aquilanti, L.; Clementi, F. Bacillus cereus foodborne outbreaks in mass catering. Int. J. Hosp. Manag. 2018, 72, 145–153. [Google Scholar] [CrossRef]
- Kim, B.; Bang, J.; Kim, H.; Kim, Y.; Kim, B.S.; Beuchat, L.R.; Ryu, J.H. Bacillus cereus and Bacillus thuringiensis spores in Korean rice: Prevalence and toxin production as affected by production area and degree of milling. Food Microbiol. 2014, 42, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Cetin-Karaca, H.; Newman, M.C. Antimicrobial efficacy of phytochemicals against Bacillus cereus in reconstituted infant rice cereal. Food Microbiol. 2018, 69, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Choma, C.; Guinebretiere, M.H.; Carlin, F.; Schmitt, P.; Nguyen-The, C. Prevalence, characterization and growth of Bacillus cereus in commercial cooked chilled foods containing vegetables. J. Appl. Microbiol. 2000, 88, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Soria, M.C.; Audisio, M.C. Inhibition of Bacillus cereus Strains by Antimicrobial Metabolites from Lactobacillus johnsonii CRL1647 and Enterococcus faecium SM21. Probiotics Antimicrob. Proteins 2014, 6, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, Virulence Genes, Antimicrobial Susceptibility, and Genetic Diversity of Bacillus cereus Isolated From Pasteurized Milk in China. Front. Microbiol. 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanovic, J.; Ornelis, V.F.M.; Madder, A.; Rajkovic, A. Bacillus cereus food intoxication and toxicoinfection. Compr. Rev. Food. Sci. Food Saf. 2021, 20, 3719–3761. [Google Scholar] [CrossRef]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore: Assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2013, 11, 33–44. [Google Scholar] [CrossRef] [PubMed]
- Webb, M.D.; Barker, G.C.; Goodburn, K.E.; Peck, M.W. Risk presented to minimally processed chilled foods by psychrotrophic Bacillus cereus. Trends Food Sci. Technol. 2019, 93, 94–105. [Google Scholar] [CrossRef]
- Paredes-Sabja, D.; Setlow, P.; Sarker, M.R. Germination of spores of Bacillales and Clostridiales species: Mechanisms and proteins involved. Trends Microbiol. 2011, 19, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.I.; Chung, M.S. Antimicrobial effect of a combination of herb extract and organic acid against Bacillus subtilis spores. Food Sci. Biotechnol. 2017, 26, 1423–1428. [Google Scholar] [CrossRef] [PubMed]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Husain, A.; Mujeeb, M.; Khan, S.A.; Najmi, A.K.; Siddique, N.A.; Damanhouri, Z.A.; Anwar, F. A review on therapeutic potential of Nigella sativa: A miracle herb. Asian Pac. Trop. Biomed. 2013, 3, 337–352. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Bai, T.; Sun, P.; Lian, L.-H.; Yao, Y.-L.; Zheng, H.-X.; Li, X.; Li, J.-B.; Wu, Y.-L.; Nan, J.-X. Thymoquinone, a bioactive component of Nigella sativa Linn seeds or traditional spice, attenuates acute hepatic failure and blocks apoptosis via the MAPK signaling pathway in mice. RSC Adv. 2015, 5, 7285–7290. [Google Scholar] [CrossRef]
- Darakhshan, S.; Bidmeshki Pour, A.; Hosseinzadeh Colagar, A.; Sisakhtnezhad, S. Thymoquinone and its therapeutic potentials. Pharmacol. Res. 2015, 95–96, 138–158. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Yan, C.; Sui, Y.; Sun, Y.; Guo, D.; Chen, Y.; Jin, T.; Peng, X.; Ma, L.; Xia, X. Thymoquinone Inhibits Virulence Related Traits of Cronobacter sakazakii ATCC 29544 and Has Anti-biofilm Formation Potential. Front. Microbiol. 2018, 8, 2220. [Google Scholar] [CrossRef] [Green Version]
- Novy, P.; Kloucek, P.; Rondevaldova, J.; Havlik, J.; Kourimska, L.; Kokoska, L. Thymoquinone vapor significantly affects the results of Staphylococcus aureus sensitivity tests using the standard broth microdilution method. Fitoterapia 2014, 94, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Chaieb, K.; Kouidhi, B.; Jrah, H.; Mahdouani, K.; Bakhrouf, A. Antibacterial activity of Thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement. Altern. Med. 2011, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hariram, U.; Labbé, R.G. Growth and inhibition by spices of growth from spores of enterotoxigenic Bacillus cereus in cooked rice. Food Control 2016, 64, 60–64. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute (CLSI). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 8th ed.; Approved Standard M7-A10; LSI: Wayne, PA, USA, 2018. [Google Scholar]
- Shi, C.; Sun, Y.; Liu, Z.; Guo, D.; Sun, H.; Sun, Z.; Chen, S.; Zhang, W.; Wen, Q.; Peng, X.; et al. Inhibition of Cronobacter sakazakii Virulence Factors by Citral. Sci. Rep. 2017, 7, 43243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Chang, T.; Yang, H.; Cui, M. Antibacterial mechanism of lactic acid on physiological and morphological properties of Salmonella Enteritidis, Escherichia coli and Listeria monocytogenes. Food Control 2015, 47, 231–236. [Google Scholar] [CrossRef]
- Guo, D.; Wang, S.; Li, J.; Bai, F.; Yang, Y.; Xu, Y.; Liang, S.; Xia, X.; Wang, X.; Shi, C. The antimicrobial activity of coenzyme Q0 against planktonic and biofilm forms of Cronobacter sakazakii. Food Microbiol. 2020, 86, 103337. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Ma, X.; Li, Y.; Xu, H.; Han, X.; Wang, R.; Zhao, P.; Li, Z.; Shi, C. Antimicrobial Activity and Antibiofilm Potential of Coenzyme Q0 against Salmonella Typhimurium. Foods 2021, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Jeon, M.J.; Ha, J.W. Bactericidal and synergistic effects of X-ray irradiation and gallic acid against foodborne pathogens on lettuce. Food Microbiol. 2020, 92, 103584. [Google Scholar] [CrossRef]
- Li, G.; Wang, X.; Xu, Y.; Zhang, B.; Xia, X. Antimicrobial effect and mode of action of chlorogenic acid on Staphylococcus aureus. Eur. Food Res. Technol. 2013, 238, 589–596. [Google Scholar] [CrossRef]
- Kang, J.-H.; Song, K.B. Antibacterial activity of the noni fruit extract against Listeria monocytogenes and its applicability as a natural sanitizer for the washing of fresh-cut produce. Food Microbiol. 2019, 84, 103260. [Google Scholar] [CrossRef] [PubMed]
- Shu, Q.; Niu, Y.; Zhao, W.; Chen, Q. Antibacterial activity and mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus. Food Control 2019, 106, 106711. [Google Scholar] [CrossRef]
- Kong, L.B.; Zhang, P.F.; Yu, J.; Setlow, P.; Li, Y.Q. Monitoring the Kinetics of Uptake of a Nucleic Acid Dye during the Germination of Single Spores of Bacillus Species. Anal. Chem. 2010, 82, 8717–8724. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Sarkar, P.K. Bacillus cereus hazard and control in industrial dairy processing environment. Food Control 2016, 69, 20–29. [Google Scholar] [CrossRef]
- Fei, P.; Xu, Y.; Zhao, S.; Gong, S.; Guo, L. Olive oil polyphenol extract inhibits vegetative cells of Bacillus cereus isolated from raw milk. J. Dairy Sci. 2019, 102, 3894–3902. [Google Scholar] [CrossRef] [PubMed]
- Diao, M.; Qi, D.; Xu, M.; Lu, Z.; Lv, F.; Bie, X.; Zhang, C.; Zhao, H. Antibacterial activity and mechanism of monolauroyl-galactosylglycerol against Bacillus cereus. Food Control 2018, 85, 339–344. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Wu, X.; Sun, Y.; Liu, Z. Effect of thyme essential oil against Bacillus cereus planktonic growth and biofilm formation. Appl. Microbiol. Biotechnol. 2018, 102, 10209–10218. [Google Scholar] [CrossRef]
- Lv, X.; Miao, L.; Ma, H.; Bai, F.; Lin, Y.; Sun, M.; Li, J. Purification, characterization and action mechanism of plantaricin JY22, a novel bacteriocin against Bacillus cereus produced by Lactobacillus plantarum JY22 from golden carp intestine. Food Sci. Biotechnol. 2018, 27, 695–703. [Google Scholar] [CrossRef]
- Wang, X.; Tian, L.; Fu, J.; Liao, S.; Yang, S.; Jia, X.; Gong, G. Evaluation of the membrane damage mechanism of thymol against Bacillus cereus and its application in the preservation of skim milk. Food Control 2022, 131, 108435. [Google Scholar] [CrossRef]
- Bot, C.; Prodan, C. Probing the membrane potential of living cells by dielectric spectroscopy. Eur. Biophys. J. Biophys. Lett. 2009, 38, 1049–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, L.; Wang, Y.; Bi, X.; Duo, K.; Sun, Q.; Yun, X.; Zhang, Y.; Fei, P.; Han, J. Antimicrobial Activity and Mechanism of Action of the Amaranthus tricolor Crude Extract against Staphylococcus aureus and Potential Application in Cooked Meat. Foods 2020, 9, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plasek, J.; Gaskova, D.; Ludwig, J.; Hofer, M. Early changes in membrane potential of Saccharomyces cerevisiae induced by varying extracellular K(+), Na (+) or H (+) concentrations. J. Bioenerg. Biomembr. 2013, 45, 561–568. [Google Scholar] [CrossRef]
- Fei, P.; Ali, M.A.; Gong, S.; Sun, Q.; Bi, X.; Liu, S.; Guo, L. Antimicrobial activity and mechanism of action of olive oil polyphenols extract against Cronobacter sakazakii. Food Control 2018, 94, 289–294. [Google Scholar] [CrossRef]
- Mempin, R.; Tran, H.; Chen, C.; Gong, H.; Ho, K.K.; Lu, S.W. Release of extracellular ATP by bacteria during growth. BMC Microbiol. 2013, 13, 301. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Sun, Y.; Zhang, X.; Zheng, Z.; Yang, M.; Ben, H.; Song, K.; Cao, Y.; Chen, Y.; Liu, X.; et al. Antimicrobial effect of lipoic acid against Cronobacter sakazakii. Food Control 2016, 59, 352–358. [Google Scholar] [CrossRef]
- Tian, L.; Wang, X.; Zhang, D.; Wu, M.; Xue, Z.; Liu, Z.; Yang, S.; Li, H.; Gong, G. Evaluation of the membrane damage mechanism of protocatechualdehyde against Yersinia enterocolitica and simulation of growth inhibition in pork. Food Chem. 2021, 363, 130340. [Google Scholar] [CrossRef]
- Han, B.; Han, X.; Ren, M.; You, Y.; Zhan, J.; Huang, W. Antimicrobial Effects of Novel H2O2-Ag+ Complex on Membrane Damage to Staphylococcus aureus, Escherichia coli and Salmonella typhimurium. J. Food Prot. 2021. [Google Scholar] [CrossRef] [PubMed]
- Borisov, V.B.; Siletsky, S.A.; Nastasi, M.R.; Forte, E. ROS Defense Systems and Terminal Oxidases in Bacteria. Antioxidants 2021, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shan, M.; Zhu, Y.; Yao, H.; Li, H.; Gu, B.; Zhu, Z. Kalopanaxsaponin A induces reactive oxygen species mediated mitochondrial dysfunction and cell membrane destruction in Candida albicans. PLoS ONE 2020, 15, e0243066. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.G.; Chen, Y.J.; Tong, J.W.; Huang, J.A.; Li, J.; Gong, Y.S.; Liu, Z.H. Tea polyphenol epigallocatechin gallate inhibits Escherichia coli by increasing endogenous oxidative stress. Food Chem. 2017, 217, 196–204. [Google Scholar] [CrossRef]
- Vatansever, F.; de Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species--bactericidal antibiotics, photodynamic therapy, and beyond. Fems Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Song, K.; Zhang, X.; Sun, Y.; Sui, Y.; Chen, Y.; Jia, Z.; Sun, H.; Sun, Z.; Xia, X. Antimicrobial Activity and Possible Mechanism of Action of Citral against Cronobacter sakazakii. PLoS ONE 2016, 11, e0159006. [Google Scholar]
- Sun, X.H.; Zhou, T.T.; Wei, C.H.; Lan, W.Q.; Zhao, Y.; Pan, Y.J.; Wu, V.C. Antibacterial effect and mechanism of anthocyanin rich Chinese wild blueberry extract on various foodborne pathogens. Food Control 2018, 94, 155–161. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.; Qu, G.; Guo, D.; Yang, Y.; Ma, X.; Wang, M.; Xu, Y.; Wang, Y.; Xia, X.; et al. Antibacterial Activity and Mechanism of Coenzyme Q0 Against Escherichia coli. Foodborne Pathog. Dis. 2021, 18, 398–404. [Google Scholar] [CrossRef]
- Kim, M.J.; Han, J.K.; Park, J.S.; Lee, J.S.; Lee, S.H.; Cho, J.I.; Kim, K.S. Various Enterotoxin and Other Virulence Factor Genes Widespread Among Bacillus cereus and Bacillus thuringiensis Strains. J. Microbiol. Biotechnol. 2015, 25, 872–879. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Park, I.; Khan, I.; Alshammari, F.H.; Kumar, P.; Chen, L.; Na, M.; Huh, Y.S.; Han, Y.K.; Shukla, S. (-)-Tetrahydroberberrubineacetate accelerates antioxidant potential and inhibits food associated Bacillus cereus in rice. Food Chem. 2021, 339, 127902. [Google Scholar] [CrossRef] [PubMed]
- Voravuthikunchai, S.P.; Dolah, S.; Charernjiratraku, W. Control of Bacillus cereus in foods by Rhodomyrtus tomentosa (Ait.) Hassk. Leaf extract and its purified compound. J. Food Prot. 2010, 73, 1907–1912. [Google Scholar] [CrossRef] [PubMed]
- Christaki, S.; Moschakis, T.; Kyriakoudi, A.; Biliaderis, C.G.; Mourtzinos, I. Recent advances in plant essential oils and extracts: Delivery systems and potential uses as preservatives and antioxidants in cheese. Trends Food Sci. Technol. 2021, 116, 264–278. [Google Scholar] [CrossRef]
- Zanetti, M.; Carniel, T.K.; Dalcanton, F.; dos Anjos, R.S.; Riella, H.G.; de Araujo, P.H.H.; de Oliveira, D.; Fiori, M.A. Use of encapsulated natural compounds as antimicrobial additives in food packaging: A brief review. Trends Food Sci. Technol. 2018, 81, 51–60. [Google Scholar] [CrossRef]
- Zhang, P.; Kong, L.; Wang, G.; Setlow, P.; Li, Y.Q. Monitoring the wet-heat inactivation dynamics of single spores of Bacillus species by using Raman tweezers, differential interference contrast microscopy, and nucleic acid dye fluorescence microscopy. Appl. Environ. Microbiol. 2011, 77, 4754–4769. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Primers | Sequence of Primers (5′-3′) |
|---|---|---|
| 16S rRNA | Forward | AGAGTTTGATCMTGGCTCAG |
| Reverse | TACGGYTACCTTGTTACGACTT | |
| nheA | Forward | AGGTAAATGCGATGAGTAG |
| Reverse | TTGTTGAATGCGAAGAG | |
| nheB | Forward | CAAGCTCCAGTTCATGCGG |
| Reverse | GATCCCATTGTGTACCATTG | |
| nheC | Forward | TGGATTCCAAGATGTAACG |
| Reverse | ATTACGACTTCTGCTTGTGC | |
| Ces | Forward | TTGTTGGAATTGTCGCAGAG |
| Reverse | GTAAGCGAACCTGTCTGTAACAACA | |
| hblA | Forward | GCTAATGTAGTTTCACCTGTAGCAAC |
| Reverse | AATCATGCCACTGCGTGGACATATAA | |
| hblC | Forward | AATCAAGAGCTGTCACGAAT |
| Reverse | CACCAATTGACCATGCTAAT | |
| hblD | Forward | AATGGTCATCGGAACTCTAT |
| Reverse | CTCGCTGTTCTGCTGTTAAT | |
| bceT | Forward | GACTACATTCACGATTACGCAGAA |
| Reverse | CTATGCTGACGAGCTACATCCATA |
| Strains | Origin | MIC (μg/mL) | MBC (μg/mL) |
|---|---|---|---|
| ATCC 14579 | ATCC | 4.0 | 8.0 |
| CMCC 63301 | CMCC | 8.0 | 8.0 |
| CMCC 63303 | CMCC | 8.0 | 8.0 |
| BR1 | Rice | 4.0 | 8.0 |
| BR4 | Rice | 4.0 | 8.0 |
| M98 | Milk | 8.0 | 8.0 |
| C7 | Milk | 8.0 | 8.0 |
| C58 | Milk | 8.0 | 8.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Deng, H.; Wang, Y.; Rui, W.; Zhao, P.; Yong, Q.; Guo, D.; Liu, J.; Guo, X.; Wang, Y.; et al. Antimicrobial Activity and Action Mechanism of Thymoquinone against Bacillus cereus and Its Spores. Foods 2021, 10, 3048. https://doi.org/10.3390/foods10123048
Wang S, Deng H, Wang Y, Rui W, Zhao P, Yong Q, Guo D, Liu J, Guo X, Wang Y, et al. Antimicrobial Activity and Action Mechanism of Thymoquinone against Bacillus cereus and Its Spores. Foods. 2021; 10(12):3048. https://doi.org/10.3390/foods10123048
Chicago/Turabian StyleWang, Shuo, Haichao Deng, Yihong Wang, Wushuang Rui, Pengyu Zhao, Qiyao Yong, Du Guo, Jie Liu, Xinyi Guo, Yutang Wang, and et al. 2021. "Antimicrobial Activity and Action Mechanism of Thymoquinone against Bacillus cereus and Its Spores" Foods 10, no. 12: 3048. https://doi.org/10.3390/foods10123048
APA StyleWang, S., Deng, H., Wang, Y., Rui, W., Zhao, P., Yong, Q., Guo, D., Liu, J., Guo, X., Wang, Y., & Shi, C. (2021). Antimicrobial Activity and Action Mechanism of Thymoquinone against Bacillus cereus and Its Spores. Foods, 10(12), 3048. https://doi.org/10.3390/foods10123048