
Effect of Gaseous Ozone on Listeria monocytogenes Planktonic Cells and Biofilm: An In Vitro Study


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. Screening of Biofilm Forming Strains: Micro-Method Assays
2.3. Biofilm Production Quantification: Macro-Method Assays
2.3.1. Biofilm Production Indices (BPIs)
2.3.2. Quantification of Viable Bacteria in the Biofilm
2.4. Ozonization Assays
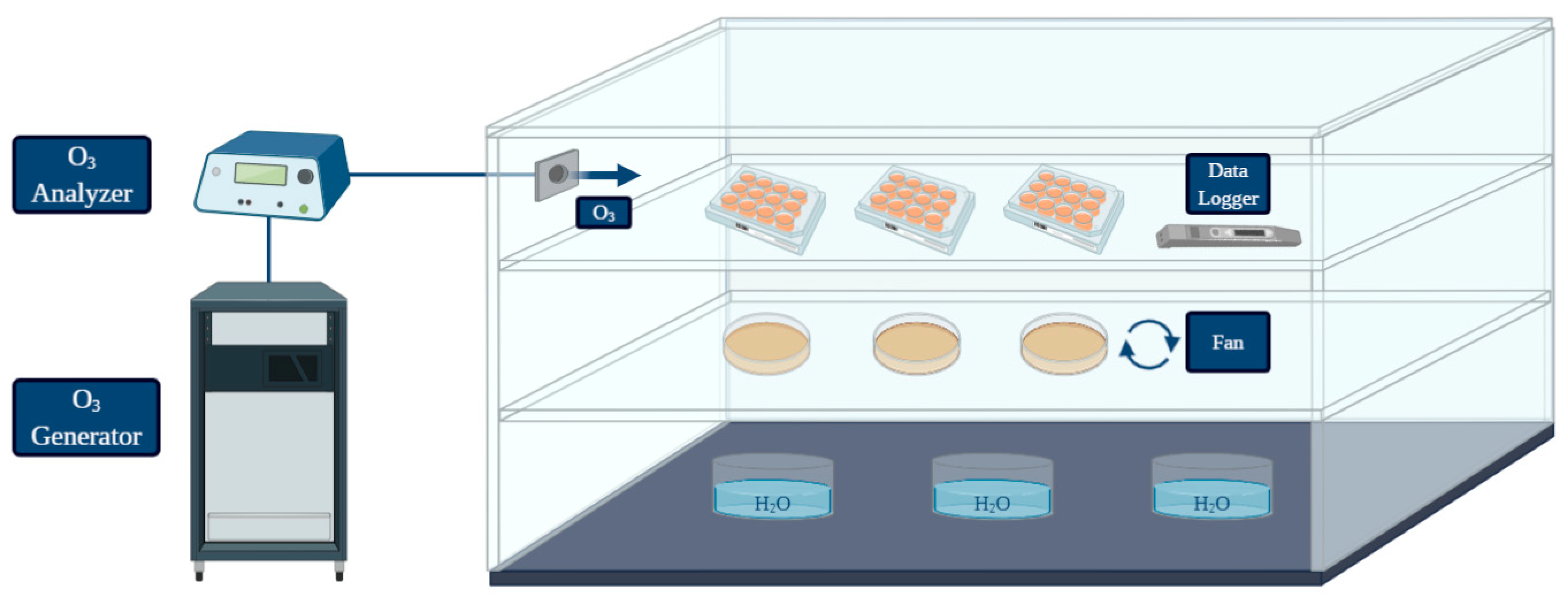
2.4.1. Treatment Chamber and Experimental Conditions
2.4.2. Effect of Gaseous Ozone on L. monocytogenes Planktonic Cells
2.4.3. Biofilm Inhibition by Gaseous Ozone
2.4.4. Biofilm Eradication by Gaseous Ozone
2.5. Statistical Analyses and Graphing
3. Results
3.1. Screening of Biofilm Forming Strains: Micro-Method Assays
3.2. Effects of O3 on L. monocytogenes Planktonic Cells
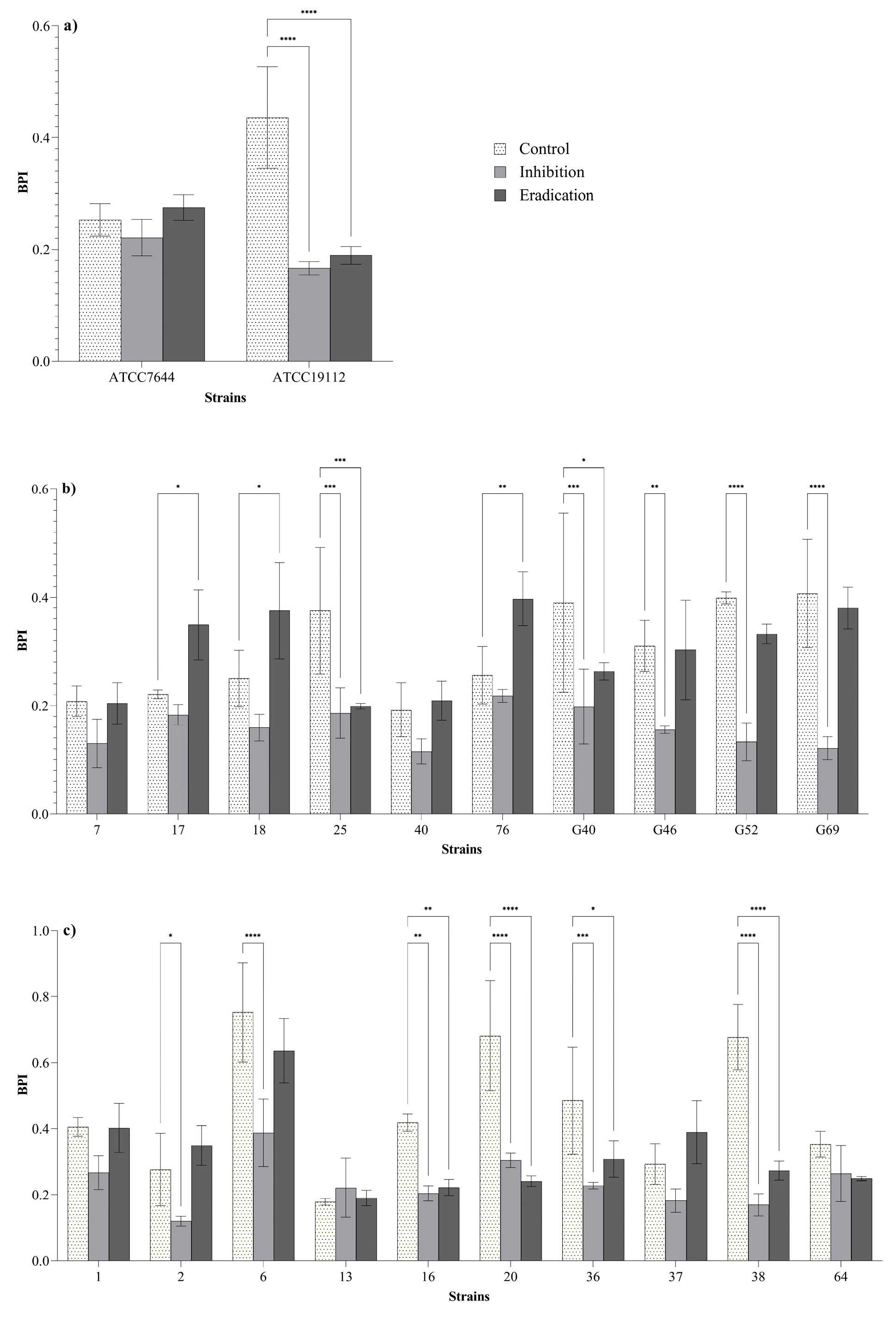
3.3. Effect of O3 on Biofilm
3.3.1. Effect on the Biofilm Biomass
3.3.2. Effect on Viable Biofilm-Detached Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- González-Rivas, F.; Ripolles-Avila, C.; Fontecha-Umaña, F.; Ríos-Castillo, A.G.; Rodríguez-Jerez, J.J. Biofilms in the spotlight: Detection, quantification, and removal methods. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1261–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Abebe, G.M. The Role of Bacterial Biofilm in Antibiotic Resistance and Food Contamination. Int. J. Microbiol. 2020, 1705814. [Google Scholar] [CrossRef] [PubMed]
- Skowron, K.; Wałecka-Zacharska, E.; Wiktorczyk-Kapischke, N.; Skowron, K.J.; Grudlewska-Buda, K.; Bauza-Kaszewska, J.; Bernaciak, Z.; Borkowski, M.; Gospodarek-Komkowska, E. Assessment of the Prevalence and Drug Susceptibility of Listeria monocytogenes Strains Isolated from Various Types of Meat. Foods 2020, 9, 1293. [Google Scholar] [CrossRef]
- European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union one health 2018 zoonoses report. EFSA J. 2019, 17, e05926. [Google Scholar] [CrossRef] [Green Version]
- Buchanan, R.L.; Gorris, L.G.M.; Hayman, M.M.; Jackson, T.C.; Whiting, R.C. A review of Listeria monocytogenes: An update on outbreaks, virulence, dose-response, ecology, and risk assessments. Food Control 2017, 75, 1–13. [Google Scholar] [CrossRef]
- Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Herman, L.; Koutsoumanis, K.; Nørrung, B.; et al. Listeria monocytogenes contamination of ready-to-eat foods and the risk for human health in the EU. EFSA J. 2018, 16, e05134. [Google Scholar] [CrossRef] [PubMed]
- Colagiorgi, A.; Bruini, I.; di Ciccio, P.A.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes biofilms in the wonderland of food industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, G.; Piccolomini, R.; Paludi, D.; D’Orio, V.; Vergara, A.; Conter, M.; Ianieri, A. Influence of temperature on biofilm formation by Listeria monocytogenes on various food-contact surfaces: Relationship with motility and cell surface hydrophobicity. J. Appl. Microbiol. 2008, 104, 1552–1561. [Google Scholar] [CrossRef]
- Fagerlund, A.; Langsrud, S.; Møretrø, T. Microbial diversity and ecology of biofilms in food industry environments associated with Listeria monocytogenes persistence. Curr. Opin. Food Sci. 2021, 37, 171–178. [Google Scholar] [CrossRef]
- Giaouris, E.; Heir, E.; Hébraud, M.; Chorianopoulos, N.; Langsrud, S.; Møretrø, T.; Habimana, O.; Desvaux, M.; Renier, S.; Nychas, G.J. Attachment and biofilm formation by foodborne bacteria in meat processing environments: Causes, implications, role of bacterial interactions and control by alternative novel methods. Meat Sci. 2014, 97, 298–309. [Google Scholar] [CrossRef] [PubMed]
- Leong, D.; NicAogáin, K.; Luque-Sastre, L.; McManamon, O.; Hunt, K.; Alvarez-Ordóñez, A.; Scollard, J.; Schmalenberger, A.; Fanning, S.; O’Byrne, C.; et al. A 3-year multi-food study of the presence and persistence of Listeria monocytogenes in 54 small food businesses in Ireland. Int. J. Food Microbiol. 2017, 249, 18–26. [Google Scholar] [CrossRef]
- Mazaheri, T.; Cervantes-Huamán, B.R.; Bermúdez-Capdevila, M.; Ripolles-Avila, C.; Rodríguez-Jerez, J.J. Listeria monocytogenes Biofilms in the Food Industry: Is the Current Hygiene Program Sufficient to Combat the Persistence of the Pathogen? Microorganisms 2021, 9, 181. [Google Scholar] [CrossRef]
- Oniciuc, E.A.; Likotrafiti, E.; Alvarez-Molina, A.; Prieto, M.; López, M.; Alvarez-Ordóñez, A. Food processing as a risk factor for antimicrobial resistance spread along the food chain. Curr. Opin. Food Sci. 2019, 30, 21–26. [Google Scholar] [CrossRef]
- Martínez-Suárez, J.V.; Ortiz, S.; López-Alonso, V. Potential impact of the resistance to quaternary ammonium disinfectants on the persistence of Listeria monocytogenes in food processing environments. Front. Microbiol. 2016, 7, 638. [Google Scholar] [CrossRef] [Green Version]
- Møretrø, T.; Schirmer, B.C.T.; Heir, E.; Fagerlund, A.; Hjemli, P.; Langsrud, S. Tolerance to quaternary ammonium compound disinfectants may enhance growth of Listeria monocytogenes in the food industry. Int. J. Food Microbiol. 2017, 241, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Guérin, A.; Bridier, A.; Le Grandois, P.; Sévellec, Y.; Palma, F.; Félix, B.; LISTADAPT Study Group; Roussel, S.; Soumet, C. Exposure to quaternary ammonium compounds selects resistance to ciprofloxacin in Listeria monocytogenes. Pathogens 2021, 10, 220. [Google Scholar] [CrossRef]
- Marino, M.; Maifreni, M.; Baggio, A.; Innocente, N. Inactivation of foodborne bacteria biofilms by aqueous and gaseous ozone. Front. Microbiol. 2018, 9, 2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colagiorgi, A.; Festa, R.; di Ciccio, P.A.; Gogliettino, M.; Balestrieri, M.; Palmieri, G.; Anastasio, A.; Ianieri, A. Rapid biofilm eradication of the antimicrobial peptide 1018-K6 against Staphylococcus aureus: A new potential tool to fight bacterial biofilms. Food Control 2020, 107, 106815. [Google Scholar] [CrossRef]
- Meireles, A.; Borges, A.; Giaouris, E.; Simões, M. The current knowledge on the application of anti-biofilm enzymes in the food industry. Food Res. Int. 2016, 86, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-Pichardo, R.; Hernández-Martínez, I.; Regalado-González, C.; Santos-Cruz, J.; Meas-Vong, Y.; Wacher-Rodarte, M.D.C.; Carrillo-Reyes, J.; Sánchez-Ortega, I.; García-Almendárez, B.E. Innovative Control of Biofilms on Stainless Steel Surfaces Using Electrolyzed Water in the Dairy Industry. Foods 2021, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, C.; Cui, H.; Lin, L. Feasibility of cold plasma for the control of biofilms in food industry. Trends Food Sci. Technol. 2020, 99, 142–151. [Google Scholar] [CrossRef]
- Botta, C.; Ferrocino, I.; Pessione, A.; Cocolin, L.; Rantsiou, K. Spatiotemporal distribution of the environmental microbiota in food processing plants as impacted by cleaning and sanitizing procedures: The case of slaughterhouses and gaseous ozone. Appl. Environ. Microbiol. 2020, 86, e01861-20. [Google Scholar] [CrossRef] [PubMed]
- Sivaranjani, S.; Prasath, V.A.; Pandiselvam, R.; Kothakota, A.; Khaneghah, A.M. Recent advances in applications of ozone in the cereal industry. LWT 2021, 111412. [Google Scholar] [CrossRef]
- Pascual, A.; Llorca, I.; Canut, A. Use of ozone in food industries for reducing the environmental impact of cleaning and disinfection activities. Trends Food Sci. Technol. 2007, 18, S29–S35. [Google Scholar] [CrossRef]
- Pandiselvam, R.; Subhashini, S.; Banuu Priya, E.P.; Kothakota, A.; Ramesh, S.V.; Shahir, S. Ozone based food preservation: A promising green technology for enhanced food safety. Ozone Sci. Eng. 2019, 41, 17–34. [Google Scholar] [CrossRef]
- Brodowska, A.J.; Nowak, A.; Śmigielski, K. Ozone in the food industry: Principles of ozone treatment, mechanisms of action, and applications: An overview. Crit. Rev. Food Sci. Nutr. 2018, 58, 2176–2201. [Google Scholar] [CrossRef] [PubMed]
- Varga, L.; Szigeti, J. Use of ozone in the dairy industry: A review. Int. J. Dairy Technol. 2016, 69, 157–168. [Google Scholar] [CrossRef]
- Sarron, E.; Gadonna-Widehem, P.; Aussenac, T. Ozone Treatments for Preserving Fresh Vegetables Quality: A Critical Review. Foods 2021, 10, 605. [Google Scholar] [CrossRef]
- Harada, A.M.; Nascimento, M.S. Efficacy of dry sanitizing methods on Listeria monocytogenes biofilms. Food Control 2021, 124, 107897. [Google Scholar] [CrossRef]
- Nicholas, R.; Dunton, P.; Tatham, A.; Fielding, L. The effect of ozone and open air factor on surface-attached and biofilm environmental Listeria monocytogenes. J. Appl. Microbiol. 2013, 115, 555–564. [Google Scholar] [CrossRef]
- Robbins, J.B.; Fisher, C.W.; Moltz, A.G.; Martin, S.E. Elimination of Listeria monocytogenes biofilms by ozone, chlorine, and hydrogen peroxide. J. Food Prot. 2005, 68, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Giménez, B.; Graiver, N.; Giannuzzi, L.; Zaritzky, N. Treatment of beef with gaseous ozone: Physicochemical aspects and antimicrobial effects on heterotrophic microflora and Listeria monocytogenes. Food Control 2021, 121, 107602. [Google Scholar] [CrossRef]
- Carpentier, B.; Cerf, O. Review—Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 2011, 145, 1–8. [Google Scholar] [CrossRef]
- Aalto-Araneda, M.; Pöntinen, A.; Pesonen, M.; Corander, J.; Markkula, A.; Tasara, T.; Stephan, R.; Korkeala, H. Strain variability of Listeria monocytogenes under NaCl stress elucidated by a high-throughput microbial growth data assembly and analysis protocol. Appl. Environ. Microbiol. 2020, 86, e02378-19. [Google Scholar] [CrossRef] [PubMed]
- Bucur, F.I.; Grigore-Gurgu, L.; Crauwels, P.; Riedel, C.U.; Nicolau, A.I. Resistance of Listeria monocytogenes to stress conditions encountered in food and food processing environments. Front. Microbiol. 2018, 9, 2700. [Google Scholar] [CrossRef] [Green Version]
- Hingston, P.; Chen, J.; Dhillon, B.K.; Laing, C.; Bertelli, C.; Gannon, V.; Tasara, T.; Allen, K.; Brinkman, F.S.L.; Hansen, L.T.; et al. Genotypes associated with Listeria monocytogenes isolates displaying impaired or enhanced tolerances to cold, salt, acid, or desiccation stress. Front. Microbiol. 2017, 8, 369. [Google Scholar] [CrossRef] [Green Version]
- Lianou, A.; Nychas, G.J.E.; Koutsoumanis, K.P. Strain variability in biofilm formation: A food safety and quality perspective. Food Res. Int. 2020, 137, 109424. [Google Scholar] [CrossRef]
- Piercey, M.J.; Ells, T.C.; Macintosh, A.J.; Truelstrup Hansen, L. Variations in biofilm formation, desiccation resistance and benzalkonium chloride susceptibility among Listeria monocytogenes strains isolated in Canada. Int. J. Food Microbiol. 2017, 257, 254–261. [Google Scholar] [CrossRef] [Green Version]
- Poimenidou, S.V.; Chrysadakou, M.; Tzakoniati, A.; Bikouli, V.C.; Nychas, G.J.; Skandamis, P.N. Variability of Listeria monocytogenes strains in biofilm formation on stainless steel and polystyrene materials and resistance to peracetic acid and quaternary ammonium compounds. Int. J. Food Microbiol. 2016, 237, 164–171. [Google Scholar] [CrossRef]
- Harter, E.; Wagner, E.M.; Zaiser, A.; Halecker, S.; Wagner, M.; Rychli, K. Stress survival islet 2, predominantly present in Listeria monocytogenes strains of sequence type 121, is involved in the alkaline and oxidative stress responses. Appl. Environ. Microbiol. 2017, 83, e00827-17. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Morvay, A.A.; Shi, X.; Suo, Y.; Shi, C.; Knøchel, S. Comparison of oxidative stress response and biofilm formation of Listeria monocytogenes serotypes 4b and 1/2a. Food Control 2018, 85, 416–422. [Google Scholar] [CrossRef]
- Muhterem-Uyar, M.; Ciolacu, L.; Wagner, K.H.; Wagner, M.; Schmitz-Esser, S.; Stessl, B. New aspects on Listeria monocytogenes ST5-ECVI predominance in a heavily contaminated cheese processing environment. Front. Microbiol. 2018, 9, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naditz, A.L.; Dzieciol, M.; Wagner, M.; Schmitz-Esser, S. Plasmids contribute to food processing environment–associated stress survival in three Listeria monocytogenes ST121, ST8, and ST5 strains. Int. J. Food Microbiol 2019, 299, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, S.; Begley, M.; Hill, C.; Gahan, C.G.M. A five-gene stress survival islet (SSI-1) that contributes to the growth of Listeria monocytogenes in suboptimal conditions. J. Appl. Microbiol. 2010, 109, 984–995. [Google Scholar] [CrossRef] [PubMed]
- Henriques, A.R.; Fraqueza, M.J. Biofilm-forming ability and biocide susceptibility of Listeria monocytogenes strains isolated from the ready-to-eat meat-based food products food chain. LWT 2017, 81, 180–187. [Google Scholar] [CrossRef]
- In Lee, S.H.; Barancelli, G.V.; de Camargo, T.M.; Corassin, C.H.; Rosim, R.E.; da Cruz, A.G.; Cappato, L.P.; de Oliveira, C.A.F. Biofilm-producing ability of Listeria monocytogenes isolates from Brazilian cheese processing plants. Food Res. Int. 2017, 91, 88–91. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Zand, E.; Pfanner, H.; Domig, K.J.; Sinn, G.; Zunabovic-Pichler, M.; Jaeger, H. Biofilm-Forming Ability of Microbacterium lacticum and Staphylococcus capitis Considering Physicochemical and Topographical Surface Properties. Foods 2021, 10, 611. [Google Scholar] [CrossRef] [PubMed]
- BioRender. Available online: https://biorender.com/ (accessed on 27 May 2021).
- Guzzon, R.; Franciosi, E.; Carafa, I.; Larcher, R.; Tuohy, K.M. L’ozono, un innovativo strumento per il controllo microbiologico in aziende lattiero-casearie tradizionali. Ind. Aliment. 2015, 54, 11–15. [Google Scholar]
- Djordjevic, D.; Wiedmann, M.; McLandsborough, L.A. Microtiter plate assay for assessment of Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 2012, 68, 2950–2958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadam, S.R.; den Besten, H.M.W.; van der Veen, S.; Zwietering, M.H.; Moezelaar, R.; Abee, T. Diversity assessment of Listeria monocytogenes biofilm formation: Impact of growth condition, serotype and strain origin. Int. J. Food Microbiol. 2013, 165, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, R.E.; Ross, T.; Bowman, J.P. Variability in biofilm production by Listeria monocytogenes correlated to strain origin and growth conditions. Int. J. Food Microbiol. 2011, 150, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Abeysundara, P.D.A.; Nannapaneni, R.; Soni, K.A.; Sharma, C.S.; Mahmoud, B. Induction and stability of oxidative stress adaptation in Listeria monocytogenes EGD (Bug600) and F1057 in sublethal concentrations of H2O2 and NaOH. Int. J. Food Microbiol. 2016, 238, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Maggio, F.; Rossi, C.; Chaves-López, C.; Serio, A.; Valbonetti, L.; Pomilio, F.; Chiavaroli, A.P.; Paparella, A. Interactions between L. monocytogenes and P. fluorescens in Dual-Species Biofilms under Simulated Dairy Processing Conditions. Foods 2021, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Bigi, F.; Haghighi, H.; Quartieri, A.; De Leo, R.; Pulvirenti, A. Impact of low-dose gaseous ozone treatment to reduce the growth of in vitro broth cultures of foodborne pathogenic/spoilage bacteria in a food storage cold chamber. J. Food Saf. 2021, e12892. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Category | Strain ID (Internal) | Strain ID (NCBI) 1 | Source | Lineage | Serogroup | Sequence Type (ST) | Clonal Complex (CC) |
|---|---|---|---|---|---|---|---|
| ATCC | ATCC 7644 | ATCC 7644 | Human | II | IIc | 122 | 9 |
| ATCC 19112 | WSLC1001 | Human | II | IIc | 12 | 7 | |
| Dairy | 17 | CFSAN045778 | Product | II | IIa | 9 | 9 |
| 25 | CFSAN045791 | Product | II | IIc | 9 | 9 | |
| 76 | CFSAN044775 | Product | II | IIc | 9 | 9 | |
| G40 | CFSAN044840 | Product | II | IIa | 325 | 31 | |
| G52 | CFSAN044807 | Product | II | IIa | 325 | 31 | |
| 7 | CFSAN045850 | Production Environment | II | IIa | 325 | 31 | |
| 18 | CFSAN045794 | Production Environment | II | IIa | 9 | 9 | |
| 40 | CFSAN044857 | Production Environment | II | IIa | 325 | 31 | |
| G46 | CFSAN044805 | Production Environment | II | IIa | 325 | 31 | |
| G69 | CFSAN044813 | Production Environment | II | IIa | 325 | 31 | |
| Meat | 16 | CFSAN045938 | Product | II | IIa | 325 | 31 |
| 20 | CFSAN045829 | Product | II | IIa | 9 | 9 | |
| 36 | CFSAN044741 | Product | II | IIc | 9 | 9 | |
| 38 | CFSAN044748 | Product | II | IIc | 9 | 9 | |
| 64 | CFSAN044767 | Product | II | IIc | 9 | 9 | |
| 1 | CFSAN046012 | Production Environment | II | IIc | 9 | 9 | |
| 2 | CFSAN045995 | Production Environment | II | IIa | 325 | 31 | |
| 6 | CFSAN045858 | Production Environment | II | IIc | 9 | 9 | |
| 13 | CFSAN045971 | Production Environment | II | IIa | 9 | 9 | |
| 37 | CFSAN046048 | Production Environment | II | IIc | 9 | 9 |
| Strain ID | Logarithmic Reduction 1,2 | ||
|---|---|---|---|
| 10 min | 30 min | 6 h | |
| ATCC 7644 | 3.2 ± 0.1 a | 3.6 ± 0.3 a | 7.2 ± 0.1 b |
| ATCC 19112 | 3.1 ± 0.2 a | 3.1 ± 0.1 a | 7.5 ± 0.3 b |
| 1 | 3.1 ± 0.3 a | 3.5 ± 0.3 a | ≥7.7 ± 0.1 b |
| 2 | 3.9 ± 0.1 a | 4.3 ± 0.1 a | ≥9.2 ± 0.0 b |
| 6 | 4.2 ± 0.1 a | 4.4 ± 0.1 a | ≥9.2 ± 0.1 b |
| 7 | 3.7 ± 0.2 a | 3.9 ± 0.1 a | ≥9.0 ± 0.1 b |
| 13 | 4.1 ± 0.1 a | 4.0 ± 0.2 a | ≥9.2 ± 0.1 b |
| 16 | 3.3 ± 0.2 a | 3.3 ± 0.1 a | ≥8.2 ± 0.0 b |
| 17 | 3.6 ± 0.0 a | 3.5 ± 0.1 a | 7.4 ± 0.1 b |
| 18 | 4.5 ± 0.1 b | 4.0 ± 0.2 a | ≥9.1 ± 0.1 c |
| 20 | 3.6 ± 0.1 a | 4.2 ± 0.1 b | ≥8.4 ± 0.0 c |
| 25 | 3.5 ± 0.4 a | 3.5 ± 0.1 a | ≥8.2 ± 0.1 b |
| 36 | 3.3 ± 0.1 a | 3.7 ± 0.1 a | ≥8.7 ± 0.1 b |
| 37 | 4.1 ± 0.1 a | 4.2 ± 0.0 a | ≥9.2 ± 0.0 b |
| 38 | 4.1 ± 0.3 a | 4.1 ± 0.3 a | 7.2 ± 0.4 b |
| 40 | 4.1 ± 0.3 a | 3.9 ± 0.1 a | 8.2 ± 0.1 b |
| 64 | 3.7 ± 0.2 a | 4.1 ± 0.4 a | ≥8.8 ± 0.2 b |
| 76 | 3.7 ± 0.2 a | 4.1 ± 0.1 a | ≥8.9 ± 0.1 b |
| G40 | 3.5 ± 0.2 a | 3.5 ± 0.1 a | ≥8.6 ± 0.1 b |
| G46 | 4.1 ± 0.2 a | 4.2 ± 0.1 a | ≥9.1 ± 0.1 b |
| G52 | 4.1 ± 0.2 a | 4.3 ± 0.2 a | ≥9.1 ± 0.2 b |
| G69 | 3.9 ± 0.0 a | 4.1 ± 0.1 a | ≥9.1 ± 0.0 b |
| Strain ID | Logarithmic Reduction 1,2 | |
|---|---|---|
| Inhibition | Eradication | |
| ATCC 7644 | 0.6 ± 0.1 | 0.6 ± 0.1 |
| ATCC 19112 | 0.7 ± 0.1 b | 1.2 ± 0.1 a |
| 1 | 1.0 ± 0.0 | 1.1 ± 0.0 |
| 2 | 1.2 ± 0.1 | 1.1 ± 0.1 |
| 6 | 0.4 ± 0.1 a | 0.1 ± 0.1 b |
| 7 | 1.3 ± 0.1 | 1.3 ± 0.1 |
| 13 | 0.2 ± 0.1 | 0.2 ± 0.1 |
| 16 | 1.1 ± 0.1 | 1.2 ± 0.1 |
| 17 | 0.6 ± 0.1 b | 0.9 ± 0.1 a |
| 18 | 1.6 ± 0.0 | 1.7 ± 0.1 |
| 20 | 0.6 ± 0.0 a | 0.1 ± 0.0 b |
| 25 | 0.9 ± 0.0 | 1.0 ± 0.0 |
| 36 | 0.9 ± 0.1 b | 1.2 ± 0.1 a |
| 37 | 0.7 ± 0.1 | 0.6 ± 0.1 |
| 38 | 0.7 ± 0.1 a | 0.3 ± 0.2 b |
| 40 | 0.1 ± 0.1 | 0.2 ± 0.1 |
| 64 | 0.6 ± 0.1 | 0.4 ± 0.1 |
| 76 | 0.6 ± 0.1 b | 1.3 ± 0.2 a |
| G40 | 0.9 ± 0.0 | 1.1 ± 0.2 |
| G46 | 0.4 ± 0.1 | 0.6 ± 0.1 |
| G52 | 1.0 ± 0.1 | 1.0 ± 0.1 |
| G69 | 0.4 ± 0.2 a | 0.0 ± 0.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panebianco, F.; Rubiola, S.; Chiesa, F.; Civera, T.; Di Ciccio, P.A. Effect of Gaseous Ozone on Listeria monocytogenes Planktonic Cells and Biofilm: An In Vitro Study. Foods 2021, 10, 1484. https://doi.org/10.3390/foods10071484
Panebianco F, Rubiola S, Chiesa F, Civera T, Di Ciccio PA. Effect of Gaseous Ozone on Listeria monocytogenes Planktonic Cells and Biofilm: An In Vitro Study. Foods. 2021; 10(7):1484. https://doi.org/10.3390/foods10071484
Chicago/Turabian StylePanebianco, Felice, Selene Rubiola, Francesco Chiesa, Tiziana Civera, and Pierluigi Aldo Di Ciccio. 2021. "Effect of Gaseous Ozone on Listeria monocytogenes Planktonic Cells and Biofilm: An In Vitro Study" Foods 10, no. 7: 1484. https://doi.org/10.3390/foods10071484
APA StylePanebianco, F., Rubiola, S., Chiesa, F., Civera, T., & Di Ciccio, P. A. (2021). Effect of Gaseous Ozone on Listeria monocytogenes Planktonic Cells and Biofilm: An In Vitro Study. Foods, 10(7), 1484. https://doi.org/10.3390/foods10071484