
Impact of Nisin and Nisin-Producing Lactococcus lactis ssp. lactis on Clostridium tyrobutyricum and Bacterial Ecosystem of Cheese Matrices



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures
2.2. Spore Production
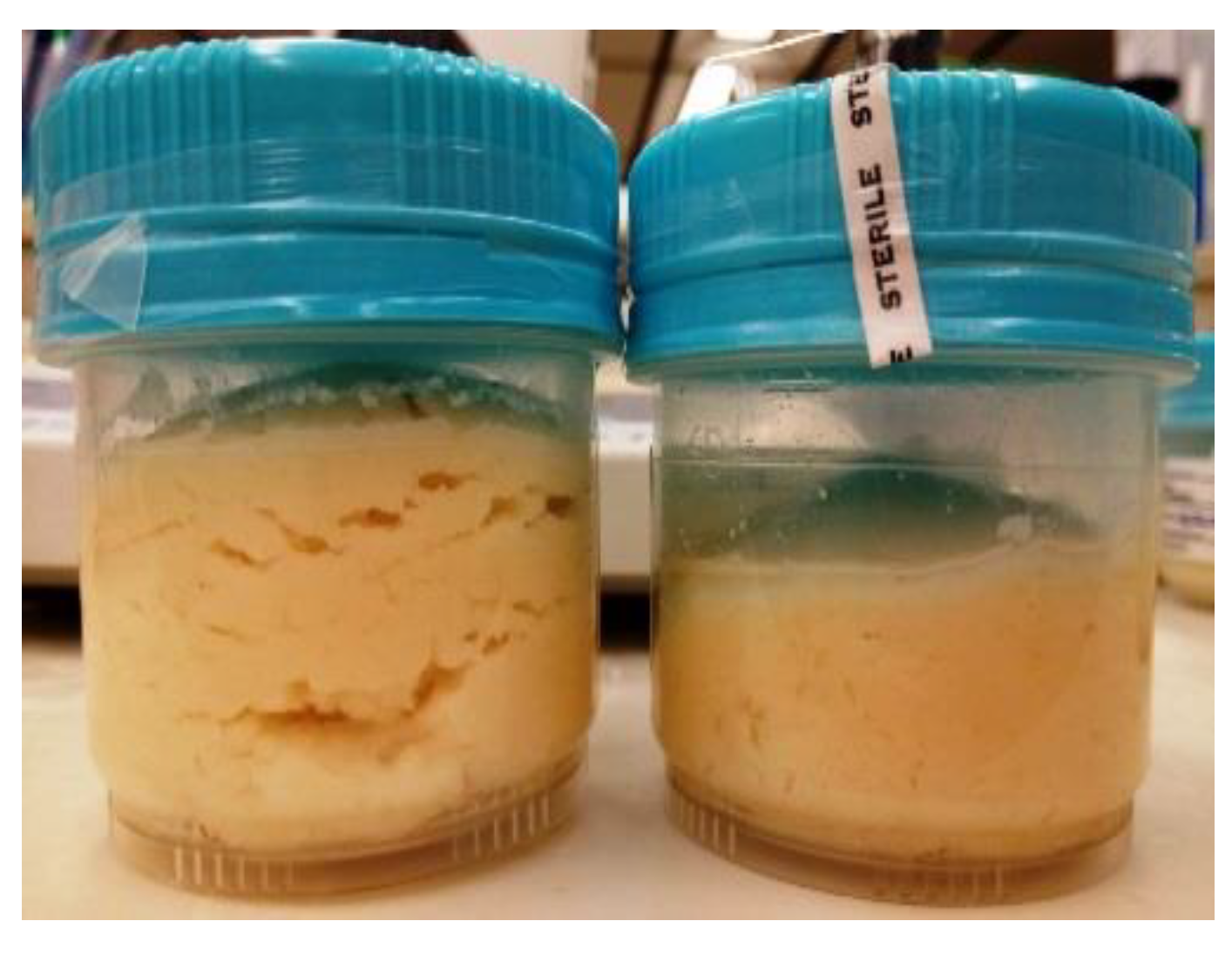
2.3. Pearce Test
2.4. Nisin Encapsulation
2.5. Cheese Slurry Preparation and Experimental Treatments
- Group 1: Cheddar cheese starter (8 log10 cfu/g) + C. tyrobutyricum (4 log10 cfu/g)
- Group 2: Cheddar cheese starter (8 log10 cfu/g) + C. tyrobutyricum (4 log10 cfu/g) + encapsulated nisin
- Group 3: Cheddar cheese starter (8 log10 cfu/g) + C. tyrobutyricum (4 log10 cfu/g) + L. lactis ssp. lactis 32 (nisin A producer)
2.6. Nisin Determination by HPLC
2.7. Metagenomic Analysis
2.7.1. Sample Preparation
2.7.2. DNA Extraction from Cheese Samples
2.7.3. 16S rRNA Sequencing and Analysis
2.8. Statistical Analysis
3. Results
3.1. Selection of Mixed Culture Ratio and Production of Encapsulated Nisin
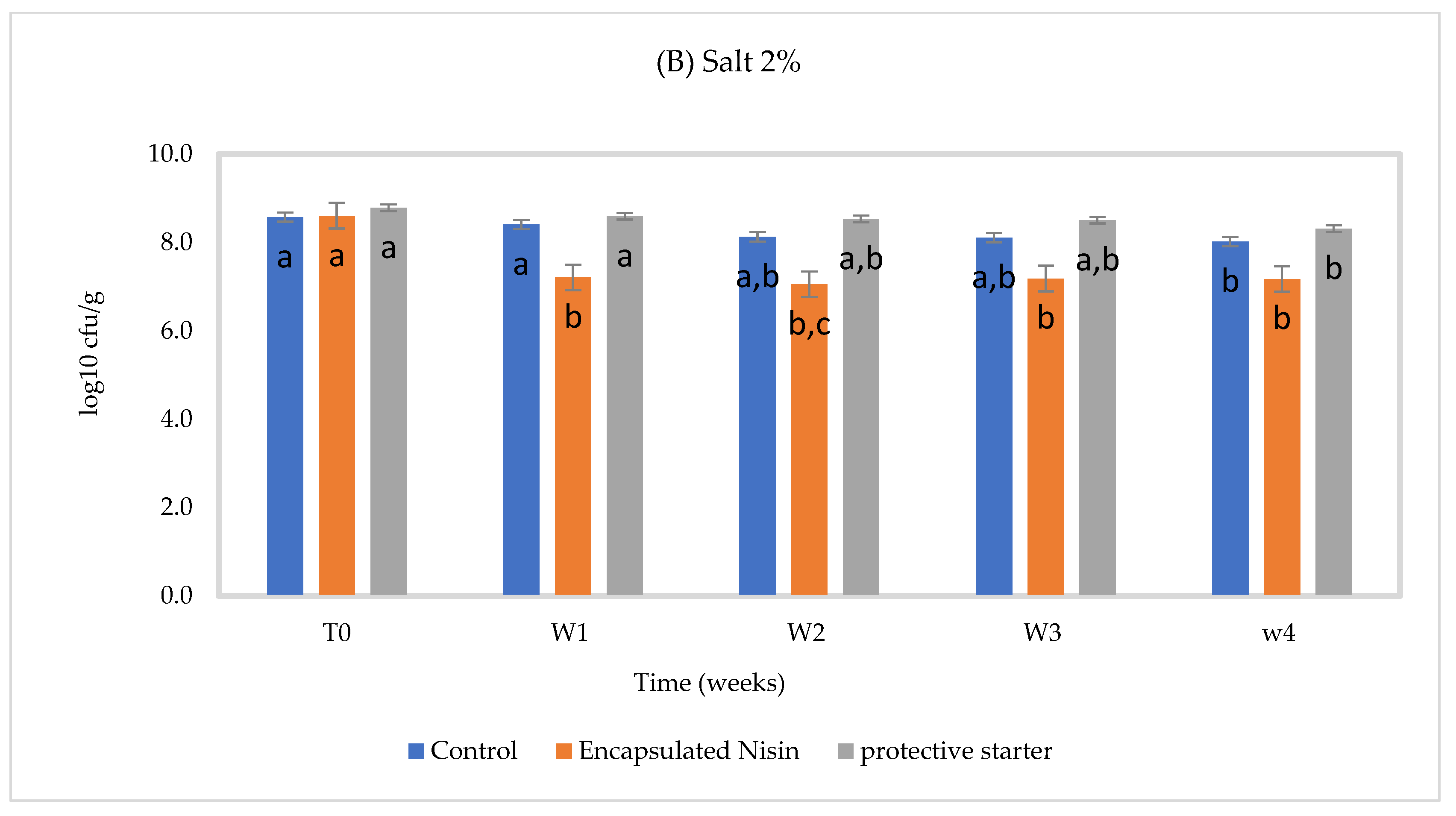
3.2. Chemical and Microbiological Composition of Cheese Slurry
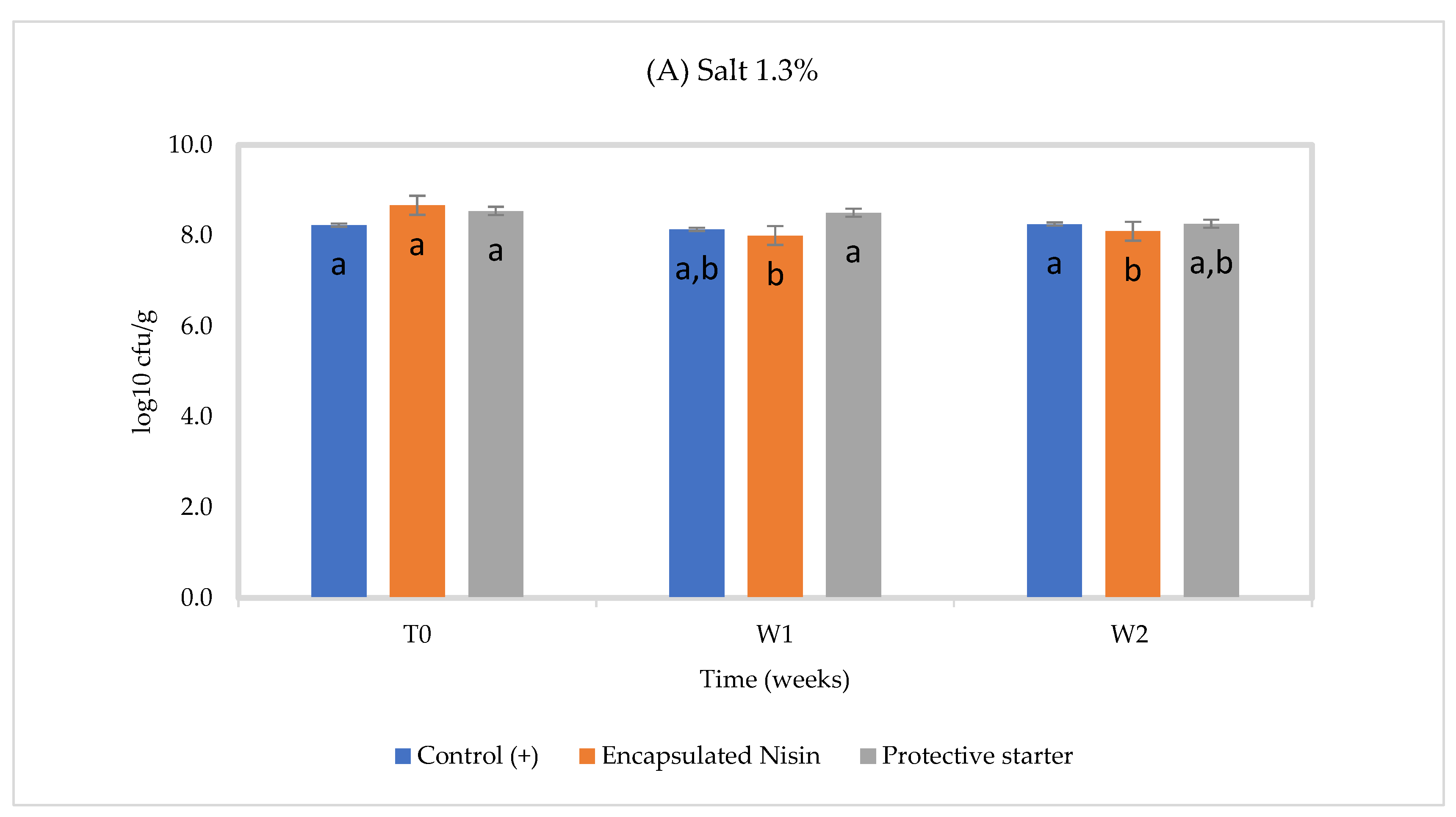
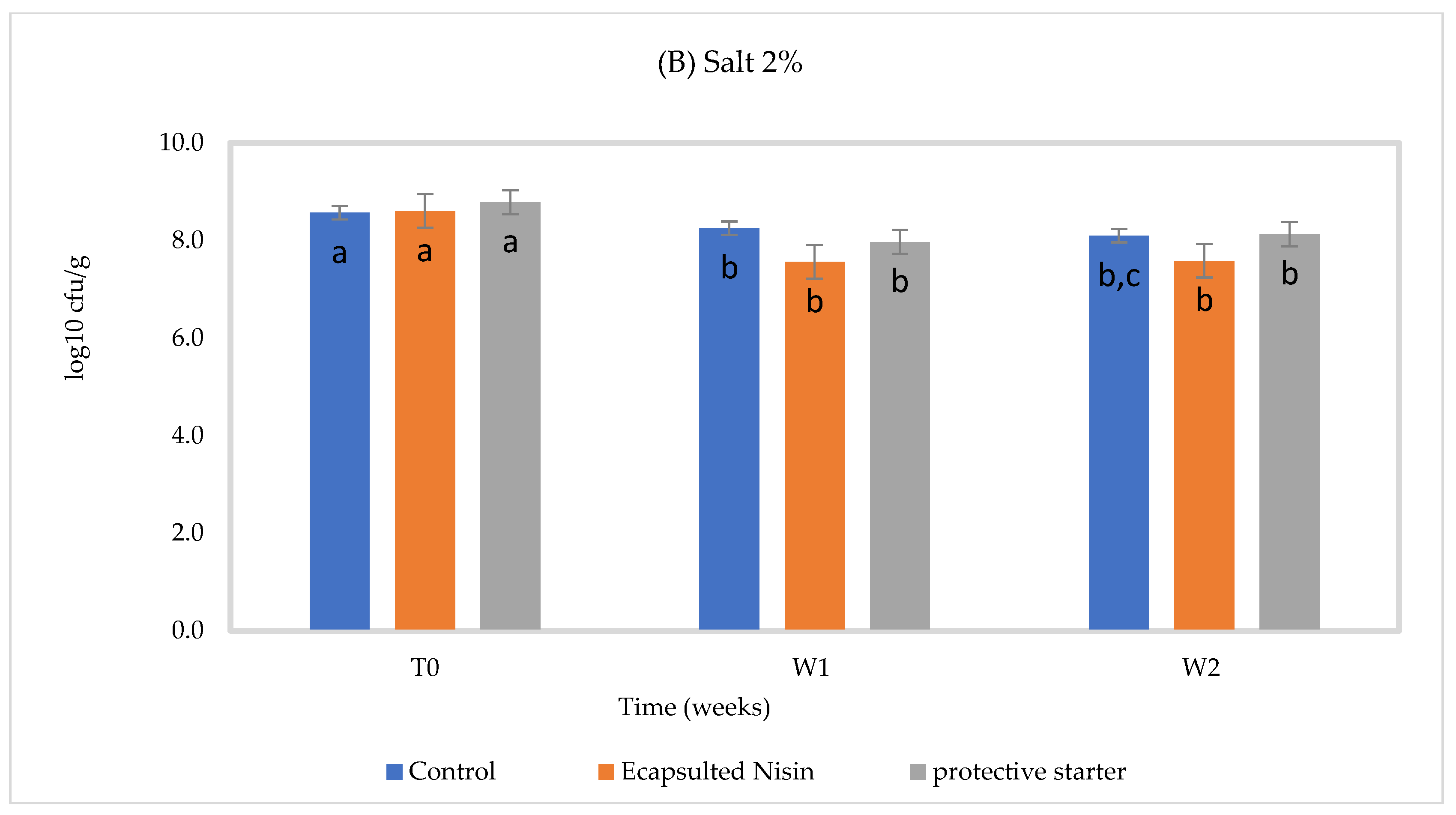
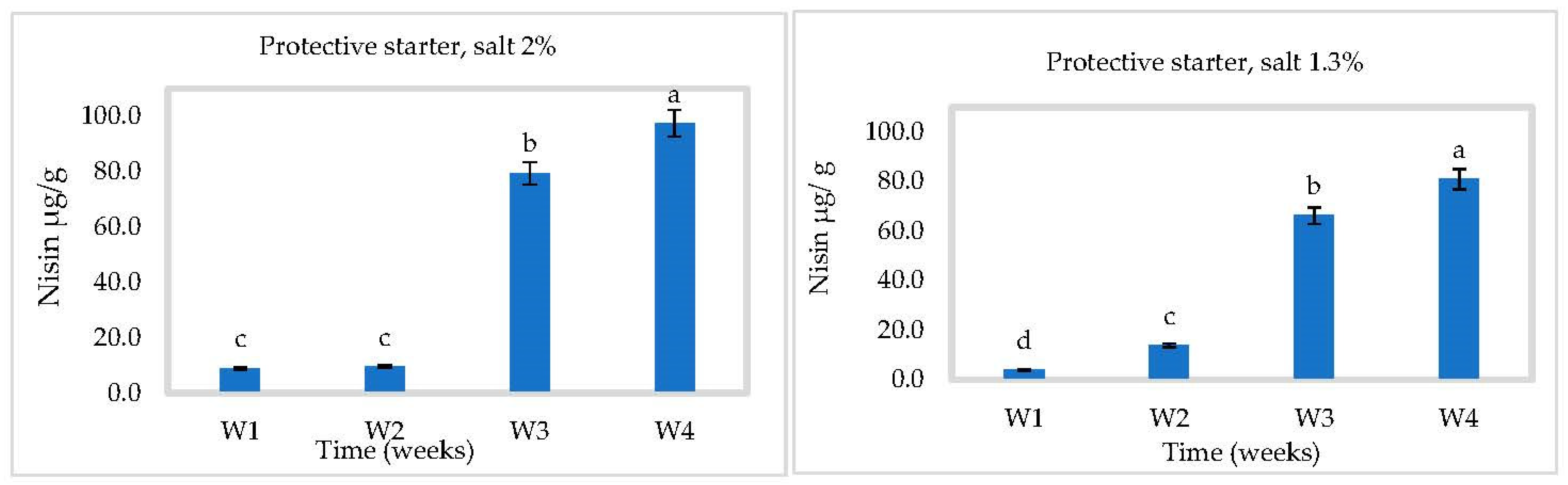
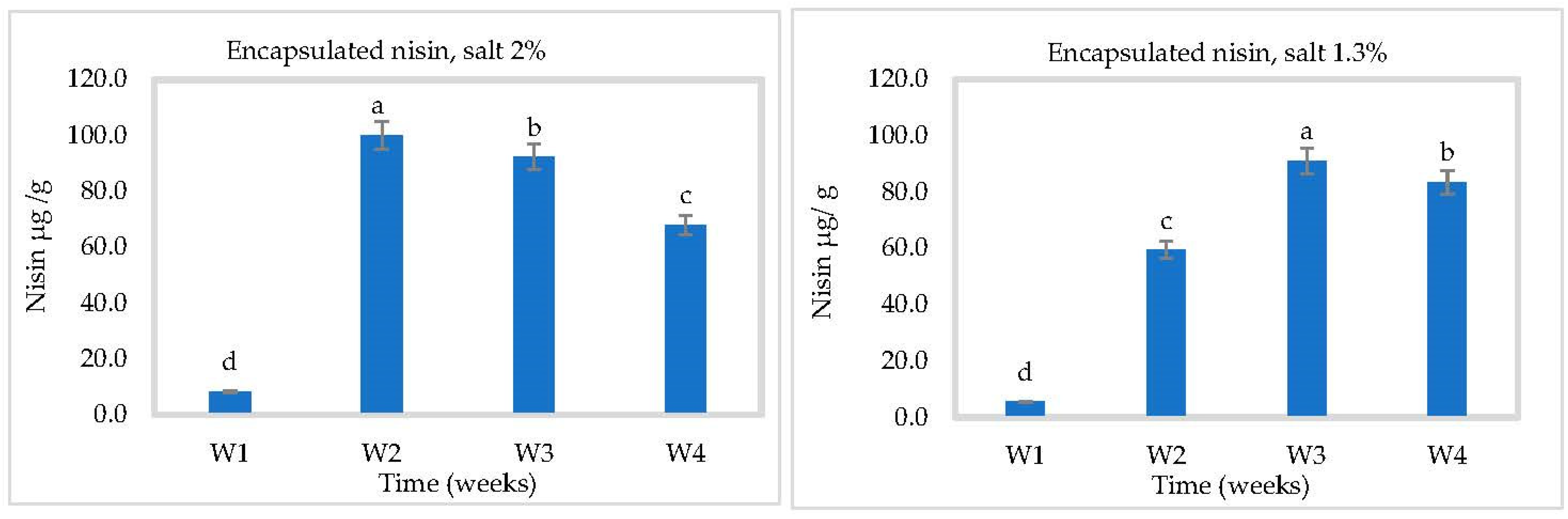
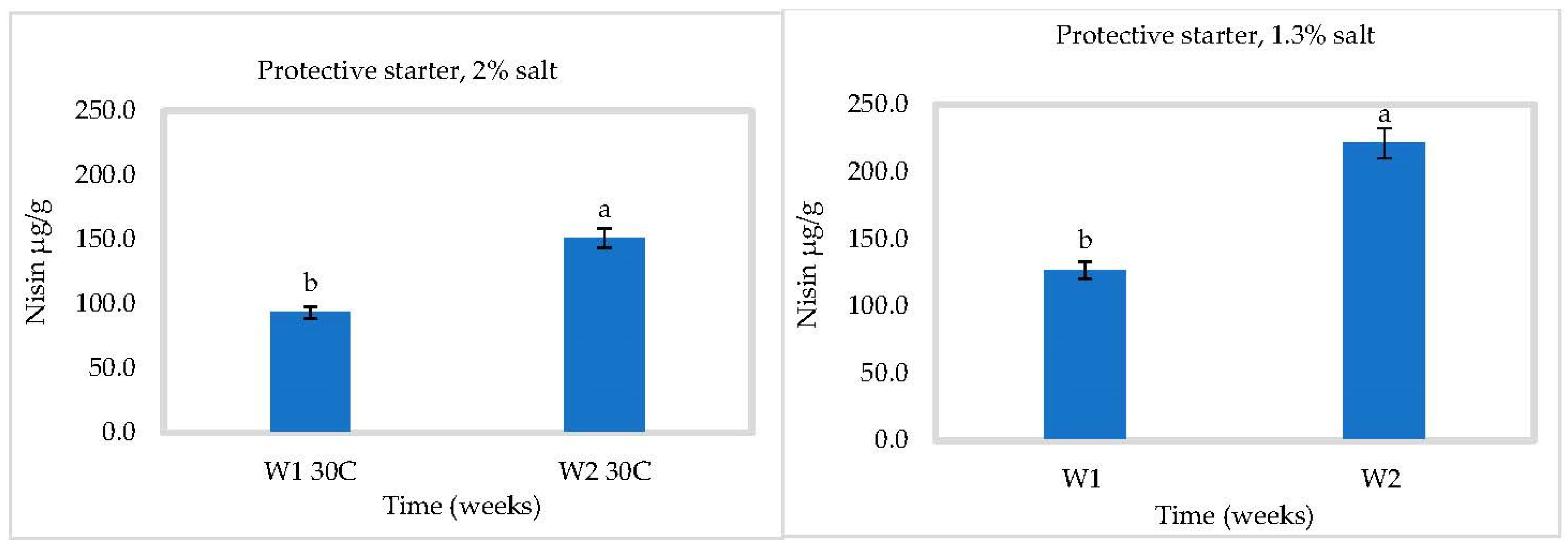
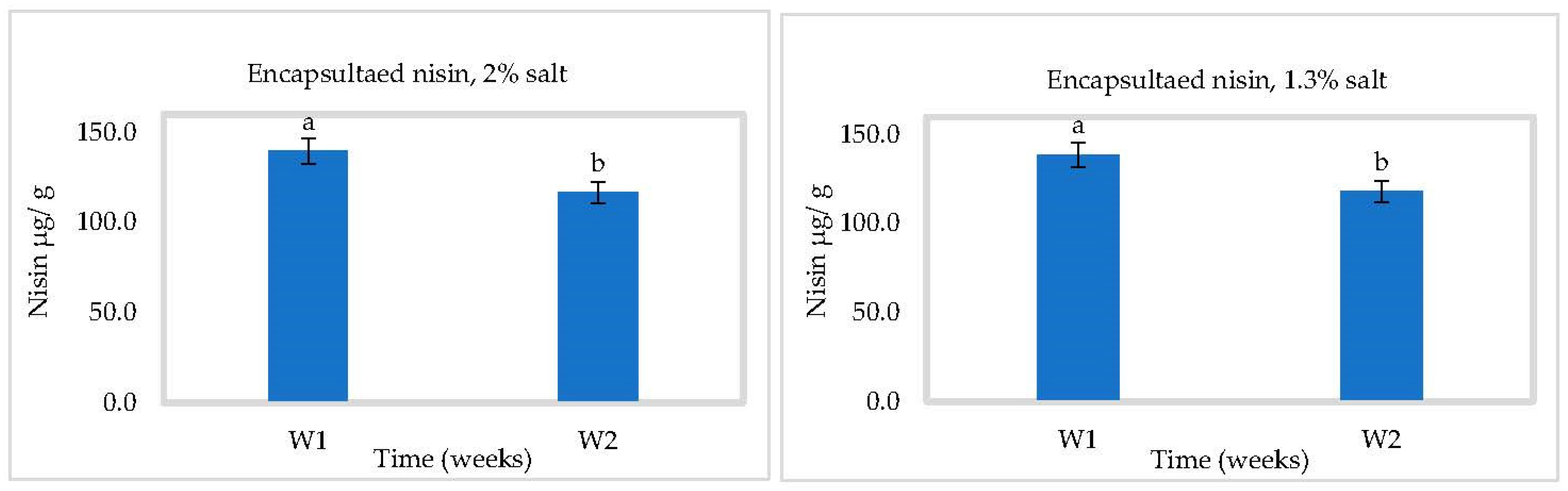
3.3. In Situ Nisin Production
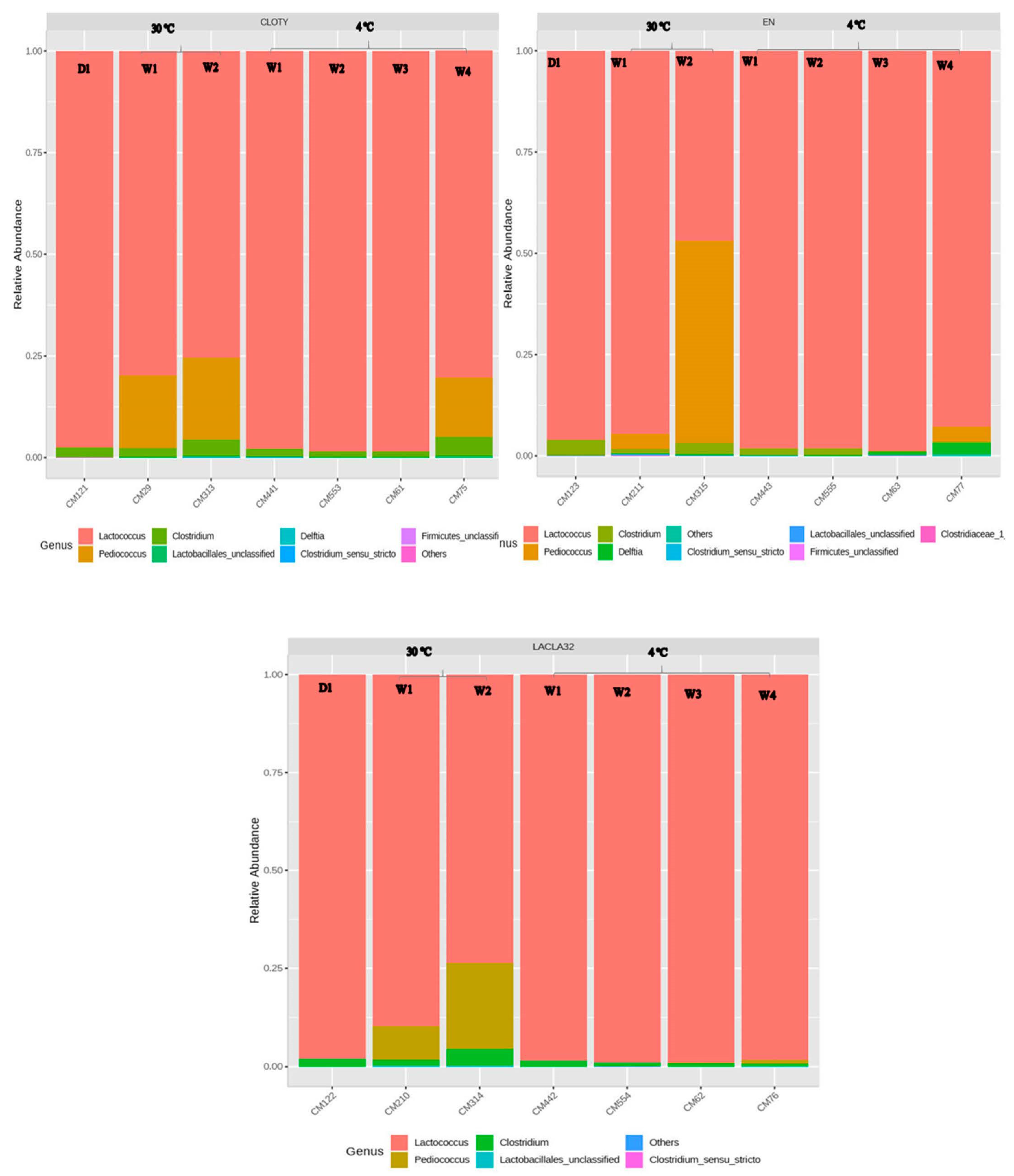
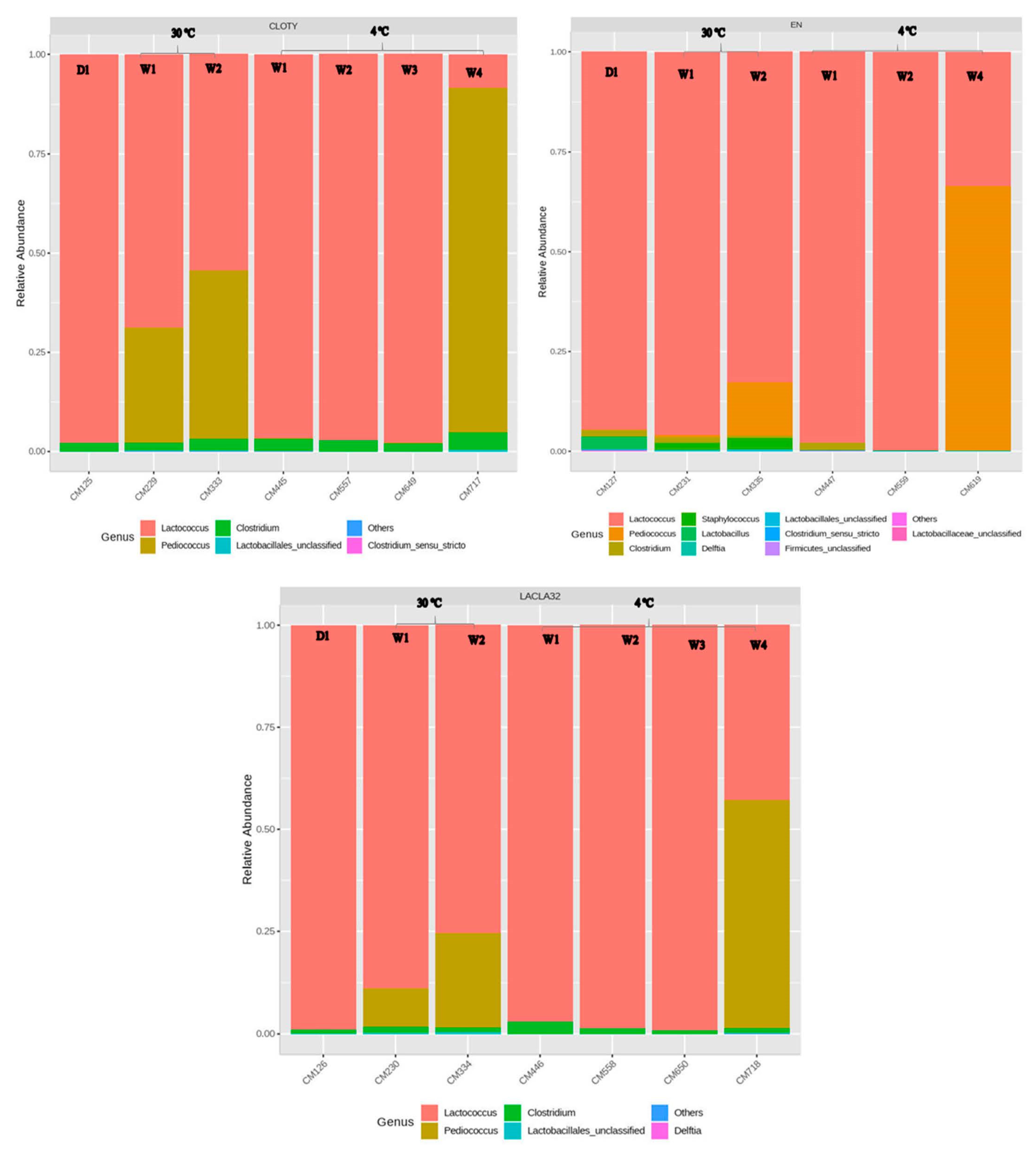
3.4. Metagenomics Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bermúdez, J.; González, M.J.; Olivera, J.A.; Burgueño, J.A.; Juliano, P.; Fox, E.M.; Reginensi, S.M. Seasonal occurrence and molecular diversity of clostridia species spores along cheesemaking streams of 5 commercial dairy plants. J. Dairy Sci. 2016, 99, 3358–3366. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Torres, N.; Garde, S.; Peirotén, Á.; Ávila, M. Impact of Clostridium spp. on cheese characteristics: Microbiology, color, formation of volatile compounds and off-flavors. Food Control 2015, 56, 186–194. [Google Scholar]
- McSweeney, P.; Fox, P. Metabolism of residual lactose and of lactate and citrate. In Cheese: Chemistry, Physics, and Microbiology; Elsevier: Amsterdam, The Netherlands, 2004; Volume 1, pp. 361–371. [Google Scholar]
- Klijn, N.; Nieuwenhof, F.; Hoolwerf, J.D.; van der Waals, C.; Weerkamp, A.H. Identification of Clostridium tyrobutyricum as the causative agent of late blowing in cheese by species-specific PCR amplification. Appl. Environ. Microbiol. 1995, 61, 2919–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández García, L.; Blanco, S.Á.; Rodriguez, F.A.R. Microfiltration applied to dairy streams: Removal of bacteria. J. Sci. Food Agric. 2013, 93, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Ávila, M.; Gómez-Torres, N.; Hernández, M.; Garde, S. Inhibitory activity of reuterin, nisin, lysozyme and nitrite against vegetative cells and spores of dairy-related Clostridium species. Int. J. Food Microbiol. 2014, 172, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, G.; Meijer, W.; Düsterhöft, E.-M.; Smit, G. Gouda, and related cheeses. In Cheese: Chemistry, Physics and Microbiology; Elsevier: Amsterdam, The Netherlands, 2004; pp. 103–140. [Google Scholar]
- Lodi, R.; Stadhouders, J. The use of lysozyme to control butyric acid fermentation. Bull. Int. Dairy Fed. 1990, 251, 51–54. [Google Scholar]
- Stadhouders, J. Prevention of butyric acid fermentation by the use of nitrate. Bull. Int. Dairy Fed. 1990, 251, 40–46. [Google Scholar]
- D’Incecco, P.; Faoro, F.; Silvetti, T.; Schrader, K.; Pellegrino, L. Mechanisms of Clostridium tyrobutyricum removal through natural creaming of milk: A microscopy study. J. Dairy Sci. 2015, 98, 5164–5172. [Google Scholar] [CrossRef] [PubMed]
- Fremont, S.; Kanny, G.; Nicolas, J.; Moneret-Vautrin, D. Prevalence of lysozyme sensitization in an egg-allergic population. Allergy 1997, 52, 224–228. [Google Scholar] [CrossRef]
- Dugat-Bony, E.; Sarthou, A.-S.; Perello, M.-C.; de Revel, G.; Bonnarme, P.; Helinck, S. The effect of reduced sodium chloride content on the microbiological and biochemical properties of a soft surface-ripened cheese. J. Dairy Sci. 2016, 99, 2502–2511. [Google Scholar] [CrossRef]
- Appel, L.J.; Angell, S.Y.; Cobb, L.K.; Limper, H.M.; Nelson, D.E.; Samet, J.M.; Brownson, R.C. Population-wide sodium reduction: The bumpy road from evidence to policy. Ann. Epidemiol 2012, 22, 417–425. [Google Scholar] [CrossRef] [Green Version]
- Cruz, A.G.; Faria, J.A.; Pollonio, M.A.; Bolini, H.M.; Celeghini, R.M.; Granato, D.; Shah, N.P. Cheeses with reduced sodium content: Effects on functionality, public health benefits and sensory properties. Trends Food Sci. Technol. 2011, 22, 276–291. [Google Scholar] [CrossRef]
- Reis, J.; Paula, A.; Casarotti, S.; Penna, A. Lactic acid bacteria antimicrobial compounds: Characteristics and applications. Food Eng. Rev. 2012, 4, 124–140. [Google Scholar] [CrossRef]
- Gálvez, A.; Abriouel, H.; López, R.L.; Omar, N.B. Bacteriocin-based strategies for food biopreservation. Int. J. Food Microbiol. 2007, 120, 51–70. [Google Scholar] [CrossRef]
- Jack, R.W.; Tagg, R.J.; Ray, B. Bacteriocins of gram-positive bacteria. Microbiol. Mol. Biol. Rev. 1995, 59, 171–200. [Google Scholar] [CrossRef]
- Delves-Broughton, J. Natural antimicrobials as additives and ingredients for the preservation of foods and beverages. In Natural Food Additives, Ingredients and Flavourings; Elsevier: Amsterdam, The Netherlands, 2012; pp. 127–161. [Google Scholar]
- Shin, J.M.; Gwak, J.W.; Kamarajan, P.; Fenno, J.C.; Rickard, A.H.; Kapila, Y.L. Biomedical applications of nisin. J. Appl. Microbiol. 2016, 120, 1449–1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verheul, A.; Russell, N.J.; Hof, R.V.T.; Rombouts, F.M.; Abee, T. Modifications of membrane phospholipid composition in nisin-resistant Listeria monocytogenes Scott. A. Appl. Environ. Microbiol. 1997, 63, 3451–3457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benech, R.-O.; Kheadr, E.; Laridi, R.; Lacroix, C.; Fliss, I. Inhibition of Listeria innocua in cheddar cheese by addition of nisin Z in liposomes or by in situ production in mixed culture. Appl. Environ. Microbiol. 2002, 68, 3683–3690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi-Zhang, Y.; Yam, K.L.; Chikindas, M.L. Effective control of Listeria monocytogenes by combination of nisin formulated and slowly released into a broth system. Int. J. Food Microbiol. 2004, 90, 15–22. [Google Scholar] [CrossRef]
- de Arauz, L.J.; AJozala, F.; Mazzola, P.G.; Penna, T.C.V. Nisin biotechnological production and application: A review. Trends Food Sci. Technol. 2009, 20, 146–154. [Google Scholar] [CrossRef]
- Hassan, H.; Gomaa, A.; Subirade, M.; Kheadr, E.; St-Gelais, D.; Fliss, I. Novel design for alginate/resistant starch microcapsules controlling nisin release. Int. J. Biol. Macromol. 2019, 15, 1186–1192. [Google Scholar] [CrossRef]
- Okereke, A.; Montville, T.J. Bacteriocin inhibition of Clostridium botulinum spores by lactic acid bacteria. J. Food Prot. 1991, 54, 349–353. [Google Scholar] [CrossRef]
- Gardner-Fortier, C.; St-Gelais, D.; Champagne, C.P.; Vuillemard, J.-C. Determination of optimal conditions for γ-aminobutyric acid production by Lactococcus lactis ssp. lactis. Int. Dairy J. 2013, 32, 136–143. [Google Scholar] [CrossRef]
- Bullard, J.; St-Gelais, D.; Turgeon, S.L. Production of set yoghurts using thermophilic starters composed of two strains with different growth biocompatibilities and producing different exopolysaccharides. Int. Dairy J. 2018, 79, 33–42. [Google Scholar] [CrossRef]
- Desfossés-Foucault, É.; Dussault-Lepage, V.; le Boucher, C.; Savard, P.; LaPointe, G.; Roy, D. Assessment of probiotic viability during Cheddar cheese manufacture and ripening using propidium monoazide-PCR quantification. Front. Microbiol. 2012, 3, 350. [Google Scholar] [CrossRef] [Green Version]
- Davidson, P.; Cekmer, H.B.; Monu, E.; Techathuvanan, C. The use of natural antimicrobials in food: An overview. In Handbook of Natural Antimicrobials for Food Safety and Quality; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1–27. [Google Scholar]
- Silva, C.C.; Silva, S.P.; Ribeiro, S.C. Application of bacteriocins and protective cultures in dairy food preservation. Front. Microbiol. 2018, 9, 594. [Google Scholar] [CrossRef]
- Roberts, R.F.; Zottola, E.; McKay, L. Use of a nisin-producing starter culture suitable for Cheddar cheese manufacture. J. Dairy Sci. 1992, 75, 2353–2363. [Google Scholar] [CrossRef]
- Nerandzic, M.M.; Donskey, C.J. Activate to eradicate inhibition of Clostridium difficile spore outgrowth by the synergistic effects of osmotic activation and nisin. PLoS ONE 2013, 8, e54740. [Google Scholar] [CrossRef] [Green Version]
- Garde, S.; Ávila, M.; Arias, R.; Gaya, P.; Nuñez, M. Outgrowth inhibition of Clostridium beijerinckii spores by a bacteriocin-producing lactic culture in ovine milk cheese. Int. J. Food Microbiol. 2011, 150, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Ávila, M.; Gómez-Torres, N.; Gaya, P.; Garde, S. Effect of a nisin-producing lactococcal starter on the late blowing defect of cheese caused by Clostridium tyrobutyricum. Int. J. Food Sci. Technol. 2020, 55, 3343–3349. [Google Scholar] [CrossRef]
- Rilla, N.; Martínez, B.; Delgado, T.; Rodríguez, A. Inhibition of Clostridium tyrobutyricum in Vidiago cheese by Lactococcus lactis ssp. lactis IPLA 729, a nisin Z producer. Int. J. Food Microbiol. 2003, 85, 23–33. [Google Scholar] [CrossRef]
- Shrestha, S.; Grieder, J.; McMahon, D.J.; Nummer, B. Survival of Listeria monocytogenes introduced as a post-aging contaminant during storage of low-salt Cheddar cheese at 4, 10, and 21 °C. J. Dairy Sci. 2011, 94, 4329–4335. [Google Scholar] [CrossRef] [PubMed]
- Kamimura, B.A.; de Filippis, F.; Sant’Ana, A.S.; Ercolini, D. Large-scale mapping of microbial diversity in artisanal Brazilian cheeses. Food Microbiol. 2019, 80, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Duthoit, F.; Callon, C.; Tessier, L.; Montel, M.-C. Relationships between sensorial characteristics and microbial dynamics in “Registered Designation of Origin” Salers cheese. Int. J. Food Microbiol. 2005, 103, 259–270. [Google Scholar] [CrossRef] [PubMed]
- Erkus, O.; de Jager, V.C.; Geene, R.T.; van Alen-Boerrigter, I.; Hazelwood, L.; van Hijum, S.A.; Kleerebezem, M.; Smid, E.J. Use of propidium monoazide for selective profiling of viable microbial cells during Gouda cheese ripening. Int. J. Food Microbiol. 2016, 228, 1–9. [Google Scholar] [CrossRef] [PubMed]














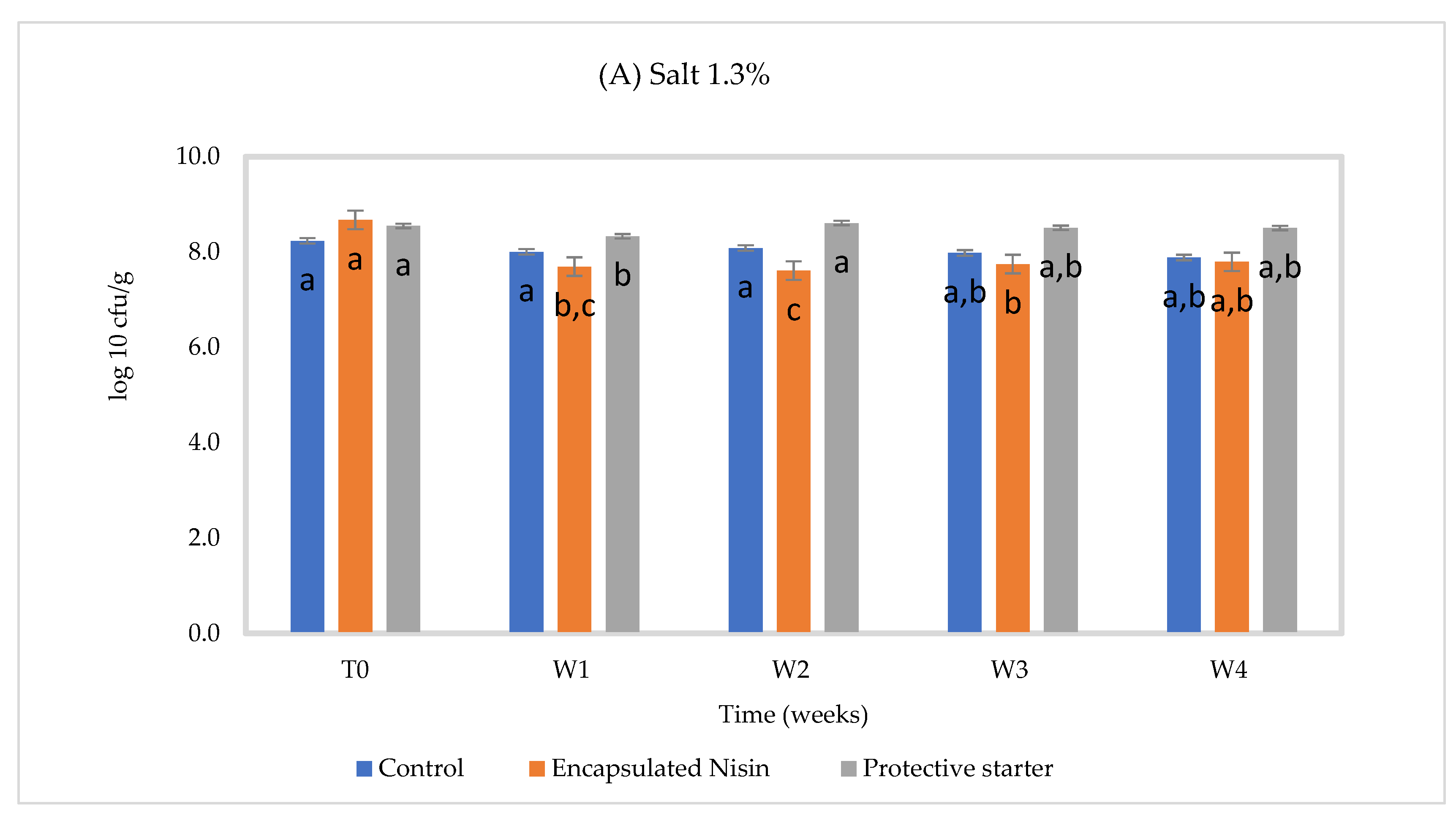{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g) | pH 5.3 | pH 5.00 | ||
|---|---|---|---|---|
| 1% NaCl | 2% NaCl | 1% NaCl | 2% NaCl | |
| Cheese powder | 180 | 180 | 180 | 180 |
| NaCl solutions pH 2.0 | 102.9 (NaCl 3.2%) | 102.9 (NaCl 3.8%) | 102.9 (NaCl 3.2%) | 102.9 (NaCl 3.8%) |
| Starter culture | 6 | 6 | 6 | 6 |
| Clostridium tyrobutyricum | 6 | 6 | 6 | 6 |
| Phosphate buffer | 5.1 or 0.6 a,b | 5.1 or 0.6 a,b | 5.1 or 0.6 a,b | 5.1 or 0.6 a,b |
| Nisin capsules (EN) a | 4.5 | 4.5 | 4.5 | 4.5 |
| Protective strain b | 4.5 | 4.5 | 4.5 | 4.5 |
| Total weight | 300 | 300 | 300 | 300 |
| Target moisture content | 38% | 38% | 38% | 38% |
| pH | |||
|---|---|---|---|
| Time Zero | 2 h | 5 h | |
| Lactococcus lactis ssp. lactis 32 | 6.54 d | 6.51 b | 5.76 b |
| Lactococcus lactis ssp. cremoris CUC 222 | 6.55 c | 6.51 b | 5.83 a |
| Lactococcus lactis ssp. lactis CUC-H | 6.57 a | 6.52 a | 5.53 g |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 (1:1) | 6.57 a | 6.51 b | 5.67 c |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. lactis CUC-H (1:1) | 6.56 b | 6.5 c | 5.48 h |
| Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1:1) | 6.55 c | 6.5 c | 5.48 h |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1:1:1) | 6.55 c | 6.5 c | 5.56 e |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1.5:1:1) | 6.55 c | 6.49 d | 5.55 f |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1.5:1:1) 2% | 6.55 c | 6.44 f | 5.23 i |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (2:1:1) | 6.55 c | 6.48 e | 5.56 e |
| Viable Count Log10 cfu/mL | |||
|---|---|---|---|
| Time Zero | 2 h | 5 h | |
| Lactococcus lactis ssp. lactis 32 | 6.10 ± 0.01 d | 7.46 ± 0.02 c | 8.69 ± e |
| Lactococcus lactis ssp. cremoris CUC 222 | 5.40 ± 0.02 h | 5.70 ± 0.03 f | 7.63 ± 0.02 g |
| Lactococcus lactis ssp. lactis CUC-H | 6.44 ± 0.02 b | 7.40 ± 0.01 d | 8.92 ± 0.02 a |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 (1:1) | 6.00 ± 0.03 e | 7.40 ± 0.02 d | 8.60 ± 0.02 f |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. lactis CUC-H (1:1) | 6.26 ± 0.01 c | 7.49 ± 0.01 b,c | 8.86 ± b |
| Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1:1) | 5.85 ± 0.02 f | 7.21 ± 0.03 e | 8.79 ± 0.01 d |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1:1:1) | 5.70 ± 0.03 g | 7.51 ± 0.03 b | 8.78 ± 0.01 d |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1.5:1:1) | 6.01 ± 0.01 e | 7.41 ± 0.01 d | 8.83 ± 0.01 c |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1.5:1:1) 2% | 6.24 ± 0.04 c | 7.81 ± 0.0 a | 8.87 ± 0.01 b |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (2:1:1) | 6.60 ± 0.01 a | 7.46 ± 0.01c | 8.80 ± 0.01 d |
| Nisin-A (AU/mL) | |||
|---|---|---|---|
| Time Zero | 2 h | 5 h | |
| L. lactis 32 | ND | ND | 64 |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 (1:1) | ND | ND | 64 |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. lactis CUC-H (1:1) | ND | ND | 64 |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1:1:1) | ND | ND | 64 |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1.5:1:1) | ND | ND | 64 |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (1.5:1:1) 2% | ND | ND | 256 |
| Lactococcus lactis ssp. lactis 32 + Lactococcus lactis ssp. cremoris CUC222 + Lactococcus lactis ssp. lactis CUC-H (2:1:1) | ND | ND | 64 |
| pH 5.0 and 1.3% salt | ||||
| Groups | NaCl % | Moisture % | Salt/Moisture (S/M) | pH |
| Control (+) | 1.36 ± 0.01 | 38.49 ± 0.1 | 3.52 ± 0.03 | 5.10 ± 0.01 |
| Encapsulated nisin | 1.25 ± 0.01 | 38.53 ± 0.1 | 2.87 ± 0.01 | 5.12 ± 0.01 |
| protective strain | 1.27 ± 0.01 | 38.42 ± 0.2 | 3.32 ± 0.01 | 5.00 ± 0.01 |
| pH 5.0 and 2% salt | ||||
| Groups | NaCl % | Moisture % | (S/M) | pH |
| Control (+) | 2.01 ± 0.01 | 37.37 ± 0.02 | 5.38 ± 0.01 | 5.00 ± 0.02 |
| Encapsulated nisin | 1.96 ± 0.01 | 38.26 ± 0.03 | 5.12 ± 0.02 | 5.10 ± 0.01 |
| protective strain | 1.86 ± 0.01 | 37.65 ± 0.02 | 4.94 ± 0.01 | 5.00 ± 0.01 |
| pH 5.3 and 1.3% salt | ||||
| Groups | NaCl % | Moisture % | (S/M) | pH |
| Control (+) | 1.34 ± 0.01 | 37.66 ± 0.02 | 3.56 ± 0.01 | 5.25 ± 0.01 |
| Encapsulated nisin | 1.33 ± 0.01 | 39.03 ± 0.01 | 3.41 ± 0.01 | 5.24 ± 0.01 |
| protective strain | 1.23 ± 0.01 | 38.30 ± 0.02 | 3.21 ± 0.01 | 5.24 ± 0.01 |
| pH 5.3 and 2% salt | ||||
| Groups | NaCl % | Moisture % | (S/M) | pH |
| Control (+) | 1.88 ± 0.02 | 37.19 ± 0.03 | 5.05 ± 0.03 | 5.26 ± 0.05 |
| Encapsulated nisin | 1.87 ± 0.02 | 37.37 ± 0.02 | 5.20 ± 0.02 | 5.28 ± 0.01 |
| protective starter | 1.87 ± 0.02 | 37.38 ± 0.01 | 5.00 ± 0.01 | 5.28 ± 0.01 |
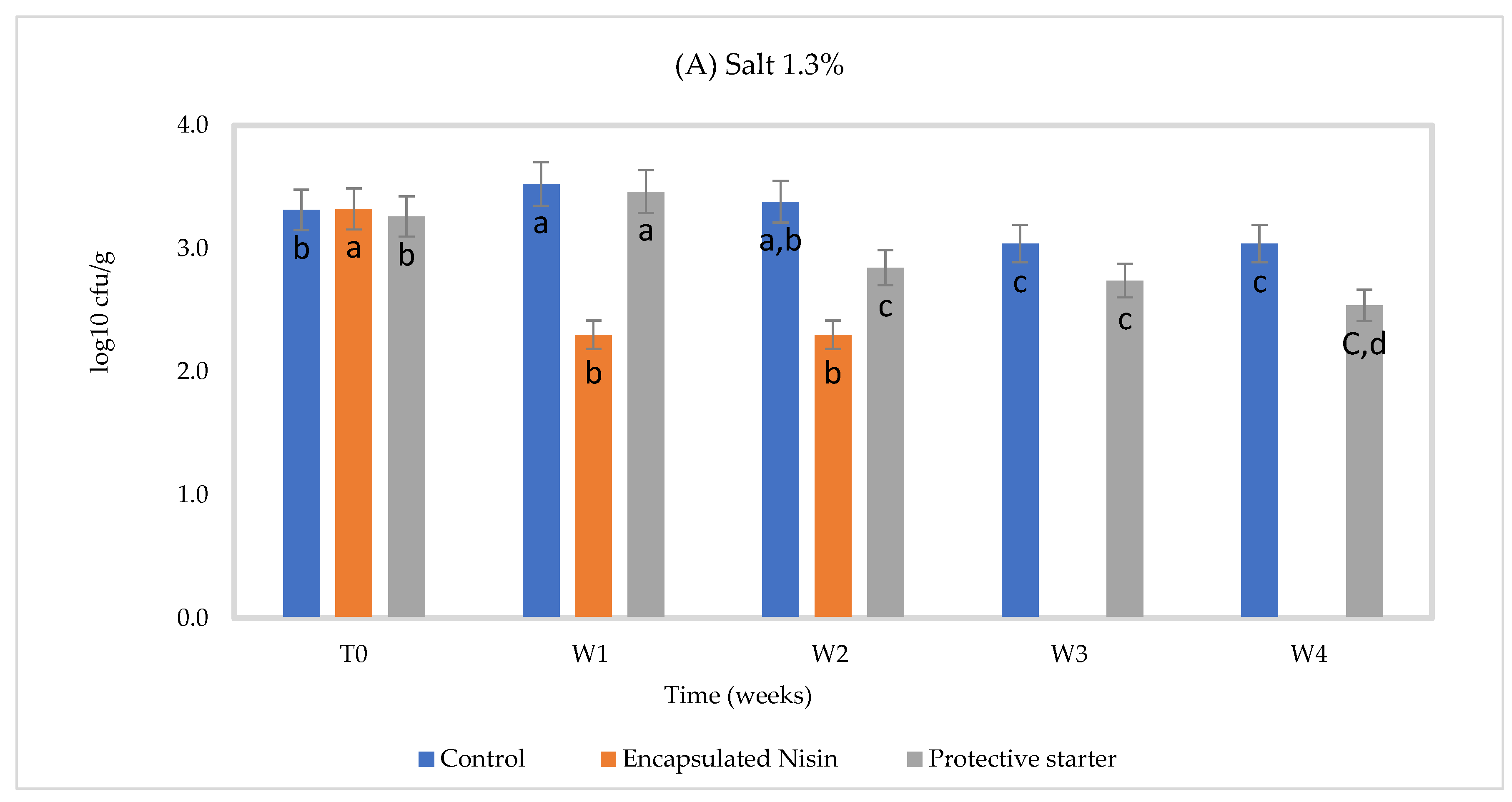
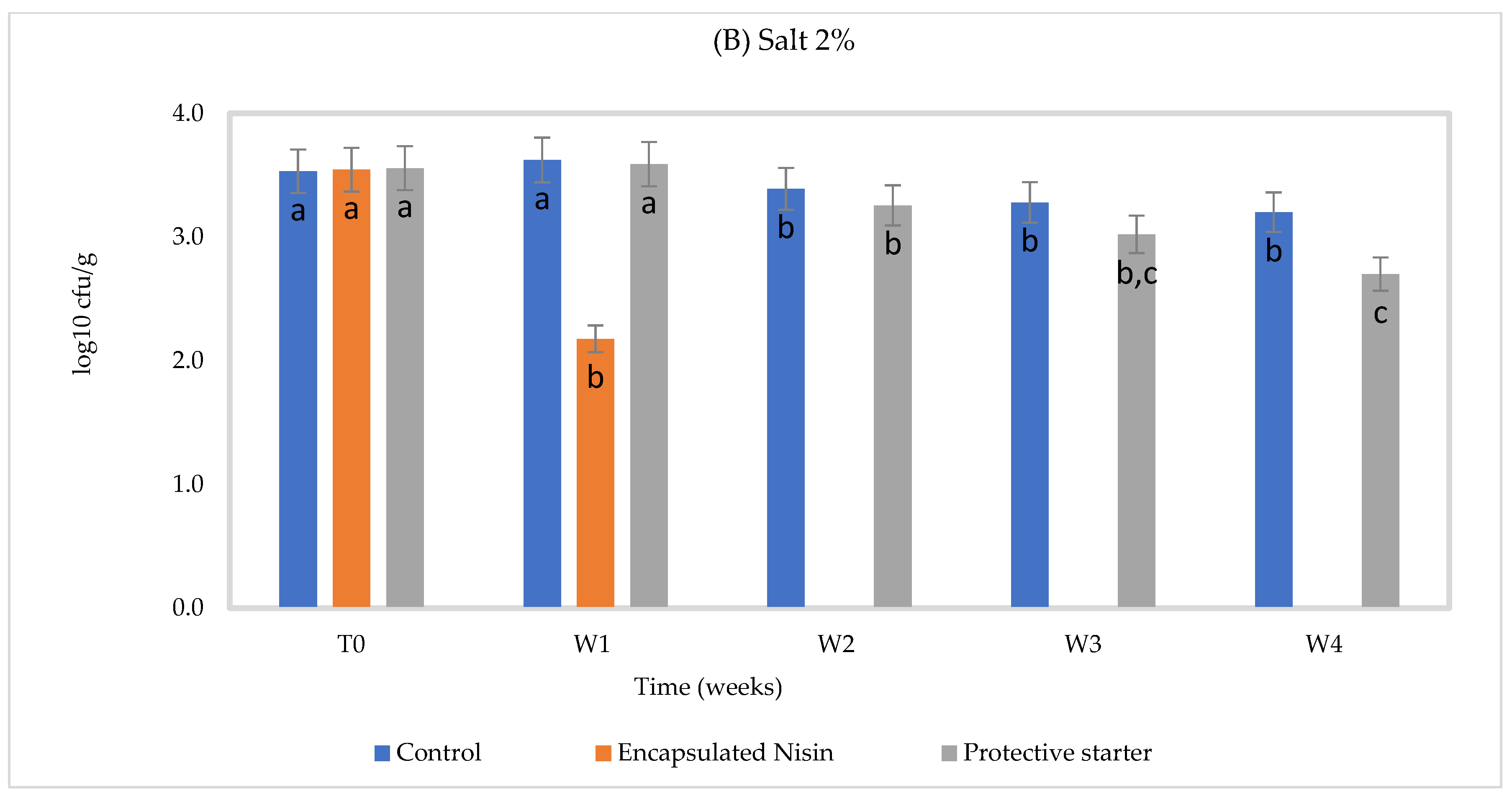
| Treatment | Log cfu/g at 2% NaCl | ||
| Time Zero | Week 1 | Week 2 | |
| Control | 3.53 ± 0.02 b | 3.70 ±0.05 a | 3.57 ± 0.05 b |
| Encapsulated nisin | 3.54 ± 0.04 a | 2.48 ± 0.01 b | 2.30 ± 0.02 c |
| protective starter | 3.56 ± 0.02 a | 3.51 ± 0.05 a | 3.10 ± 0.01 b |
| Treatment | Log cfu/g at 1.3% NaCl | ||
| Time Zero | Week 1 | Week 2 | |
| Control | 3.32 ± 0.02 a | 3.34 ± 0.02 a | Uncountable (high number) |
| Encapsulated nisin | 3.32 ± 0.02 b | 2.54 ± 0.01 c | 3.81 ± 0.04 a |
| protective starter | 3.26 ± 0.03 b | 3.23 ± 0.03 b | 3.80 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, H.; St-Gelais, D.; Gomaa, A.; Fliss, I. Impact of Nisin and Nisin-Producing Lactococcus lactis ssp. lactis on Clostridium tyrobutyricum and Bacterial Ecosystem of Cheese Matrices. Foods 2021, 10, 898. https://doi.org/10.3390/foods10040898
Hassan H, St-Gelais D, Gomaa A, Fliss I. Impact of Nisin and Nisin-Producing Lactococcus lactis ssp. lactis on Clostridium tyrobutyricum and Bacterial Ecosystem of Cheese Matrices. Foods. 2021; 10(4):898. https://doi.org/10.3390/foods10040898
Chicago/Turabian StyleHassan, Hebatoallah, Daniel St-Gelais, Ahmed Gomaa, and Ismail Fliss. 2021. "Impact of Nisin and Nisin-Producing Lactococcus lactis ssp. lactis on Clostridium tyrobutyricum and Bacterial Ecosystem of Cheese Matrices" Foods 10, no. 4: 898. https://doi.org/10.3390/foods10040898
APA StyleHassan, H., St-Gelais, D., Gomaa, A., & Fliss, I. (2021). Impact of Nisin and Nisin-Producing Lactococcus lactis ssp. lactis on Clostridium tyrobutyricum and Bacterial Ecosystem of Cheese Matrices. Foods, 10(4), 898. https://doi.org/10.3390/foods10040898