
The Egg Yolk Content in ω-3 and Conjugated Fatty Acids Can Be Sustainably Increased upon Long-Term Feeding of Laying Hens with a Diet Containing Flaxseeds and Pomegranate Seed Oil

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Birds, Housing and Diets
2.3. Sample Collection and Measurements
2.4. Lipid Extraction and Fatty Acid Analysis
2.5. Identification and Quantification of Fatty Acids by GC–FID and –MS
2.6. Statistical Analysis
3. Results
3.1. Dietary Fats and Hens’ Performance
3.2. Fatty Acid Composition of Egg Yolks
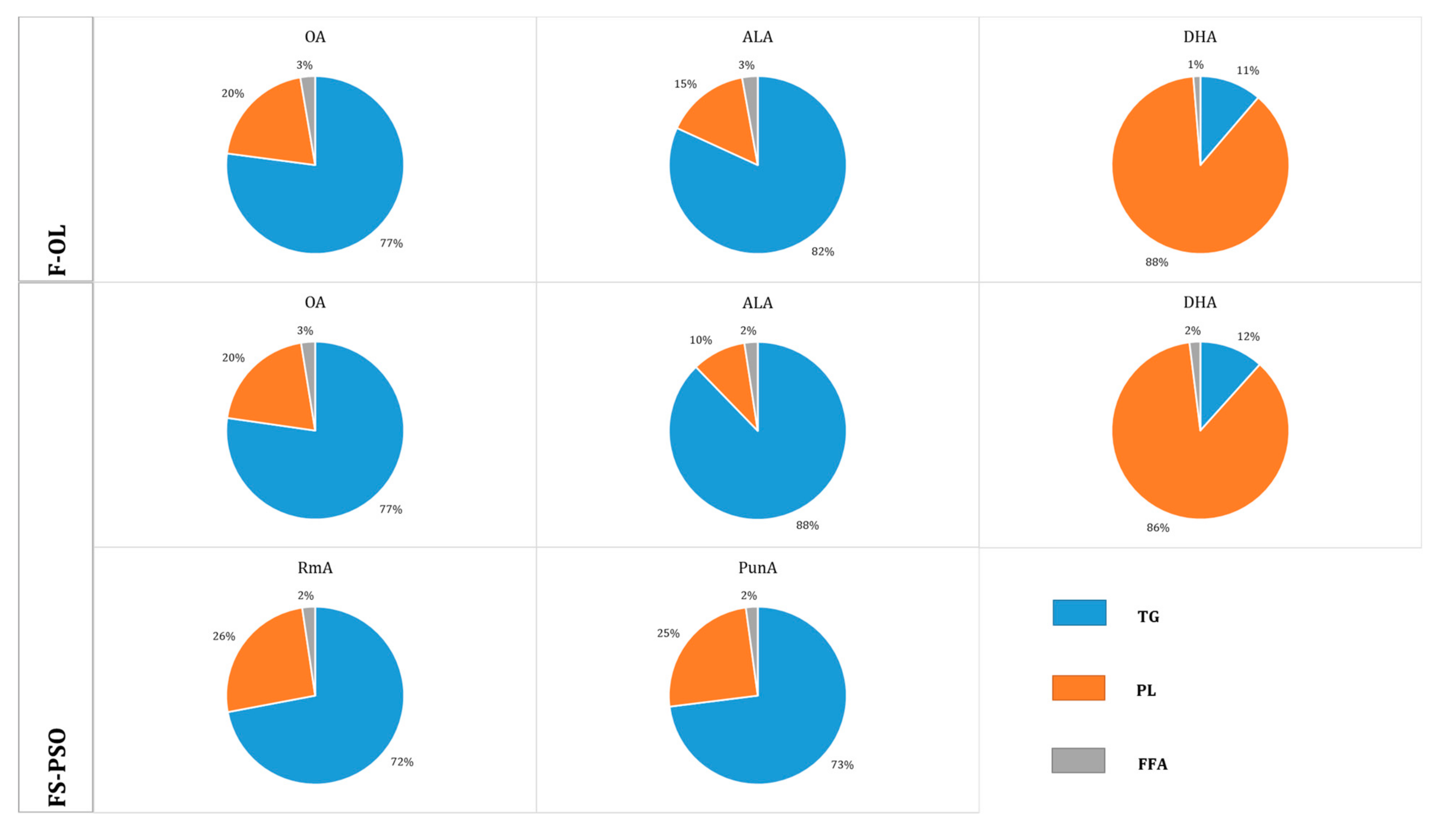
3.3. Distribution of OA, ALA, RmA and PunA of Yolk into Lipid Classes
3.4. Incorporation of Dietary Lipids on Tissues of Hens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roberfroid, M. 1-Defining functional foods and associated claims. In Functional Foods, 2nd ed.; Saarela, M., Ed.; Woodhead Publishing limited: Sawston, Cambridge, UK, 2011; pp. 3–24. [Google Scholar] [CrossRef]
- Siró, I.; Kápolna, E.; Kápolna, B.; Lugasi, A. Functional food. Product development, marketing and consumer acceptance A review. Appetite 2008, 51, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Surai, P.F.; Sparks, N.H.C. Designer eggs: From improvement of egg composition to functional food. Trends Food Sci. Technol. 2001, 12, 7–16. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reifen, R.; Karlinsky, A.; Stark, A.H.; Berkovich, Z.; Nyska, A. α-Linolenic acid (ALA) is an anti-inflammatory agent in inflammatory bowel disease. J. Nutr. Biochem. 2015, 26, 1632–1640. [Google Scholar] [CrossRef] [PubMed]
- Avallone, R.; Vitale, G.; Bertolotti, M. Omega-3 Fatty Acids and Neurodegenerative Diseases: New Evidence in Clinical Trials. Int. J. Mol. Sci 2019, 20, 4256. [Google Scholar] [CrossRef] [Green Version]
- Bowen, K.J.; Harris, W.S.; Kris-Etherton, P.M. Omega-3 Fatty Acids and Cardiovascular Disease: Are There Benefits? Curr. Treat. Options Cardiovasc. Med. 2016, 18, 69. [Google Scholar] [CrossRef] [Green Version]
- Harris, W.S.; Tintle, N.L.; Imamura, F.; Qian, F.; Korat, A.V.A.; Marklund, M.; Djoussé, L.; Bassett, J.K.; Carmichael, P.-H.; Chen, Y.-Y.; et al. Blood n-3 fatty acid levels and total and cause-specific mortality from 17 prospective studies. Nat. Commun. 2021, 12, 2329. [Google Scholar] [CrossRef]
- Brenna, J.T.; Carlson, S.E. Docosahexaenoic acid and human brain development: Evidence that a dietary supply is needed for optimal development. J. Hum. Evol. 2014, 77, 99–106. [Google Scholar] [CrossRef]
- Moloney, F.; Yeow, T.P.; Mullen, A.; Nolan, J.J.; Roche, H.M. Conjugated linoleic acid supplementation, insulin sensitivity, and lipoprotein metabolism in patients with type 2 diabetes mellitus. Am. J. Clin. Nutr. 2004, 80, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Nekooeian, A.A.; Eftekhari, M.H.; Adibi, S.; Rajaeifard, A. Effects of pomegranate seed oil on insulin release in rats with type 2 diabetes. Iran. J. Med. Sci. 2014, 39, 130–135. [Google Scholar]
- Riserus, U.; Vessby, B.; Arnlov, J.; Basu, S. Effects of cis-9,trans-11 conjugated linoleic acid supplementation on insulin sensitivity, lipid peroxidation, and proinflammatory markers in obese men. Am. J. Clin. Nutr. 2004, 80, 279–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sailas, B.; Friedrich, S. Conjugated linoleic acids as functional food: An insight into their health benefits. Nutr. Metab. 2009, 6. [Google Scholar] [CrossRef] [Green Version]
- Dhar Dubey, K.K.; Sharma, G.; Kumar, A. Conjugated linolenic acids: Implication in cancer. J. Agric. Food Chem. 2019, 67, 6091–6101. [Google Scholar] [CrossRef]
- Chin, S.F.; Liu, W.; Storkson, J.M.; Ha, Y.L.; Pariza, M.W. Dietary sources of conjugated dienoic isomers of linoleic acid, a newly recognized class of anticarcinogens. J. Food Compos. Anal. 1992, 5, 185–197. [Google Scholar] [CrossRef]
- Hennessy, A.A.; Ross, R.P.; Devery, R.; Stanton, C. The health promoting properties of the conjugated isomers of α-linolenic acid. Lipids 2011, 46, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.C.; Mignolet, E.; Schneider, Y.J.; Larondelle, Y. Uptake of conjugated linolenic acids and conversion to cis-9, trans-11-or trans-9, trans-11-conjugated linoleic acids in Caco-2 cells. Br. J. Nutr. 2013, 109, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.; Sinclair, A.J.; Xu, C.; Li, D. Incorporation and metabolism of punicic acid in healthy young humans. Mol. Nutr. Food Res. 2009, 53, 1336–1342. [Google Scholar] [CrossRef]
- Tsuzuki, T.; Tokuyama, Y.; Igarashi, M.; Nakagawa, K.; Ohsaki, Y.; Komai, M.; Miyazawa, T. α-Eleostearic acid (9Z11E13E-18:3) is quickly converted to conjugated linoleic acid (9Z11E-18:2) in rats. J. Nutr. 2004, 134, 2634–2639. [Google Scholar] [CrossRef] [Green Version]
- Ngo Njembe, M.T.; Dormal, E.; Gardin, C.; Mignolet, E.; Debier, C.; Larondelle, Y. Effect of the dietary combination of flaxseed and Ricinodendron heudelotii or Punica granatum seed oil on the fatty acid profile of eggs. Food Chem. 2021, 344, 128668. [Google Scholar] [CrossRef]
- de Melo, I.L.P.; de Oliveira e Silva, A.M.; de Carvalho, E.B.T.; Yoshime, L.T.; Sattler, J.A.G.; Mancini-Filho, J. Incorporation and effects of punicic acid on muscle and adipose tissues of rats. Lipids Health Dis. 2016, 15, 40. [Google Scholar] [CrossRef] [Green Version]
- Arao, K.; Wang, Y.-M.; Inoue, N.; Hirata, J.; Cha, J.-Y.; Nagao, K.; Yanagita, T. Dietary effect of pomegranate seed oil rich in 9cis, 11trans, 13cis conjugated linolenic acid on lipid metabolism in obese, hyperlipidemic OLETF rats. Lipids Health Dis. 2004, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.-F.; Sinclair, A.J.; Sun, H.-Y.; Li, D. Fatty acid composition in tissues of mice fed diets containing conjugated linolenic acid and conjugated linoleic acid. J. Food Lipids 2009, 16, 148–163. [Google Scholar] [CrossRef]
- Ngo Njembe, M.T.; Pachikian, B.; Lobysheva, I.; Van Overstraeten, N.; Dejonghe, L.; Verstraelen, E.; Buchet, M.; Rasse, C.; Gardin, C.; Mignolet, E.; et al. A Three-Month Consumption of Eggs Enriched with ω-3, ω-5 and ω-7 Polyunsaturated Fatty Acids Significantly Decreases the Waist Circumference of Subjects at Risk of Developing Metabolic Syndrome: A Double-Blind Randomized Controlled Trial. Nutrients 2021, 13, 663. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Johnston, S.A.; Gous, R.M. Modelling the changes in the proportions of the egg components during a laying cycle. Br. Poult. Sci. 2007, 48, 347–353. [Google Scholar] [CrossRef]
- Bean, L.D.; Leeson, S. Long-term effects of feeding flaxseed on performance and egg fatty acid composition of brown and white hens. Poult. Sci. 2003, 82, 388–394. [Google Scholar] [CrossRef]
- Kostogrys, R.B.; Filipiak-Florkiewicz, A.; Deren, K.; Drahun, A.; Czyzynska-Cichon, I.; Cieslik, E.; Szymczyk, B.; Franczyk-Zarow, M. Effect of dietary pomegranate seed oil on laying hen performance and physicochemical properties of eggs. Food Chem. 2017, 221, 1096–1103. [Google Scholar] [CrossRef]
- Tsuzuki, T.; Kawakami, Y.; Abe, R.; Nakagawa, K.; Koba, K.; Imamura, J.; Iwata, T.; Ikeda, I.; Miyazawa, T. Conjugated linolenic acid is slowly absorbed in rat intestine, but quickly converted to conjugated linoleic acid. J. Nutr. 2006, 136, 2153–2159. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, R.; Yasui, Y.; Kohno, H.; Miyamoto, S.; Hosokawa, M.; Miyashita, K.; Tanaka, T. Catalpa seed oil rich in 9t,11t,13c-conjugated linolenic acid suppresses the development of colonic aberrant crypt foci induced by azoxymethane in rats. Oncol. Rep. 2006, 16, 989–996. [Google Scholar] [CrossRef]
- Kinsella, J.E. Alpha-linolenic acid: Functions and effects on linoleic acid metabolism and eicosanoid-mediated reactions. Adv. Food Nutr. Res. 1991, 35, 1–184. [Google Scholar] [CrossRef] [PubMed]
- Banni, S. Conjugated linoleic acid metabolism. Curr. Opin. Lipidol. 2002, 13, 261–266. [Google Scholar] [CrossRef]
- Choi, Y.; Park, Y.; Storkson, J.M.; Pariza, M.W.; Ntambi, J.M. Inhibition of stearoyl-CoA desaturase activity by the cis-9, trans-11 isomer and the trans-10, cis-12 isomer of conjugated linoleic acid in MDA-MB-231 and MCF-7 human breast cancer cells. Biochem. Biophys. Res. Commun. 2002, 294, 785–790. [Google Scholar] [CrossRef]
- Haug, A.; Nyquist, N.F.; Thomassen, M.; Høstmark, A.T.; Ostbye, T.K. N-3 fatty acid intake altered fat content and fatty acid distribution in chicken breast muscle, but did not influence mRNA expression of lipid-related enzymes. Lipids Health Dis. 2014, 13, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speake, B.K.; Murray, A.M.; Noble, R.C. Transport and transformations of yolk lipids during development of the avian embryo. Prog. Lipid Res. 1998, 37, 1–32. [Google Scholar] [CrossRef]
- Banni, S.; Carta, G.; Angioni, E.; Murru, E.; Scanu, P.; Melis, M.P.; Bauman, D.E.; Fischer, S.M.; Ip, C. Distribution of conjugated linoleic acid and metabolites in different lipid fractions in the rat liver. J. Lipid Res. 2001, 42, 1056–1061. [Google Scholar] [CrossRef]
- Belury, M.A.; Kempa-Steczko, A. Conjugated linoleic acid modulates hepatic lipid composition in mice. Lipids 1997, 32, 199–204. [Google Scholar] [CrossRef]


{kind=link}
| Fatty Acids | Fat Sources | Dietary Treatments | |||
|---|---|---|---|---|---|
| Olive Oil | Flaxseed Oil | Pomegranate Seed Oil | F-OL | FS-PSO | |
| % wt. of total fatty acids identified | mg/g of feed | ||||
| Palmitic acid (C16:0) | 9.68 | 5.27 | 2.61 | 19.36 | 9.08 |
| Stearic acid (C18:0) | 3.18 | 4.20 | 1.74 | 4.74 | 2.96 |
| Oleic acid (C18:1c9) | 78.46 | 16.66 | 4.39 | 91.06 | 14.74 |
| Cis-vaccenic acid (C18:1c11) | 1.61 | 0.67 | 0.40 | 2.16 | 0.67 |
| Linoleic acid (C18:2c9,c12) | 4.82 | 15.22 | 5.67 | 23.63 | 21.16 |
| Alpha-linolenic acid (C18:3c9,c12,c15) | 0.71 | 56.80 | 0.73 | 2.09 | 16.57 |
| Punicic acid (C18:3c9,t11,c13) | ND | ND | 73.21 | ND | 39.53 |
| Alpha-eleostearic acid (C18:3c9,t11,t13) * | ND | ND | 0.64 | ND | 0.35 |
| Alpha-calendic acid (C18:3t8,t10,c12) * | ND | ND | 0.82 | ND | 0.44 |
| Catalpic acid (C18:3t9,t11,c13) * | ND | ND | 4.85 | ND | 2.62 |
| Beta-eleostearic acid (C18:3t9,t11,t13) * | ND | ND | 1.99 | ND | 1.08 |
| Beta-calendic acid (C18:3t8,t10,t12) * | ND | ND | 0.56 | ND | 0.30 |
| Σ SFA | 13.56 | 9.99 | 5.70 | 25.74 | 13.53 |
| Σ MUFA | 80.76 | 17.44 | 5.51 | 94.26 | 15.52 |
| Σ n-6 PUFA | 4.82 | 15.27 | 5.70 | 23.63 | 21.26 |
| Σ n-3 PUFA | 0.84 | 57.26 | 1.02 | 2.25 | 16.69 |
| Σ CLnA | ND | ND | 82.07 | ND | 44.36 |
| Diet Effect | Diet × Period Effect | p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Dietary Treatments | SE | Period 1 | Period 2 | Period 3 | Period 4 | Period 5 | Period 6 | Diet | Period | Diet × Period | ||
| Laying rate (%) | F-OL | 85.75 | 1.61 | 76.97 | 86.81 | 89.65 | 89.85 | 85.69 | 85.51 | 0.75 | 0.12 | 0.93 |
| FS-PSO | 85.01 | 79.19 | 82.89 | 92.17 | 85.37 | 85.44 | 84.98 | |||||
| Egg weight (g) | F-OL | 62.86 | 0.47 | 63.41 | 65.54 | 60.92 | 60.97 | 62.63 | 63.64 | 0.06 | 0.06 | 0.11 |
| FS-PSO | 61.60 | 60.11 | 61.04 | 62.34 | 60.14 | 62.44 | 63.53 | |||||
| Yolk weight (g) | F-OL | 16.57 | 0.13 | 16.71 | 17.28 | 16.06 | 16.07 | 16.51 | 16.78 | 0.07 | 0.05 | 0.12 |
| FS-PSO | 16.24 | 15.84 | 16.09 | 16.43 | 15.85 | 16.46 | 16.75 | |||||
| Diet Effect | Diet × Period Effect | p-Value | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acids (In mg/g of Yolk) | Dietary Treatments | SE | Period 1 | Period 2 | Period 3 | Period 4 | Period 5 | Period 6 | Diet | Period | Diet × Period | |
| Palmitic acid (C16:0) | F-OL | 59.15 a | 0.77 | 75.85 a | 60.04 b,c,d | 53.50 d | 56.98 d | 56.57 d | 51.93 d | 0.0031 | <0.0001 | 0.0858 |
| FS-PSO | 62.34 b | 74.49 a | 65.56 b | 59.44 b,c,d | 64.34 b,c | 57.34 c,d | 52.88 d | |||||
| Hypogeic acid (C16:1c7) | F-OL | 4.98 a | 0.09 | 5.14 a,b | 4.57 b | 4.72 a,b | 5.33 a,b | 5.55 a | 4.58 b | <0.0001 | 0.0045 | 0.0186 |
| FS-PSO | 2.38 b | 3.04 c | 2.10 c | 2.32 c | 2.27 c | 2.24 c | 2.32 c | |||||
| Palmitoleic acid (C16:1c9) | F-OL | 3.72 a | 0.11 | 6.90 a | 3.54 b | 2.94 b,c | 3.09 b,c | 3.24 b,c | 2.57 b,c | 0.0667 | <0.0001 | 0.1002 |
| FS-PSO | 3.43 a | 5.96 a,b | 3.73 b | 2.82 b,c | 3.40 b | 2.57 b,c | 2.08 c | |||||
| Cis-7, trans-9-hexadecadienoic acid (C16:2c7,t9) | F-OL | 0.07 a | 0.07 | 0.12 c | 0.06 c | 0.06 c | 0.07 c | 0.06 c | Traces c | <0.0001 | 0.0173 | 0.0059 |
| FS-PSO | 3.75 b | 2.82 b | 3.64 a,b | 3.88 a | 3.77 a | 4.08 a | 4.31 a | |||||
| Stearic acid (C18:0) | F-OL | 20.27 a | 0.31 | 24.32 a | 19.89 b,c | 18.84 c | 20.36 b,c | 19.62 c | 18.62 c | <0.0001 | <0.0001 | 0.2497 |
| FS-PSO | 23.49 b | 25.26 a | 24.63 a | 22.49 a,b,c | 24.22 a | 23.01 a,b | 21.32 a,b,c | |||||
| Oleic acid (C18:1c9) | F-OL | 153.40 a | 1.70 | 154.86 a | 160.25 a | 144.25 a | 158.13 a | 156.14 a | 146.78 a | <0.0001 | <0.0001 | <0.0001 |
| FS-PSO | 72.59 b | 103.87 b | 70.04 c | 67.19 c | 71.52 c | 63.30 c | 59.68 c | |||||
| Cis-vaccenic acid (C18:1c11) | F-OL | 4.30 a | 0.05 | 5.78 a | 4.25 b | 4.00 b,c | 4.16 b,c | 4.03 b,c | 3.56 c | <0.0001 | <0.0001 | 0.3349 |
| FS-PSO | 2.45 b | 4.23 b,c | 2.34 d | 2.28 d | 2.29 d | 1.86 d | 1.68 d | |||||
| Linoleic acid (C18:2c9,c12) | F-OL | 33.11 a | 0.63 | 45.48 a,b | 30.17 d,e | 29.70 d,e | 32.67 c,d,e | 31.75 d,e | 28.86 e | <0.0001 | <0.0001 | 0.4329 |
| FS-PSO | 39.46 b | 49.45 a | 36.65 c,d | 39.63 b,c | 37.98 c | 36.15 c,d,e | 36.89 c,d,e | |||||
| Rumenic acid (C18:2c9,t11) | F-OL | 0.13 a | 0.47 | 0.28 c | 0.16 c | 0.08 c | 0.08 c | 0.15 c | 0.06 c | <0.0001 | 0.0026 | 0.0016 |
| FS-PSO | 35.93 b | 28.64 b | 35.58 a | 35.25 a,b | 37.70 a | 38.23 a | 40.19 a | |||||
| Trans-9, trans-11-octadecadienoic acid (C18:2t9,t11) * | F-OL | ND a | 0.02 | ND a | ND a | ND a | ND a | ND a | ND a | <0.0001 | 0.6133 | 0.4942 |
| FS-PSO | 0.96 b | 1.05 b | 1.01 b | 0.92 b | 0.92 b | 0.92 b | 0.92 b | |||||
| Cis-6, cis-9, trans-11-octadecatrienoic acid (C18:3c6,c9,t11) | F-OL | 0.05 a | 0.02 | 0.07 b | 0.06 b | ND b | 0.07 b | 0.06 b | 0.05 b | <0.0001 | 0.7764 | 0.7149 |
| FS-PSO | 0.88 b | 0.94 a | 0.89 a | 0.94 a | 0.82 a | 0.84 a | 0.84 a | |||||
| Alpha-linolenic acid (C18:3c9,c12,c15) | F-OL | 0.91 a | 0.19 | 1.50 c | 0.86 c | 0.74 c | 0.84 c | 0.85 c | 0.67 c | <0.0001 | 0.4431 | 0.0327 |
| FS-PSO | 6.15 b | 4.76 b | 6.85 a,b | 5.61 a,b | 6.03 a,b | 6.35 a,b | 7.29 a | |||||
| Punicic acid (C18:3c9,t11,c13) | F-OL | ND a | 0.40 | ND c | ND c | ND c | ND c | ND c | ND c | <0.0001 | 0.0007 | 0.0007 |
| FS-PSO | 20.00 b | 21.47 a,b | 19.14 b | 16.93 b | 18.05 b | 24.16 a | 20.23 a,b | |||||
| Alpha-eleostearic acid (C18:3c9,t11,t13) ** | F-OL | ND a | 0.004 | ND d | ND d | ND d | ND d | ND d | ND d | <0.0001 | 0.0023 | 0.0023 |
| FS-PSO | 0.25 b | 0.30 a | 0.26 a,b | 0.26 a,b | 0.21 c | 0.25 a,b,c | 0.23 b,c | |||||
| Alpha-calendic acid (C18:3t8,t10,c12) ** | F-OL | ND a | 0.004 | ND d | ND d | ND d | ND d | ND d | ND d | <0.0001 | <0.0001 | <0.0001 |
| FS-PSO | 0.20 b | 0.25 a,b | 0.26 a | 0.20 b | 0.23 a,b | 0.14 c | 0.12 c | |||||
| Catalpic acid (C18:3t9,t11,c13) ** | F-OL | ND a | 0.02 | ND c | ND c | ND c | ND c | ND c | ND c | <0.0001 | 0.1392 | 0.1426 |
| FS-PSO | 1.06 b | 1.18 a,b | 1.05 a,b | 1.16 a | 0.91 b | 1.07 a,b | 1.02 a,b | |||||
| Beta-eleostearic acid (C18:3t9,t11,t13) ** | F-OL | ND a | 0.01 | ND c | ND c | ND c | ND c | ND c | ND c | <0.0001 | 0.0597 | 0.0600 |
| FS-PSO | 0.69 b | 0.75 a,b | 0.68 a,b | 0.69 a,b | 0.58 b | 0.74 a | 0.70 a,b | |||||
| Beta-calendic acid (C18:3t8,t10,t12) ** | F-OL | ND a | 0.01 | ND c | ND c | ND c | ND c | ND c | ND c | <0.0001 | 0.1362 | 0.1362 |
| FS-PSO | 0.33 b | 0.38 a | 0.34 a,b | 0.34 a,b | 0.29 b | 0.34 a,b | 0.32 a,b | |||||
| Arachidonic acid (C20:4c5,c8,c11,c14) | F-OL | 5.66 a | 0.08 | 5.91 a,b | 5.42 a,b | 5.34 a,b,c | 5.86 a,b | 6.01 a | 5.40 a,b,c | <0.0001 | <0.0001 | <0.0001 |
| FS-PSO | 3.95 b | 5.05 b,c | 3.78 d,e | 4.45 c,d | 3.82 d,e | 3.61 d,e | 3.00e | |||||
| Eicosapentaenoic acid (C20:5c5,c8,c11,c14,c17) | F-OL | ND a | 0.005 | ND c | ND c | ND c | ND c | ND c | ND c | <0.0001 | 0.1236 | 0.1289 |
| FS-PSO | 0.16 b | 0.19 a | 0.18 a,b | 0.12 b | 0.16 a,b | 0.16 a,b | 0.15 a,b | |||||
| n-6 Docosapentaenoic acid (C22:5c4,c7,c10,c13,c16) | F-OL | 1.36 a | 0.03 | 1.47 a,b | 1.18 b | 1.52 a,b | 1.51 a | 1.33 a,b | 1.16 b,c | <0.0001 | <0.0001 | 0.0012 |
| FS-PSO | 0.35 b | 0.80 c | 0.31 d | 0.34 d | 0.29 d | 0.20 d | 0.16 d | |||||
| n-3 Docosapentaenoic acid (C22:5c7,c10,c13,c16,c19) | F-OL | 0.48 a | 0.40 | 0.44 d | 0.41 d | 0.49 c,d | 0.56 c,d | 0.53 c,d | 0.41 d | <0.0001 | <0.0001 | 0.0002 |
| FS-PSO | 1.39 b | 0.92 b,c | 1.72 a | 1.25 a,b | 1.44 a | 1.66 a | 1.35 a,b | |||||
| Docosahexaenoic acid (C22:6c4,c7c10,c13,c16,c19) | F-OL | 2.22 a | 0.10 | 2.57 c | 2.22 c | 2.27 c | 2.28 c | 2.08 c | 1.95 c | <0.0001 | 0.0898 | 0.0020 |
| FS-PSO | 4.90 b | 3.82 b | 5.06 a | 5.54 a | 5.14 a | 4.80 a,b | 5.03 a,b | |||||
| Σ SFA | F-OL | 81.79 a | 1.02 | 103.21 a | 82.50 d,e | 74.47e | 79.72e | 78.32e | 72.51e | <0.0001 | <0.0001 | 0.0934 |
| FS-PSO | 88.87 b | 102.89 a,b | 93.48 a,b,c | 84.98 c,d,e | 91.75 b,c,d | 83.23 c,d,e | 76.86e | |||||
| Σ MUFA | F-OL | 166.99 a | 1.87 | 171.46 a | 173.58 a | 156.68 a | 171.65 a | 170.07 a | 158.49 a | <0.0001 | <0.0001 | 0.0001 |
| FS-PSO | 80.95 b | 116.79 b | 79.01 c | 69.58 c | 80.40 c | 70.81 c | 69.12 c | |||||
| Σ n-6 PUFA | F-OL | 42.03 a | 0.71 | 56.63 a | 38.18 b,c | 38.18 b,c | 41.78 b,c | 40.67 b,c | 36.76 c | 0.0015 | <0.0001 | 0.2487 |
| FS-PSO | 45.28 b | 56.43 a | 42.37 b,c | 46.23 b | 43.80 b,c | 41.45 b,c | 41.38 b,c | |||||
| Σ n-3 PUFA | F-OL | 3.67 a | 0.30 | 4.53 a | 3.55 a | 3.57 a | 3.74 a | 3.54 a | 3.08 a | <0.0001 | 0.6020 | 0.5341 |
| FS-PSO | 14.14 b | 14.55 b | 14.89 b | 12.73 b | 13.87 b | 14.00 b | 14.81 b | |||||
| Σ CLA | F-OL | 0.20 a | 0.46 | 0.38 c | 0.22 c | 0.13 c | 0.15 c | 0.21 c | 0.13 c | <0.0001 | 0.0016 | 0.0007 |
| FS-PSO | 37.04 b | 29.87 b | 36.79 a | 36.20 a | 38.81 a | 40.40 a | 41.29 a | |||||
| Σ CLnA | F-OL | 0.05 a | 0.39 | 0.07 c | 0.06 c | ND c | 0.07 c | 0.06 c | 0.05 c | <0.0001 | 0.0001 | 0.0002 |
| FS-PSO | 23.34 b | 25.28 a,b | 22.62 b | 20.30 b | 20.85 b | 27.53 a | 23.47 a,b | |||||
| Triglycerides | Phospholipids | |||
|---|---|---|---|---|
| % wt. of total fatty acids identified | F-OL | FS-PS | F-OL | FS-PS |
| Palmitic acid (C16:0) | 17.91 ± 1.93 a | 19.92 ± 1.32 b | 25.07 ± 0.35 a | 23.40 ± 0.49 b |
| Hypogeic acid (C16:1c7) | 2.38 ± 0.12 a | 0.96 ± 0.09 b | 0.56 ± 0.03 a | 0.17 ± 0.01 b |
| Palmitoleic acid (C16:1c9) | 1.12 ± 0.08 a | 1.09 ± 0.07 a | 0.41 ± 0.02 a | 0.27 ± 0.01 b |
| Cis-7, trans-9-hexadecadienoic acid (C16:2cis7,t9) | 0.01 ± 0.004 a | 1.87 ± 0.27 b | 0.02 ± 0.002 a | 0.20 ± 0.01 a |
| Stearic acid (C18:0) | 4.22 ± 0.20 a | 5.60 ± 0.21 b | 17.83 ± 0.50 a | 19.39 ± 0.45 b |
| Oleic acid (C18:1c9) | 30.45 ± 9.19 a | 17.90 ± 3.84 b | 28.34 ± 0.55 a | 12.58 ± 0.36 b |
| Cis-vaccenic acid (C18:1c11) | 30.08 ± 8.64 a | 8.85 ± 3.47 b | 1.12 ± 0.04 a | 0.38 ± 0.01 b |
| Linoleic acid (C18:2c9,c12) | 11.02 ± 0.34 a | 13.62 ± 0.87 a | 12.39 ± 0.31 a | 12.04 ± 0.26 a |
| Rumenic acid (C18:2c9,t11) | 0.06 ± 0.01 a | 14.62 ± 0.41 b | 0.04 ± 0.01 a | 9.64 ± 0.32 b |
| Trans-9, trans-11-octadecadienoic acid (C18:2t9,t11) * | ND a | 0.01 ± 0.005 b | ND a | ND a |
| Cis-6, cis-9, trans-11-octadecatrienoic acid (C18:3c6,c9,t11) | 0.01 ± 0.004 a | 0.24 ± 0.010 b | 0.01 ± 0.002 a | 0.56 ± 0.03 b |
| Alpha-linolenic acid (C18:3c9,c12,c15) | 0.30 ± 0.01 a | 2.89 ± 0.24 b | 0.11 ± 0.004 a | 0.60 ± 0.04 b |
| Punicic acid (C18:3c9,t11,c13) | ND a | 7.90 ± 1.15 b | ND a | 4.98 ± 0.3 b |
| Alpha-eleostearic acid (C18:3c9,t11,t13) ** | ND a | 0.08 ± 0.004 b | ND a | 0.05 ± 0.002 b |
| Alpha-calendic acid (C18:3t8,t10,c12) ** | ND a | 0.06 ± 0.01 b | ND a | 0.05 ± 0.004 b |
| Catalpic acid (C18:3t9,t11,c13) ** | ND a | 0.35 ± 0.07 b | ND a | 0.28 ± 0.01 b |
| Beta-eleostearic acid (C18:3t9,t11,t13) ** | ND a | 0.24 ± 0.02 b | ND a | 0.19 ± 0.01 b |
| Beta-calendic acid (C18:3t8,t10,t12) ** | ND a | 0.11 ± 0.01 b | ND a | 0.07 ± 0.004 b |
| Arachidonic acid (C20:4c5,c8,c11,c14) | 0.72 ± 0.10 a | 0.41 ± 0.06 b | 6.48 ± 0.28 a | 4.51 ± 0.14 b |
| Eicosapentaenoic acid (C20:5c5,c8,c11,c14,c17) | ND a | 0.05 ± 0.01 b | ND a | 0.16 ± 0.01 b |
| n-6 Docosapentaenoic acid (C22:5c4,c7,c10,c13,c16) | 0.13 ± 0.03 a | 0.03 ± 0.01 b | 1.40 ± 0.19 a | 0.26 ± 0.03 b |
| n-3 Docosapentaenoic acid (C22:5c7,c10,c13,c16,c19) | 0.12 ± 0.008 a | 0.39 ± 0.03 b | 0.43 ± 0.01 a | 1.41 ± 0.10 b |
| Docosahexaenoic acid (C22:6c4,c7c10,c13,c16,c19) | 0.20 ± 0.03 a | 0.48 ± 0.07 b | 2.83 ± 0.13 a | 6.55 ± 0.38 b |
| Σ SFA | 22.48 ± 0.63 a | 26.20 ± 0.49 b | 44.00 ± 0.86 a | 43.35 ± 0.92 a |
| Σ MUFA | 64.44 ± 0.81 a | 29.01 ± 0.73 b | 30.90 ± 0.57 a | 13.60 ± 0.32 b |
| Σ n-6 PUFA | 12.25 ± 0.43 a | 14.52 ± 0.27 b | 21.44 ± 0.38 a | 17.71 ± 0.35 b |
| Σ n-3 PUFA | 0.64 ± 0.03 a | 3.91 ± 0.23 b | 3.41 ± 0.12 a | 8.83 ± 0.43 b |
| Σ CLA | 0.07 ± 0.01 a | 14.67 ± 0.43 b | 0.05 ± 0.01 a | 9.66 ± 0.33 b |
| Σ CLnA | 0.01 ± 0.004 a | 8.98 ± 0.33 b | 0.01 ± 0.002 a | 6.18 ± 0.28 b |
| Liver | Heart | Muscle | Adipose Tissue | ||||||
|---|---|---|---|---|---|---|---|---|---|
| % wt. of total fatty acids identified | Lipid fractions | F-OL | FS-PSO | F-OL | FS-PSO | F-OL | FS-PSO | F-OL | FS-PSO |
| Palmitic acid (C16:0) | TG | 16.55 ± 0.77 a | 18.15 ± 1.02 a | 12.23 ± 0.34 a | 14.25 ± 0.34 b | 13.99 ± 0.41 a | 15.59 ± 0.49 a | 11.04 ± 0.29 a | 13.62 ± 0.29 b |
| PL | 22.14 ± 0.15 a | 20.90 ± 0.51 b | 20.48 ± 0.36 a | 19.49 ± 0.46 a | 17.88 ± 0.43 a | 18.09 ± 0.80 a | ND | ND | |
| Hypogeic acid (C16:1c7) | TG | 1.53 ± 0.10 a | 0.70 ± 0.08 b | 0.66 ± 0.04 a | 0.47 ± 0.05 b | 0.94 ± 0.06 a | 0.65 ± 0.04 b | 0.52 ± 0.03 a | 0.39 ± 0.03 b |
| PL | 0.41 ± 0.03 a | 0.16 ± 0.02 b | 0.14 ± 0.005 a | 0.07 ± 0.01 b | 0.14 ± 0.01 a | 0.05 ± 0.01 b | ND | ND | |
| Palmitoleic acid (C16:1c9) | TG | 0.91 ± 0.16 a | 0.83 ± 0.04 a | 0.85 ± 0.10 a | 0.98 ± 0.11 a | 0.89 ± 0.11 a | 1.34 ± 0.22 a | 0.64 ± 0.06 a | 0.80 ± 0.06 a |
| PL | 0.37 ± 0.06 a | 0.28 ± 0.02 a | 0.10 ± 0.01 a | 0.08 ± 0.01 a | 0.12 ± 0.004 a | 0.13 ± 0.01 a | ND | ND | |
| Stearic acid (C18:0) | TG | 5.19 ± 0.30 a | 7.22 ± 0.50 b | 3.21 ± 0.11 a | 4.20 ± 0.25 b | 4.27 ± 0.13 a | 5.92 ± 0.92 a | 3.30 ± 0.13 a | 4.49 ± 0.42 b |
| PL | 18.54 ± 0.55 a | 17.13 ± 0.88 a | 20.76 ± 0.61 a | 20.74 ± 0.75 a | 18.65 ± 0.49 a | 16.89 ± 1.24 a | ND | ND | |
| Oleic acid (C18:1c9) | TG | 60.22 ± 0.65 a | 27.91 ± 1.88 b | 48.28 ± 0.30 a | 33.23 ± 1.18 b | 58.49 ± 0.45 a | 35.14 ± 1.66 b | 63.42 ± 0.24 a | 30.88 ± 2.01 b |
| PL | 26.16 ± 0.68 a | 12.12 ± 0.85 b | 13.65 ± 0.49 a | 5.19 ± 0.57 b | 21.10 ± 0.35 a | 5.84 ± 0.58 b | ND | ND | |
| Cis-vaccenic acid (C18:1c11) | TG | 1.37 ± 0.07 a | 1.03 ± 0.08 b | 14.76 ± 0.09 a | 1.23 ± 0.05 b | 1.84 ± 0.02 a | 1.52 ± 0.11 b | 2.47 ± 0.11 a | 1.69 ± 0.12 b |
| PL | 1.04 ± 0.03 a | 0.44 ± 0.04 b | 2.00 ± 0.16 a | 0.69 ± 0.10 b | 2.07 ± 0.05 a | 0.82 ± 0.08 b | ND | ND | |
| Linoleic acid (C18:2c9,c12) | TG | 11.59 ± 1.23 a | 17.60 ± 0.37 b | 17.77 ± 0.55 a | 23.85 ± 0.67 b | 17.05 ± 0.68 a | 20.05 ± 0.78 b | 16.40 ± 0.33 a | 23.63 ± 0.60 b |
| PL | 11.66 ± 0.36 a | 11.51 ± 0.45 a | 10.61 ± 0.54 a | 13.71 ± 0.53 b | 16.02± 0.75 a | 24.15 ± 0.81 b | ND | ND | |
| Rumenic acid (C18:2c9,t11) | TG | 0.02 ± 0.004 a | 9.88 ± 1.09 b | 0.01 ± 0.001 a | 9.04 ± 0.85 b | 0.02 ± 0.003 a | 8.68 ± 1.05 b | 0.01 ± 0.005 a | 10.07 ± 1.33 b |
| PL | 0.02 ± 0.001 a | 8.34 ± 1.10 b | Traces a | 5.73 ± 0.47 b | 0.02 ± 0.01 a | 5.26 ± 0.48 b | ND | ND | |
| Cis-6, cis-9, trans-11-octadecatrienoic acid (C18:3c6,c9,t11) | TG | ND a | 1.72 ± 0.26 b | ND a | 1.38 ± 0.14 b | ND a | 1.26 ± 0.13 b | ND a | 1.94 ± 0.21 b |
| PL | ND a | 3.80 ± 0.92b | ND a | 2.14 ± 0.44 b | ND a | 2.42 ± 0.23 b | ND | ND | |
| Alpha-linolenic acid (C18:3c9,c12,c15) | TG | 0.39 ± 0.07 a | 3.92 ± 1.08 b | 0.73 ± 0.04 a | 5.93 ± 1.27 b | 0.73 ± 0.05 a | 4.05 ± 0.71 b | 1.09 ± 0.04 a | 5.96 ± 1.32 b |
| PL | 0.09 ± 0.02 a | 0.47 ± 0.09 b | 0.08 ± 0.002 a | 0.29 ± 0.07 b | 0.12 ± 0.005 a | 0.45 ± 0.11 b | ND | ND | |
| Punicic acid (C18:3c9,t11,c13) | TG | ND a | 4.69 ± 0.99 b | ND a | 1.38 ± 0.43 b | ND a | 1.38 ± 0.29 b | ND a | 1.44 ± 0.37 b |
| PL | ND a | 3.18 ± 0.70b | ND a | 0.67 ± 0.10 b | ND a | 0.91 ± 0.28 b | ND | ND | |
| Arachidonic acid (C20:4c5,c8,c11,c14) | TG | 0.36 ± 0.07 a | 0.33 ± 0.05 a | 0.14 ± 0.01 a | 0.12 ± 0.02 a | 0.15 ± 0.03 a | 0.12 ± 0.03 a | 0.08 ± 0.01 a | 0.05 ± 0.01 a |
| PL | 10.85 ± 0.30 a | 7.53 ± 0.82 b | 26.14 ± 1.48 a | 24.08 ± 1.08 a | 13.04 ± 0.89 a | 10.42 ± 1.41 a | ND | ND | |
| Eicosapentaenoic acid (C20:5c5,c8,c11,c14,c17) | TG | Traces a | 0.02 ± 0.01 b | Traces a | 0.01 ± 0.001 a | Traces a | 0.01 ± 0.01 b | ND | ND |
| PL | 0.03 ± 0.002 a | 0.50 ± 0.16 b | 0.05 ± 0.01 a | 0.47 ± 0.16 b | 0.03 ± 0.004 a | 0.29 ± 0.08 b | ND | ND | |
| n-6 Docosapentaenoic acid (C22:5c4,c7,c10,c13,c16) | TG | 0.06 ± 0.01 a | ND b | 0.01 ± 0.001 a | Traces b | 0.01 ± 0.004 a | ND b | ND | ND |
| PL | 1.98 ± 0.17 a | 0.20 ± 0.05 b | 0.91 ± 0.06 a | 0.09 ± 0.02 b | 1.60 ± 0.19 a | 0.16 ± 0.03 b | ND | ND | |
| n-3 Docosapentaenoic acid (C22:5c7,c10,c13,c16,c19) | TG | 0.04 ± 0.01 a | 0.16 ± 0.02 b | 0.01 ± 0.002 a | 0.03 ± 0.005 a | 0.02 ± 0.005 a | 0.04 ± 0.01 a | ND a | 0.01 ± 0.01 b |
| PL | 0.40 ± 0.05 a | 1.40 ± 0.15 b | 0.32 ± 0.04 a | 0.95 ± 0.09 b | 0.86 ± 0.09 a | 1.76 ± 0.16 b | ND | ND | |
| Docosahexaenoic acid (C22:6c4,c7c10,c13,c16,c19) | TG | 0.11 ± 0.02 a | 0.30 ± 0.02 b | 0.02 ± 0.001 a | 0.04 ± 0.004 a | 0.04 ± 0.01 a | 0.08 ± 0.02 a | ND a | 0.01 ± 0.01 b |
| PL | 3.80 ± 0.46 a | 8.30 ± 0.84 b | 0.59 ± 0.04 a | 1.42 ± 0.11 b | 4.13 ± 0.17 a | 7.61 ± 0.32 b | ND | ND | |
| Σ SFA | TG | 22.55 ± 0.84 a | 26.83 ± 1.43 b | 15.99 ± 0.42 a | 19.34 ± 0.56 b | 19.03 ± 0.34 a | 23.00 ± 1.61 b | 15.18 ± 0.34 a | 19.33 ± 0.74 b |
| PL | 41.46 ± 0.48 a | 38.75 ± 1.03 b | 43.27 ± 0.97 a | 42.50 ± 0.77 a | 35.43 ± 3.55 a | 37.42 ± 0.61 a | ND | ND | |
| Σ MUFA | TG | 64.50 ± 0.81 a | 30.98 ± 2.00 b | 65.08 ± 0.41 a | 36.17 ± 1.33 b | 62.44 ± 0.59 a | 39.07 ± 1.84 b | 67.10 ± 0.18 a | 34.16 ± 2.13 b |
| PL | 28.30 ± 0.79 a | 13.19 ± 0.89 b | 16.14 ± 0.38 a | 6.18 ± 0.64 b | 30.08 ± 6.38 a | 7.08 ± 0.65 b | ND | ND | |
| Σ n-6 PUFA | TG | 12.32 ± 1.35 a | 18.29 ± 0.35 b | 18.09 ± 0.55 a | 24.19 ± 0.68 b | 17.44 ± 0.73 a | 20.41 ± 0.79 b | 16.56 ± 0.33 a | 23.87 ± 0.62 b |
| PL | 25.82 ± 0.71 a | 20.30 ± 1.20 b | 39.43 ± 1.08 a | 38.76 ± 0.86 a | 29.88 ± 2.23 a | 35.73 ± 0.89 b | ND | ND | |
| Σ n-3 PUFA | TG | 0.54 ± 0.11 a | 4.52 ± 1.10 b | 0.76 ± 0.04 a | 6.05 ± 1.27 b | 0.79 ± 0.07 a | 4.20 ± 0.73 b | 1.09 ± 0.04 a | 5.98 ± 1.31 b |
| PL | 4.35 ± 0.53 a | 10.97 ± 0.97 b | 1.06 ± 0.09 a | 3.25 ± 0.31 b | 4.45 ± 0.72 a | 10.23 ± 0.51 b | ND | ND | |
| Σ CLA | TG | 0.02 ± 0.01 a | 9.99 ± 1.10 b | 0.02 ± 0.002 a | 9.12 ± 0.86 b | 0.04 ± 0.004 a | 8.79 ± 1.10 b | 0.01 ± 0.005 a | 10.23 ± 1.20 b |
| PL | 0.02 ± 0.002 a | 8.40 ± 1.10 b | Traces a | 5.85 ± 0.48 b | 0.03 ± 0.01 a | 5.48 ± 0.50 b | ND | ND | |
| Σ CLnA | TG | ND a | 8.25 ± 1.57 b | ND a | 4.15 ± 0.30 b | ND a | 3.59 ± 0.33 b | ND a | 4.72 ± 0.46 b |
| PL | ND a | 7.84 ± 1.73 b | ND a | 3.00 ± 0.52 b | ND a | 3.75 ± 0.58 b | ND | ND | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngo Njembe, M.T.; Dejonghe, L.; Verstraelen, E.; Mignolet, E.; Leclercq, M.; Dailly, H.; Gardin, C.; Buchet, M.; Waingeh Nain, C.; Larondelle, Y. The Egg Yolk Content in ω-3 and Conjugated Fatty Acids Can Be Sustainably Increased upon Long-Term Feeding of Laying Hens with a Diet Containing Flaxseeds and Pomegranate Seed Oil. Foods 2021, 10, 1134. https://doi.org/10.3390/foods10051134
Ngo Njembe MT, Dejonghe L, Verstraelen E, Mignolet E, Leclercq M, Dailly H, Gardin C, Buchet M, Waingeh Nain C, Larondelle Y. The Egg Yolk Content in ω-3 and Conjugated Fatty Acids Can Be Sustainably Increased upon Long-Term Feeding of Laying Hens with a Diet Containing Flaxseeds and Pomegranate Seed Oil. Foods. 2021; 10(5):1134. https://doi.org/10.3390/foods10051134
Chicago/Turabian StyleNgo Njembe, Monique T., Louis Dejonghe, Eleonore Verstraelen, Eric Mignolet, Matthieu Leclercq, Hélène Dailly, Cécile Gardin, Marine Buchet, Caroline Waingeh Nain, and Yvan Larondelle. 2021. "The Egg Yolk Content in ω-3 and Conjugated Fatty Acids Can Be Sustainably Increased upon Long-Term Feeding of Laying Hens with a Diet Containing Flaxseeds and Pomegranate Seed Oil" Foods 10, no. 5: 1134. https://doi.org/10.3390/foods10051134
APA StyleNgo Njembe, M. T., Dejonghe, L., Verstraelen, E., Mignolet, E., Leclercq, M., Dailly, H., Gardin, C., Buchet, M., Waingeh Nain, C., & Larondelle, Y. (2021). The Egg Yolk Content in ω-3 and Conjugated Fatty Acids Can Be Sustainably Increased upon Long-Term Feeding of Laying Hens with a Diet Containing Flaxseeds and Pomegranate Seed Oil. Foods, 10(5), 1134. https://doi.org/10.3390/foods10051134