
Genotypic Characterization of Antimicrobial Resistant Salmonella spp. Strains from Three Poultry Processing Plants in Colombia

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
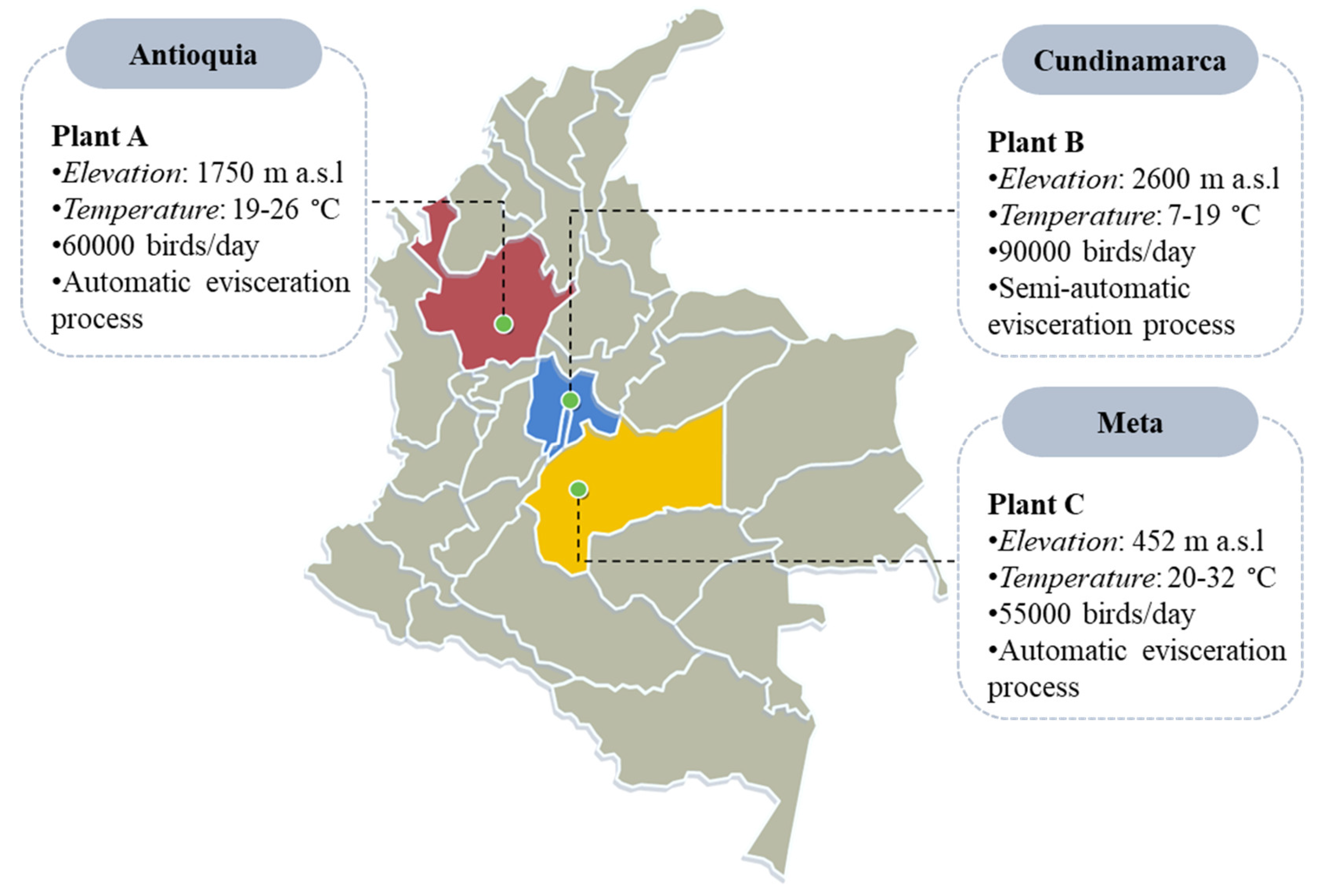
2.1. Characteristics of the Poultry Processing Plants
2.2. Experimental Design
2.3. Chicken Carcass Rinsate Sample Collection
2.4. Fecal Sample Collection
2.5. Microbiological Analysis
2.5.1. Salmonella spp. Detection and Isolation
2.5.2. Enumeration of Enterobacteriaceae
2.6. Whole Genome Sequencing (WGS)
2.7. Bioinformatic Analysis
2.8. Statistical Analysis
3. Results
3.1. Salmonella spp. Prevalence
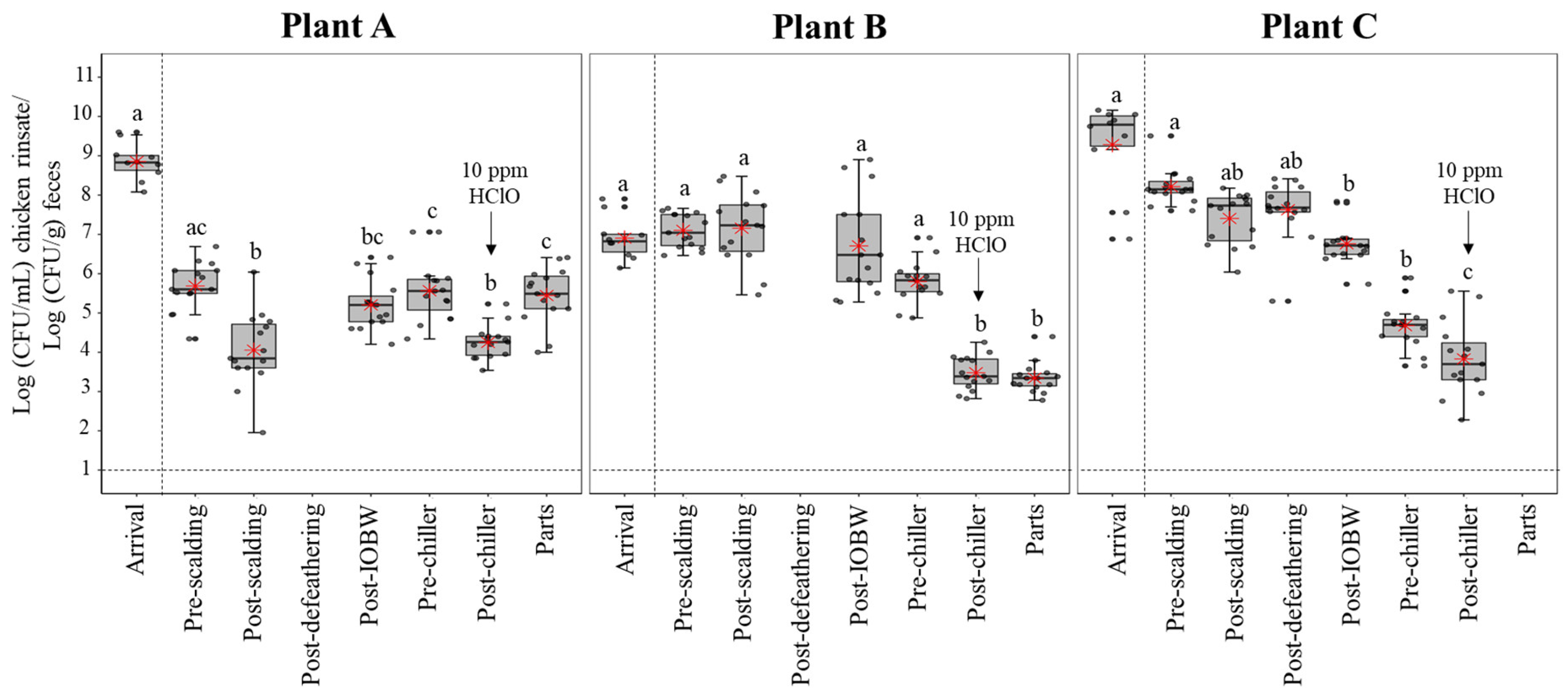
3.2. Enterobacteriaceae Levels
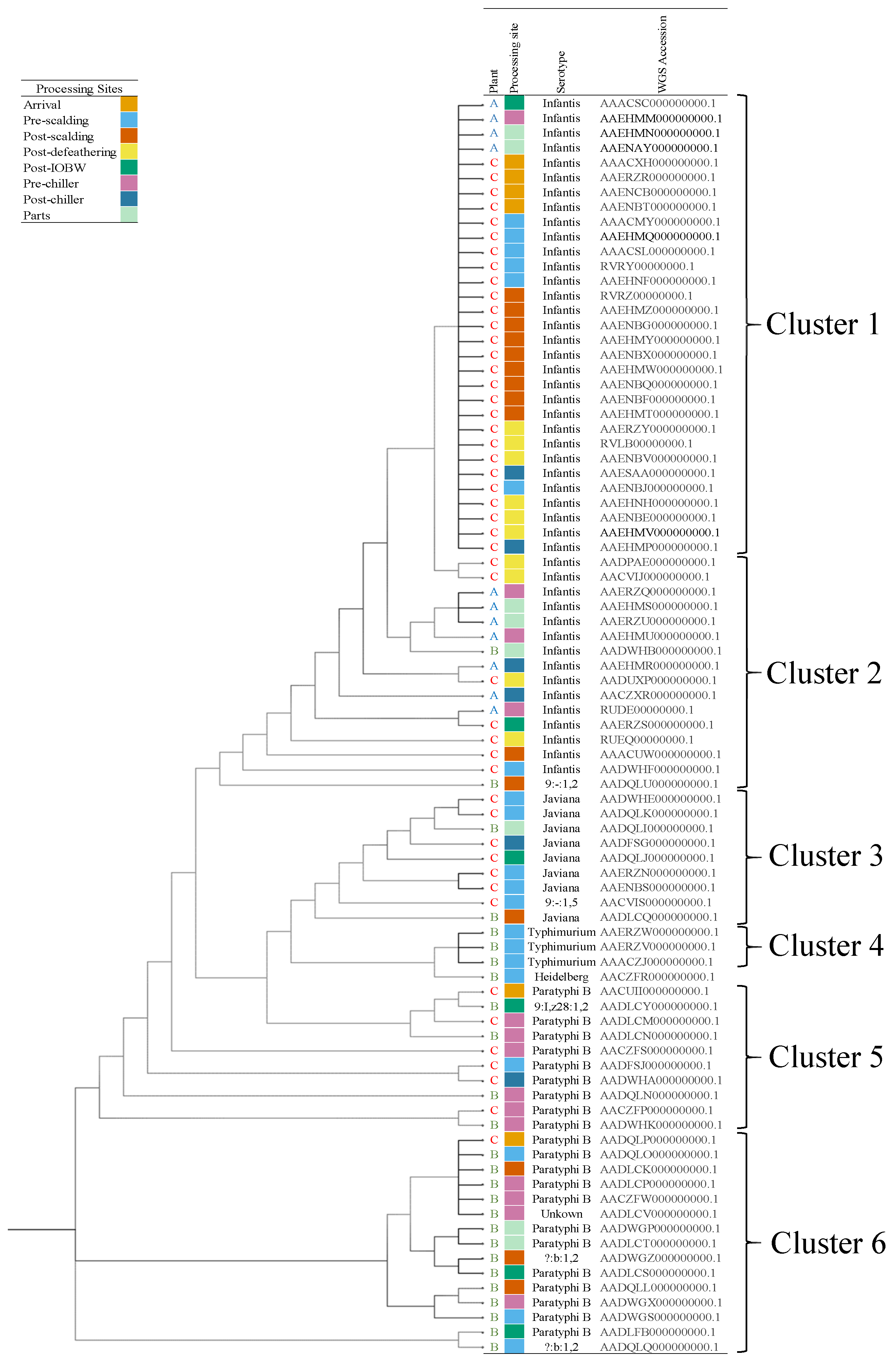
3.3. WGS of Salmonella spp. Isolates
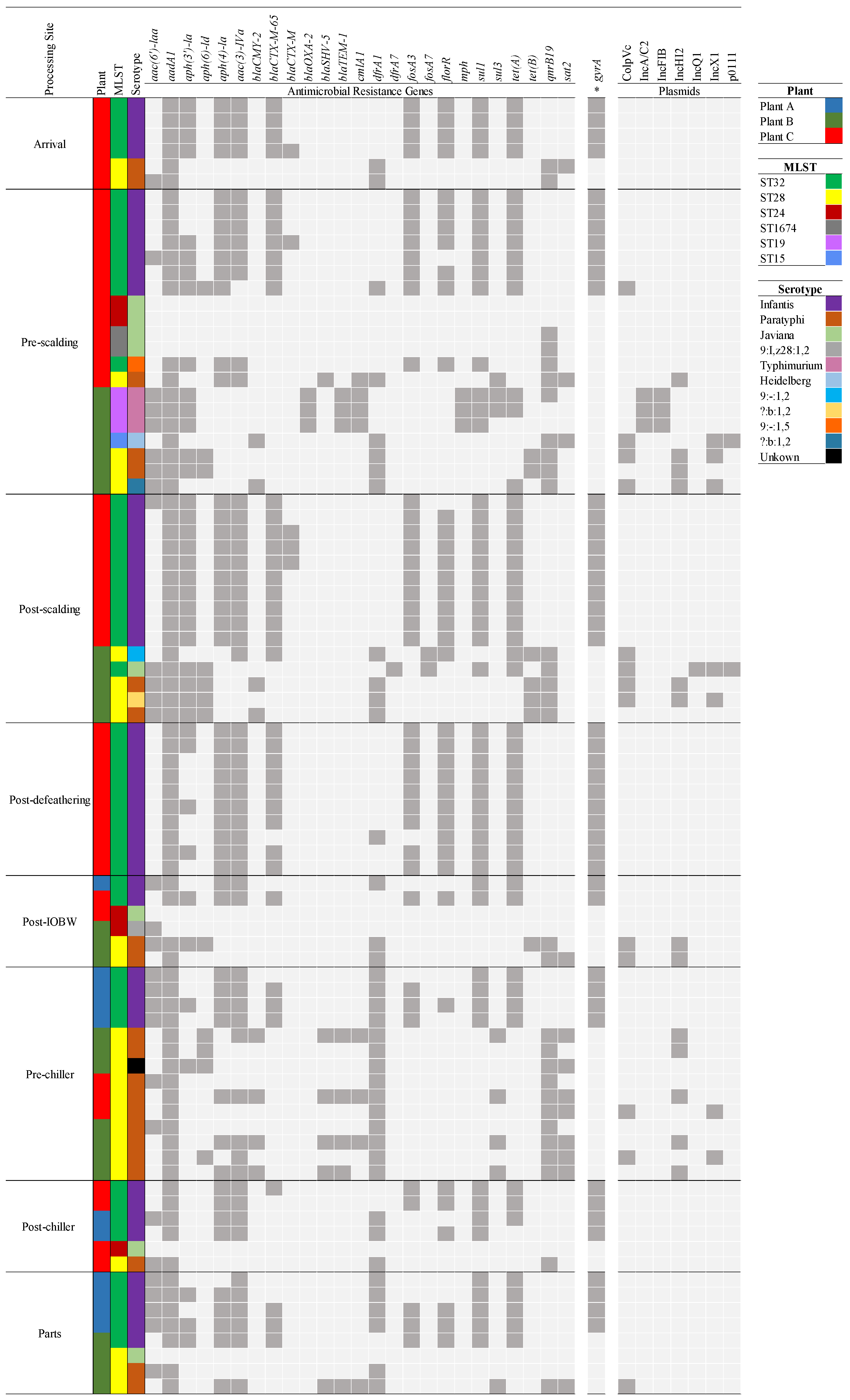
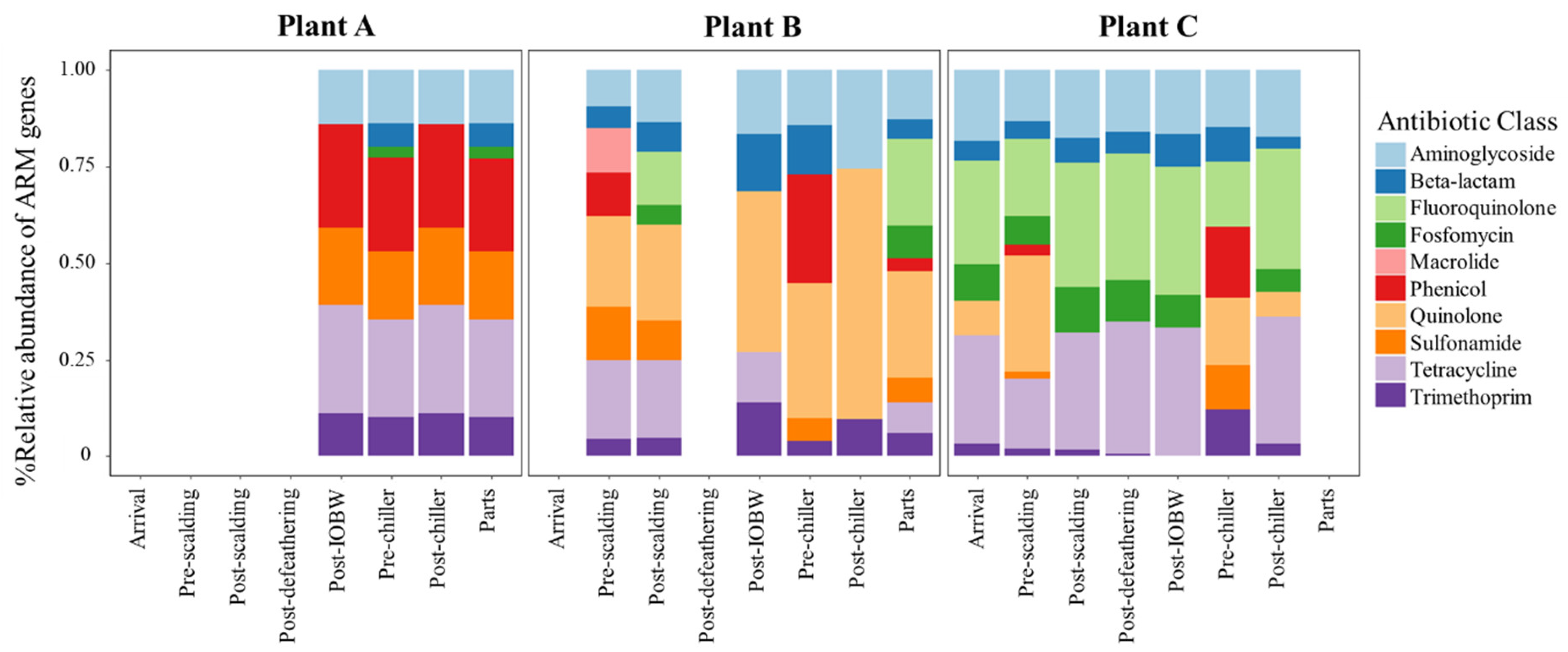
3.4. The Relative Abundance of AMR Genes within Processing Sites
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Procolombia. Colombian Poultry Sector: Features of the Colombia Poultry Sectore. 2016. Available online: https://compradores.procolombia.co/en/explore-business-opportunities/colombian-poultry-sector (accessed on 15 October 2018).
- Instituto Colombiano Agropecuario (ICA)/Colombian Agricultural Institute. Censo Pecuario Nacional año 2019/National Livestock Census Year. 2019. Available online: https://www.ica.gov.co/areas/pecuaria/servicios/epidemiologia-veterinaria/censos-2016/censo-2018 (accessed on 26 September 2018).
- Díaz, M.A. Determinantes del Desarrollo en la Avicultura en Colombia: Instituciones, Organizaciones y Tecnologías. Documento de Trabajo Sobre Economía Regional/Determinants Poultry Farming in Colombia Development: Institutions, Organizations and Technologies. White Paper on Regional Economy. 2014. Available online: http://www.banrep.gov.co/sites/default/files/publicaciones/archivos/dtser_214.pdf (accessed on 25 September 2018).
- Instituto Nacional de Vigilancia de Medicamentos y Alimentos (INVIMA)/National Institute of Food and Drug Surveillance. Plantas de Beneficio, Desposte, Desprese y Acondicionadores que Cumplen con los Requisitos Sanitarios Exigidos en el Decreto 1500 de 2007, decreto 2270 de 2012 y Resoluciones Reglamentarias/Processing Plants, Slaughtering Process, Cutting and Deboning to Comply with the Sanitary Requirements of Decree 1500 of 2007, Decree 2270 of 2012 and Regulatory Resolutions. 2019; Updated 20 August 2019. Available online: https://www.invima.gov.co/documents/20143/426809/PLANTAS-DE-BENEFICIO-DESPOSTE-Y-DESPRESE-AUTORIZADAS-POR-EL-INVIMA-PARA-SU-FUNCIONAMIENTO-A-JULIO-DE-2019-as-asp.pdf (accessed on 14 March 2020).
- Instituto Nacional de Salud (INS)/National Health Institute. Boletín Epidemiológico Semanal, las Enfermedades Transmitidas por Alimentos-ETA. Semana Epidemiológica 52/Weekly Epidemiological Bulletin, Foodborne Diseases. Epidemiological Week 52. 2018. Available online: https://www.ins.gov.co/buscador-eventos/BoletinEpidemiologico/2018%20Bolet%C3%ADn%20epidemiol%C3%B3gico%20semana%2052.pdf (accessed on 14 March 2020).
- Instituto Nacional de Salud (INS)/National Health Institute. Informe de Vigilancia por Laboratorio de Salmonella spp.: “Colombia 1997–2017”/Laboratory Surveillance for Salmonella spp.: “Colombia 1997–2017”. 2019. Available online: https://www.ins.gov.co/buscador-eventos/Informacin%20de%20laboratorio/Informe%20de%20vigilancia%20por%20laboratorio%20Salmonella%20spp%20Colombia%201997%202018.pdf (accessed on 18 November 2019).
- Ministerio de Salud y Protección Social, Instituto Nacional de Salud, Unidad de Evaluación de Riesgos Para la Inocuidad de Los alimentos/Ministry of Health and Social Protection, National Health Institute, Risk Assessment Unit for Food Safety. Perfil de riesgo Salmonella spp. (No Tifoideas) en Pollo Entero y en Piezas/Risk profile of Salmonella spp. in Broilers and Chicken Parts. 2011. Available online: https://www.minsalud.gov.co/sites/rid/Lists/BibliotecaDigital/RIDE/IA/INS/perfil-salmonella-spp.pdf (accessed on 15 October 2018).
- Ministerio de Salud y Protección Social, Instituto Nacional de Salud/Ministry of Health and Social Protection, National Health Institute. Perfil del Riesgo Campylobacter spp. en Pollos de Engorde/Risk Profile of Campylobacter spp. In Broilers. 2013. Available online: https://www.minsalud.gov.co/sites/rid/Lists/BibliotecaDigital/RIDE/IA/INS/Perfil-campylobacter-en-pollos.pdf (accessed on 9 October 2019).
- Ramirez-Hernandez, A.; Bugarel, M.; Kumar, S.; Thippareddi, H.; Brashears, M.M.; Sanchez-Plata, M.X. Phenotypic and genotypic characterization of antimicrobial resistance in Salmonella strains isolated from chicken carcasses and parts collected at different stages during processing. J. Food Protect. 2019, 82, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Hernandez, A. Assessing the Effect of Interventions on Pathogens and the Microbial Ecology of the Poultry Processing Chain by Microbial Profiling and the Phenotypic and Genotypic Characterization of Antimicrobial Resistance. Ph.D. Thesis, Texas Tech University, Lubbock, TX, USA, 2019. [Google Scholar]
- Keener, K.M.; Bashor, M.P.; Curtis, P.A.; Sheldon, B.W.; Kathariou, S. Campylobacter and Poultry Processing. Compr. Rev. Food Sci. Safe 2004, 3, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Byrd, J.A.; Rand McKee, S.R. Improving Slaughter and Processing Technologies. In Food Safety Control in the Poultry Industry; Mead, G.C., Ed.; CRC Press L.L.C.: Boca Raton, FL, USA, 2005; pp. 310–327. [Google Scholar]
- Nagel, G.M.; Bauermeister, L.J.; Bratcher, C.L.; Singh, M.; McKee, S.R. Salmonella and Campylobacter reduction and quality characteristics of poultry carcasses treated with various antimicrobials in a post-chill immersion tank. Int. J. Food Microbiol. 2013, 165, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.P.; Northcutt, J.K.; Cason, J.A.; Hinton, A., Jr.; Buhr, R.I.; Ingram, K.D. Effect of external or internal fecal contamination on numbers of bacteria on prechilled broiler carcasses. Poultry Sci. 2003, 86, 1241–1244. [Google Scholar] [CrossRef]
- Rouger, A.; Tresse, O.; Zagorec, M. Bacterial contaminants of poultry meat: Sources, species, and dynamics. Microorganisms 2017, 5, 50. [Google Scholar] [CrossRef]
- Vihavainen, E.J.; Björkroth, J. Microbial Ecology and Spoilage of Poultry Meat and Poultry Meat Products. In Handbook of Poultry Science and Technology, Secondary Processing; John Wiley & Sons, Inc: New Jersey, NY, USA, 2010; pp. 485–493. [Google Scholar]
- Donado-Godoy, P.; Clavijo, V.; León, M.; Tafur, M.A.; Gonzales, S.; Hume, M.; Alali, W.; Walls, I.; Lo Fo Wong, D.M.A.; Doyle, M.P. Prevalence of Salmonella on retail broiler chicken meat carcasses in Colombia. J. Food Protect. 2012, 75, 1134–1138. [Google Scholar] [CrossRef] [Green Version]
- Donado-Godoy, P.; Byrne, B.A.; Leon, M.; Castellanos, R.; Vanegas, C.; Coral, A.; Arévalo, A.; Clavijo, V.; Vargas, M.; Romero Zuñiga, J.J.; et al. Prevalence, resistance patterns, and risk factors for antimicrobial resistance in bacteria from retail chicken meat in Colombia. J. Food Protect. 2015, 78, 751–759. [Google Scholar] [CrossRef]
- Karczmarczyk, M.; Martins, M.; McCusker, M.; Mattar, S.; Amaral, L.; Leonard, N.; Aarestrup, F.N.; Fanning, S. Characterization of antimicrobial resistance in Salmonella enterica food and animal isolates from Colombia: Identification of a qnrB19-mediated quinolone resistance marker in two novel serovars. FEMS Microbiol. Lett. 2010, 313, 10–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Hernandez, A.; Varón-García, A.; Sanchez-Plata, M.X. Microbiological profile of three commercial poultry processing plants in Colombia. J. Food Protect. 2017, 80, 1980–1986. [Google Scholar] [CrossRef]
- Ciorba, V.; Odone, A.; Veronesi, L.; Pasquarella, C.; Signorelli, P. Antibiotic resistance as a major public health concern: Epidemiology and economic impact. Ann. Igiene Med. Prev. Comunita 2015, 27, 562–579. [Google Scholar]
- Dutil, L.; Irwin, R.; Finley, R.; Ng, L.K.; Avery, B.; Boerlin, P.; Bourgault, A.; Cole, L.; Daignault, D.; Desruisseau, A.; et al. Ceftiofur resistance in Salmonella enterica serovar Heidelberg from chicken meat and humans Canada. Emerg. Infect. Dis. 2010, 16, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Laxminarayan, R.; Matsoso, P.; Pant, S.; Brower, C.; Røttingen, J.A.; Klugman, K.; Davies, S. Access to effective antimicrobials: A worldwide challenge. Lancet 2016, 387, 168–175. [Google Scholar] [CrossRef]
- McEwen, S.A.; Fedorka-Cray, P.J. Antimicrobial use and resistance in animals and human beings. Clin. Infect. Dis. 2002, 34, S93–S106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donado-Godoy, P.; Castellanos, R.; León, M.; Arévalo, A.; Clavijo, V.; Bernal, J.; León, D.; Tafur, M.A.; Byrne, B.A.; Smith, W.A.; et al. The establishment of the Colombian integrated program for antimicrobial resistance surveillance (C.O.I.P.A.R.S.): A pilot project on poultry farms, slaughterhouses and retail market. Zoonoses Public Health 2015, 62, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Sefton, A.M. Mechanisms of antimicrobial resistance: Their clinical relevance in the new millennium. Drugs 2002, 62, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Sosa, A.D.J.; Byarugaba, D.K.; Amabile-Cuevas, C.F.; Hsueh, P.R.; Kariuki, S.; Okeke, I.N. Antimicrobial Resistance Mechanism in Developing Countries; Springer: New York, NY, USA, 2010. [Google Scholar]
- Li, Y.; Pulford, C.V.; Díaz, P.; Perez-Sepulveda, B.M.; Duarte, C.; Predeus, A.V.; Wiesner, M.; Heavens, D.; Low, R.; Shudoma, C.; et al. Genomic and phylogenetic analysis of Salmonella Typhimurium and its monophasic variants responsible for invasive endemic infections in Colombia. BioRxiv 2019. [Google Scholar] [CrossRef]
- Castro-Vargas, R.; Fandiño de Rubio, L.C.; Vega, A.; Rondón-Barragán, I. Phenotypic and genotypic resistance of Salmonella Heidelberg isolated from one of the largest poultry production region from Colombia. Int. J. Poult. Sci. 2019, 18, 610–617. [Google Scholar] [CrossRef] [Green Version]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nüesch-Inderbinen, M. Salmonella enterica serovar Infantis from Food and Human Infections, Switzerland, 2010–2015: Poultry-related multidrug resistant clones and an emerging ESBL producing clonal lineage. Front. Microbiol. 2017, 8, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marder, M.E.; Griffin, P.; Cieslak, P.; Dunn, J.; Hurd, S.; Jervis, R.; Lathrop, S.; Muse, A.; Ryan, P.; Smith, K.; et al. Preliminary incidence and trends of infections with pathogens transmitted commonly through food—Foodborne diseases active surveillance network, 10 U.S. sites, 2006-2017. MMWR-Morbid. Mortal Wkly. Rep. 2018, 23, 324–328. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Outbreak of Multidrug-Resistant Salmonella Infections liNked to Raw Chicken Products. 2019. Available online: https://www.cdc.gov/salmonella/infantis-10-18/index.html (accessed on 15 March 2020).
- Tate, H.; Folster, J.P.; Hsu, C.H.; Chen, J.; Hoffmann, M.; Li, C.; Morales, C.; Tyson, G.H.; Mukherjee, S.; Brown, A.C.; et al. Comparative analysis of extended-spectrum-β-lactamase CTX-M-65-producing Salmonella enterica serovar Infantis isolates from humans, food animals, and retail chickens in the United States. Antimicrob. Agents Chemother 2017, 61. [Google Scholar] [CrossRef] [Green Version]
- Nógrády, N.; Király, M.; Davies, B.; Nagy, B. Multidrug resistant clones of Salmonella Infantis of broiler origin in Europe. Int. J. Food Microbiol. 2012, 157, 108–112. [Google Scholar] [CrossRef]
- Burke, L.; Hopkins, K.L.; Meunier, D.; De Pinna, E.; Fitzgerald-Hughes, D.; Humphreys, H.; Woodford, N. Resistance to third-generation cephalosporins in human non-typhoidal Salmonella enterica isolates from England and Wales, 2010–2012. J. Antimicrob. Chemother. 2014, 69, 977–981. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Di Giannatale, E.; et al. Emergence of a clonal lineage of multidrug-resistant ESBL-producing Salmonella Infantis transmitted from broilers and broiler meat to humans in Italy between 2011 and 2014. PLoS ONE 2015, 10, e0144802. [Google Scholar] [CrossRef] [Green Version]
- Cartelle-Gestal, M.; Zurita, J.; Paz-Mino, A.; Ortega-Paredes, D.; Alcocer, I. Characterization of a small outbreak of Salmonella enterica serovar Infantis that harbor CTX-M-65 in Ecuador. Braz. J. Infect. Dis. 2005, 2, 406–407. [Google Scholar]
- Castellanos, L.R.; van der Graaf-van Bloois, L.; Donado-Godoy, P.; León, M.; Clavijo, V.; Arévalo, A.; Bernal, J.F.; Mevius, D.J.; Wagenaar, J.A.; Zomer, A.; et al. Genomic characterization of extended-spectrum cephalosporin-resistant Salmonella enterica in the Colombian poultry chain. Front. Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Donado-Godoy, P.; Clavijo, V.; Leon, M.; Arevalo, A.; Castellanos, R.; Bernal, J.; Tafur, M.A.; Ovalle, M.V.; Alali, M.Q.; Hume, M.; et al. Counts, serovars, and antimicrobial resistance phenotypes of Salmonella on raw chicken meat at retail in Colombia. J. Food Protect. 2014, 77, 227–235. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Processing Site | Plant A | Plant B | Plant C | |||
|---|---|---|---|---|---|---|
| No. Positive (%) | 95% CI | No. Positive (%) | 95% CI | No. Positive (%) | 95% CI | |
| Fecal material | ||||||
| Arrival | 1/10 (10) | 0.5–46 | 0/1 (0) | 0–34 | 4/10 (40) | 14–73 |
| Chicken carcass rinse | ||||||
| Pre-scalding | 2/15 (13) a | 23–42 | 12/15 (80) a | 51–97 | 12/15 (80) a | 51–97 |
| Post-scalding | 14/15 (93) b | 66–99 | 8/15 (53) a | 27–78 | 15/15 (100) a | 75–100 |
| Post-defeathering | - | - | 13/15 (87) a | 58–98 | ||
| Post-IOBW (inside-outside body wash) | 15/15 (100) b | 75–100 | 6/15 (40) a | 17–67 | 11/15 (73) a | 45–91 |
| Pre-chiller | 15/15 (100) b | 75–100 | 12/15 (80) a | 51–97 | 13/15 (87) a | 58–98 |
| Post-chiller | 15/15 (100) b | 75–100 | 7/15 (47) a | 22–73 | 12/15 (80) a | 51–97 |
| Parts | 15/15 (100) b | 75–100 | 13/15 (87) a | 58–98 | - | |
| Processing Site | Plant A | Plant B | Plant C |
|---|---|---|---|
| (n = 11) | (n = 26) | (n = 48) | |
| Arrival | Infantis (4) | ||
| Paratyphi B (2) | |||
| Pre-scalding | Typhimurium (3) | Infantis (7) | |
| Heidelberg (1) | Javiana (4) | ||
| Paratyphi B (2) | 9:-:1,5 * (1) | ||
| ?:b:1,2 * (1) | Paratyphi B (1) | ||
| Post-scalding | 9:-:1,2 * (1) | Infantis (10) | |
| Javiana (1) | |||
| ?:b:1,2 * (1) | |||
| Paratyphi B (2) | |||
| Post-defeathering | Infantis (10) | ||
| Post-IOBW | Infantis (1) | 9:I,z28:1,2 * (1) | Infantis (1) |
| Paratyphi B (2) | Javiana (1) | ||
| Pre-chiller | Infantis (4) | Paratyphi B (6) | Paratyphi B (3) |
| Unknown * (1) | |||
| Post-chiller | Infantis (2) | Infantis (2) | |
| Javiana (1) | |||
| Paratyphi B (1) | |||
| Parts | Infantis (4) | Infantis (1) | |
| Javiana (1) | |||
| Paratyphi B (2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramirez-Hernandez, A.; Carrascal-Camacho, A.K.; Varón-García, A.; Brashears, M.M.; Sanchez-Plata, M.X. Genotypic Characterization of Antimicrobial Resistant Salmonella spp. Strains from Three Poultry Processing Plants in Colombia. Foods 2021, 10, 491. https://doi.org/10.3390/foods10030491
Ramirez-Hernandez A, Carrascal-Camacho AK, Varón-García A, Brashears MM, Sanchez-Plata MX. Genotypic Characterization of Antimicrobial Resistant Salmonella spp. Strains from Three Poultry Processing Plants in Colombia. Foods. 2021; 10(3):491. https://doi.org/10.3390/foods10030491
Chicago/Turabian StyleRamirez-Hernandez, Alejandra, Ana K. Carrascal-Camacho, Andrea Varón-García, Mindy M. Brashears, and Marcos X. Sanchez-Plata. 2021. "Genotypic Characterization of Antimicrobial Resistant Salmonella spp. Strains from Three Poultry Processing Plants in Colombia" Foods 10, no. 3: 491. https://doi.org/10.3390/foods10030491
APA StyleRamirez-Hernandez, A., Carrascal-Camacho, A. K., Varón-García, A., Brashears, M. M., & Sanchez-Plata, M. X. (2021). Genotypic Characterization of Antimicrobial Resistant Salmonella spp. Strains from Three Poultry Processing Plants in Colombia. Foods, 10(3), 491. https://doi.org/10.3390/foods10030491