
Anti-Viral and Immunomodulatory Properties of Propolis: Chemical Diversity, Pharmacological Properties, Preclinical and Clinical Applications, and In Silico Potential against SARS-CoV-2

 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Ethnopharmacology
3. Chemical Composition of Propolis
4. Anti-Viral Activity
5. Propolis as a Treatment for COVID-19
6. Immunomodulatory Activity
7. Clinical Applications of Propolis as an Antiviral and Immunomodulatory Agents
8. In Silico Drug Discovery
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pavlovic, R.; Borgonovo, G.; Leoni, V.; Giupponi, L.; Ceciliani, G.; Sala, S.; Bassoli, A.; Giorgi, A. Effectiveness of different analytical methods for the characterization of propolis: A case of study in Northern Italy. Molecules 2020, 25, 504. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.G., Jr.; Yoshida, N.C.; Garcez, W.S.; Perdomo, R.T.; Matos, M.d.F.C.; Garcez, F.R. Metabolomics approach expands the classification of propolis samples from midwest Brazil. J. Nat. Prod. 2020, 83, 333–343. [Google Scholar] [CrossRef]
- Soroy, L.; Bagus, S.; Yongkie, I.P.; Djoko, W. The effect of a unique propolis compound (PropoelixTM) on clinical outcomes in patients with dengue hemorrhagic fever. Infect. Drug Resist. 2014, 7, 323–329. [Google Scholar]
- Jasprica, I.; Mornar, A.; Debeljak, Ž.; Smolčić-Bubalo, A.; Medić-Šarić, M.; Mayer, L.; Romić, Ž.; Bućan, K.; Balog, T.; Sobočanec, S. In vivo study of propolis supplementation effects on antioxidative status and red blood cells. J. Ethnopharmacol. 2007, 110, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Sforcin, J.M.; Bankova, V. Propolis: Is there a potential for the development of new drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef]
- Paulino, N.; Dantas, A.P.; Bankova, V.; Longhi, D.T.; Scremin, A.; de Castro, S.L.; Calixto, J.B. Bulgarian propolis induces analgesic and anti-inflammatory effects in mice and inhibits in vitro contraction of airway smooth muscle. J. Pharmacol. Sci. 2003, 93, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Liu, J.; Hu, Y.; Fan, Y.; Wang, D.; Guo, L.; Nguyen, T.L.; Zhao, X.; Liu, X.; Liu, C. The immunological activity of propolis flavonoids liposome on the immune response against ND vaccine. Int. J. Biol. Macromol. 2012, 51, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Burdock, G.A. Review of the biological properties and toxicity of bee propolis (propolis). Food Chem. Toxicol. 1998, 36, 347–363. [Google Scholar] [CrossRef]
- Skaba, D.; Morawiec, T.; Tanasiewicz, M.; Mertas, A.; Bobela, E.; Szliszka, E.; Skucha-Nowak, M.; Dawiec, M.; Yamamoto, R.; Ishiai, S.; et al. Influence of the toothpaste with Brazilian ethanol extract propolis on the oral cavity health. Evid. Based Complement. Altern. Med. 2013, 2013, 215391. [Google Scholar] [CrossRef]
- Sforcin, J.M. Biological properties and therapeutic applications of propolis. Phyther. Res. 2016, 30, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Pasupuleti, V.R.; Sammugam, L.; Ramesh, N.; Gan, S.H. Honey, propolis, and royal jelly: A comprehensive review of their biological actions and health benefits. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Amoros, M.; Sauvager, F.; Girre, L.; Cormier, M. In vitro antiviral activity of propolis. Apidologie 1992, 23, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Schnitzler, P.; Neuner, A.; Nolkemper, S.; Zundel, C.; Nowack, H.; Sensch, K.H.; Reichling, J. Antiviral activity and mode of action of propolis extracts and selected compounds. Phyther. Res. 2010, 24, S20–S28. [Google Scholar] [CrossRef]
- Kujumgiev, A.; Tsvetkova, I.; Serkedjieva, Y.; Bankova, V.; Christov, R.; Popov, S. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J. Ethnopharmacol. 1999, 64, 235–240. [Google Scholar] [CrossRef]
- Kwon, M.J.; Shin, H.M.; Perumalsamy, H.; Wang, X.; Ahn, Y.-J. Antiviral effects and possible mechanisms of action of constituents from Brazilian propolis and related compounds. J. Apic. Res. 2020, 59, 413–425. [Google Scholar] [CrossRef]
- Serkedjieva, J.; Manolova, N.; Bankova, V. Anti-influenza virus effect of some propolis constituents and their analogues (esters of substituted cinnamic acids). J. Nat. Prod. 1992, 55, 294–297. [Google Scholar] [CrossRef]
- Debiaggi, M.; Tateo, F.; Pagani, L.; Luini, M.; Romero, E. Effects of propolis flavonoids on virus infectivity and replication. Microbiologica 1990, 13, 207–213. [Google Scholar] [PubMed]
- Refaat, H.; Mady, F.M.; Sarhan, H.A.; Rateb, H.S.; Alaaeldin, E. Optimization and evaluation of propolis liposomes as a promising therapeutic approach for COVID-19. Int. J. Pharm. 2020, 120028. [Google Scholar] [CrossRef]
- Sahlan, M.; Irdiani, R.; Flamandita, D.; Aditama, R.; Alfarraj, S.; Javed Ansari, M.; Cahya Khayrani, A.; Kartika Pratami, D.; Lischer, K. Molecular interaction analysis of Sulawesi propolis compounds with SARS-CoV-2 main protease as preliminary study for COVID-19 drug discovery. J. King Saud Univ. Sci. 2020, 101234. [Google Scholar] [CrossRef]
- Hashem, H. In silico approach of some selected honey constituents as SARS-CoV-2 main protease (COVID-19) inhibitors. ChemRxiv. 2020. [Google Scholar] [CrossRef]
- Zabaiou, N.; Fouache, A.; Trousson, A.; Baron, S.; Zellagui, A.; Lahouel, M.; Lobaccaro, J.-M.A. Biological properties of propolis extracts: Something new from an ancient product. Chem. Phys. Lipids 2017, 207, 214–222. [Google Scholar] [CrossRef]
- Zaccaria, V.; Garzarella, E.U.; Di Giovanni, C.; Galeotti, F.; Gisone, L.; Campoccia, D.; Volpi, N.; Arciola, C.R.; Daglia, M. Multi dynamic extraction: An innovative method to obtain a standardized chemically and biologically reproducible polyphenol extract from poplar-type propolis to be used for its anti-infective properties. Materials (Basel) 2019, 12, 3746. [Google Scholar] [CrossRef] [Green Version]
- Popova, M.; Trusheva, B.; Bankova, V. Propolis of stingless bees: A phytochemist’s guide through the jungle of tropical biodiversity. Phytomedicine 2019, 86, 153098. [Google Scholar] [CrossRef]
- Tolba, M.F.; Azab, S.S.; Khalifa, A.E.; Abdel-Rahman, S.Z.; Abdel-Naim, A.B. Caffeic acid phenethyl ester, a promising component of propolis with a plethora of biological activities: A review on its anti-inflammatory, neuroprotective, hepatoprotective, and cardioprotective effects. IUBMB Life 2013, 65, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Tezuka, Y.; Kadota, S. Recent progress in pharmacological research of propolis. Phyther. Res. 2001, 15, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Picolotto, A.; Pergher, D.; Pereira, G.P.; Machado, K.G.; da Silva Barud, H.; Roesch-Ely, M.; Gonzalez, M.H.; Tasso, L.; Figueiredo, J.G.; Moura, S. Bacterial cellulose membrane associated with red propolis as phytomodulator: Improved healing effects in experimental models of diabetes mellitus. Biomed. Pharmacother. 2019, 112, 108640. [Google Scholar] [CrossRef] [PubMed]
- Siheri, W.; Alenezi, S.; Tusiimire, J.; Watson, D.G. The chemical and biological properties of propolis. In Bee Products—Chemical and Biological Properties; Alvarez-Suarez, J.M., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 137–178. ISBN 978-3-319-59689-1. [Google Scholar]
- Granados-Pineda, J.; Uribe-Uribe, N.; García-López, P.; Ramos-Godinez, M.D.P.; Rivero-Cruz, J.F.; Pérez-Rojas, J.M. Effect of pinocembrin isolated from Mexican brown propolis on diabetic nephropathy. Molecules 2018, 23, 852. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.; Almeida, M.O.; Lemos, M.; Arruda, C.; Casoti, R.; Somensi, L.B.; Boeing, T.; Mariott, M.; Stein, B.D.P.; de Souza, P. Artepillin C, drupanin, aromadendrin-4′-O-methyl-ether and kaempferide from Brazilian green propolis promote gastroprotective action by diversified mode of action. J. Ethnopharmacol. 2018, 226, 82–89. [Google Scholar] [CrossRef]
- Rojczyk, E.; Klama-Baryła, A.; Łabuś, W.; Wilemska-Kucharzewska, K.; Kucharzewski, M. Historical and modern research on propolis and its application in wound healing and other fields of medicine and contributions by Polish studies. J. Ethnopharmacol. 2020, 262, 113159. [Google Scholar] [CrossRef] [PubMed]
- Governa, P.; Cusi, M.G.; Borgonetti, V.; Sforcin, J.M.; Terrosi, C.; Baini, G.; Miraldi, E.; Biagi, M. Beyond the biological effect of a chemically characterized poplar propolis: Antibacterial and antiviral activity and comparison with flurbiprofen in cytokines release by lps-stimulated human mononuclear cells. Biomedicines 2019, 7, 73. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.; Somensi, L.B.; Mariano, L.N.B.; Boeing, T.; Longo, B.; Perfoll, E.; de Souza, P.; Gushiken, L.F.S.; Pellizzon, C.H.; Rodrigues, D.M. Role of the antioxidant properties in the gastroprotective and gastric healing activity promoted by Brazilian green propolis and the healing efficacy of Artepillin C. Inflammopharmacology 2019, 28, 109–1025. [Google Scholar] [CrossRef] [PubMed]
- Tolba, M.F.; Omar, H.A.; Azab, S.S.; Khalifa, A.E.; Abdel-Naim, A.B.; Abdel-Rahman, S.Z. Caffeic acid phenethyl ester: A review of its antioxidant activity, protective effects against ischemia-reperfusion injury and drug adverse reactions. Crit. Rev. Food Sci. Nutr. 2016, 56, 2183–2190. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Dhanjal, J.K.; Bhargava, P.; Kaul, A.; Wang, J.; Zhang, H.; Kaul, S.C.; Wadhwa, R.; Sundar, D. Withanone and Withaferin-A are predicted to interact with transmembrane protease serine 2 (TMPRSS2) and block entry of SARS-CoV-2 into cells. J. Biomol. Struct. Dyn. 2020. [Google Scholar] [CrossRef]
- Anjum, S.I.; Ullah, A.; Khan, K.A.; Attaullah, M.; Khan, H.; Ali, H.; Bashir, M.A.; Tahir, M.; Ansari, M.J.; Ghramh, H.A. Composition and functional properties of propolis (bee glue): A review. Saudi J. Biol. Sci. 2019, 26, 1695–1703. [Google Scholar] [CrossRef]
- Zainullin, R.A.; Kunakova, R.V.; Gareev, V.F.; Galyautdinov, I.V.; Sadretdinova, Z.R.; Muslimov, Z.S.; Odinokov, V.N. flavanones and flavones from bashkir propolis. Chem. Nat. Compd. 2018, 54, 975–977. [Google Scholar] [CrossRef]
- Saito, É.; Sacoda, P.; Paviani, L.C.; Paula, J.T.; Cabral, F.A. Conventional and supercritical extraction of phenolic compounds from Brazilian red and green propolis. Sep. Sci. Technol. 2020. [Google Scholar] [CrossRef]
- Dudoit, A.; Mertz, C.; Chillet, M.; Cardinault, N.; Brat, P. Antifungal activity of Brazilian red propolis extract and isolation of bioactive fractions by thin-layer chromatography-bioautography. Food Chem. 2020, 327, 127060. [Google Scholar] [CrossRef]
- Farooqui, T.; Farooqui, A.A. Beneficial effects of propolis on human health and neurological diseases Tahira. Front. Biosci. 2012, 4, 779–793. [Google Scholar] [CrossRef]
- Oroian, M.; Dranca, F.; Ursachi, F. Comparative evaluation of maceration, microwave and ultrasonic-assisted extraction of phenolic compounds from propolis. J. Food Sci. Technol. 2020, 57, 70–78. [Google Scholar] [CrossRef]
- Berretta, A.A.; Silveira, M.A.D.; Cóndor Capcha, J.M.; De Jong, D. Propolis and its potential against SARS-CoV-2 infection mechanisms and COVID-19 disease: Running title: Propolis against SARS-CoV-2 infection and COVID-19. Biomed. Pharmacother. 2020, 131, 110622. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Lv, X.; Zhang, L.; Fu, X.; Song, S.; Su, A.; Chen, D.; Xu, L.; Wang, Y.; Wu, Z.; et al. Wogonin inhibits in vitro herpes simplex virus type 1 and 2 infection by modulating cellular NF-κB and MAPK pathways. BMC Microbiol. 2020, 20, 227. [Google Scholar] [CrossRef] [PubMed]
- Yildirim, A.; Duran, G.G.; Duran, N.; Jenedi, K.; Bolgul, B.S.; Miraloglu, M.; Muz, M. Antiviral activity of hatay propolis against replication of herpes simplex virus type 1 and type 2. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2016, 22, 422–430. [Google Scholar] [CrossRef]
- Mazia, R.S.; de Araújo Pereira, R.R.; de Francisco, L.M.B.; Natali, M.R.M.; Dias Filho, B.P.; Nakamura, C.V.; Bruschi, M.L.; Ueda-Nakamura, T. Formulation and evaluation of a mucoadhesive thermoresponsive system containing Brazilian green propolis for the treatment of lesions caused by herpes simplex type I. J. Pharm. Sci. 2016, 105, 113–121. [Google Scholar] [CrossRef]
- Sartori, G.; Pesarico, A.P.; Pinton, S.; Dobrachinski, F.; Roman, S.S.; Pauletto, F.; Rodrigues, L.C.; Prigol, M. Protective effect of brown Brazilian propolis against acute vaginal lesions caused by herpes simplex virus type 2 in mice: Involvement of antioxidant and anti-inflammatory mechanisms. Cell Biochem. Funct. 2012, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Nolkemper, S.; Reichling, J.; Sensch, K.H.; Schnitzler, P. Mechanism of herpes simplex virus type 2 suppression by propolis extracts. Phytomedicine 2010, 17, 132–138. [Google Scholar] [CrossRef]
- Bankova, V.; Galabov, A.S.; Antonova, D.; Vilhelmova, N.; Di Perri, B. Chemical composition of propolis extract ACF® and activity against herpes simplex virus. Phytomedicine 2014, 21, 1432–1438. [Google Scholar] [CrossRef]
- Búfalo, M.C.; Figueiredo, A.S.; De Sousa, J.P.B.; Candeias, J.M.G.; Bastos, J.K.; Sforcin, J.M. Anti-poliovirus activity of Baccharis dracunculifolia and propolis by cell viability determination and real-time PCR. J. Appl. Microbiol. 2009, 107, 1669–1680. [Google Scholar] [CrossRef]
- Labská, K.; Plodková, H.; Pumannová, M.; Sensch, K.H. Antiviral activity of propolis special extract GH 2002 against Varicella zoster virus in vitro. Die Pharm. Int. J. Pharm. Sci. 2018, 73, 733–736. [Google Scholar]
- Silva-Beltrán, N.P.; Balderrama-Carmona, A.P.; Umsza-Guez, M.A.; Souza Machado, B.A. Antiviral effects of Brazilian green and red propolis extracts on Enterovirus surrogates. Environ. Sci. Pollut. Res. 2020, 27, 28510–28517. [Google Scholar] [CrossRef] [PubMed]
- Gekker, G.; Hu, S.; Spivak, M.; Lokensgard, J.R.; Peterson, P.K. Anti-HIV-1 activity of propolis in CD4+ lymphocyte and microglial cell cultures. J. Ethnopharmacol. 2005, 102, 158–163. [Google Scholar] [CrossRef]
- Shimizu, T.; Hino, A.; Tsutsumi, A.; Park, Y.K.; Watanabe, W.; Kurokawa, M. Anti-influenza virus activity of propolis in vitro and its efficacy against influenza infection in mice. Antivir. Chem. Chemother. 2008, 19, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Búrquez, M.d.J.G.; Mosqueda, M.d.L.J.; Mendoza, H.R.; Zárate, C.I.S.; Miranda, L.C.; Sánchez, T.A.C. Protective effect of a Mexican propolis on MDBK cells exposed to aujeszky’s disease virus (pseudorabies virus). African J. Tradit. Complement. Altern. Med. 2015, 12, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Laila, U.; Akram, M.; Shariati, M.A.; Hashmi, A.M.; Akhtar, N.; Tahir, I.M.; Ghauri, A.O.; Munir, N.; Riaz, M.; Akhter, N.; et al. Role of medicinal plants in HIV/AIDS therapy. Clin. Exp. Pharmacol. Physiol. 2019, 46, 1063–1073. [Google Scholar] [CrossRef] [Green Version]
- Ito, J.; Chang, F.-R.; Wang, H.-K.; Park, Y.K.; Ikegaki, M.; Kilgore, N.; Lee, K.-H. Anti-AIDS agents. 48. Anti-HIV activity of moronic acid derivatives and the new melliferone-related triterpenoid isolated from Brazilian propolis. J. Nat. Prod. 2001, 64, 1278–1281. [Google Scholar] [CrossRef]
- Kai, H.; Obuchi, M.; Yoshida, H.; Watanabe, W.; Tsutsumi, S.; Park, Y.K.; Matsuno, K.; Yasukawa, K.; Kurokawa, M. In vitro and in vivo anti-influenza virus activities of flavonoids and related compounds as components of Brazilian propolis (AF-08). J. Funct. Foods 2014, 8, 214–223. [Google Scholar] [CrossRef]
- Erukhimovitch, V.; Talyshinsky, M.; Souprun, Y.; Huleihel, M. Spectroscopic investigation of herpes simplex viruses infected cells and their response to antiviral therapy. J. Mol. Struct. 2006, 792–793, 99–103. [Google Scholar] [CrossRef]
- Bachevski, D.; Damevska, K.; Simeonovski, V.; Dimova, M. Back to the basics: Propolis and COVID-19. Dermatol. Ther. 2020, 33, e13780. [Google Scholar] [CrossRef] [PubMed]
- Keni, R.; Alexander, A.; Nayak, P.G.; Mudgal, J.; Nandakumar, K. COVID-19: Emergence, spread, possible treatments, and global burden. Front. Public Health 2020, 8, 216–228. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Mohamed, B.S.; Elashal, M.H.; Du, M.; Guo, Z.; Zhao, C.; Musharraf, S.G.; Boskabady, M.H.; El-Seedi, H.H.R.; Efferth, T.; et al. Comprehensive overview on multiple strategies fighting COVID-19. Int. J. Environ. Res. Public Health 2020, 17, 5813. [Google Scholar] [CrossRef] [PubMed]
- Beigel, J.H.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S.; et al. Remdesivir for the treatment of Covid-19—Final report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Yosri, N.; El-Mallah, M.F.; Ghonaim, R.; Guo, Z.; Musharraf, S.G.; Du, M.; Khatib, A.; Xiao, J.; Saeed, A.; et al. Screening for natural and derived bio-active compounds in preclinical and clinical studies: One of the frontlines of fighting the coronaviruses pandemic. Phytomedicine 2020, 85, 153311–153333. [Google Scholar] [CrossRef]
- Shaldam, M.A.; Yahya, G.; Mohamed, N.H.; Abdel-Daim, M.M.; Al Naggar, Y. In silico screening of potent bioactive compounds from honey bee products against COVID-19 target enzymes. ChemRxiv 2020, 12644102, 1–18. [Google Scholar]
- Cheng, P.C.; Wong, G. Honey bee propolis: Prospects in medicine. Bee World 1996, 77, 8–15. [Google Scholar] [CrossRef]
- Lima, W.G.; Brito, J.C.M.; da Cruz Nizer, W.S. Bee products as a source of promising therapeutic and chemoprophylaxis strategies against COVID-19 (SARS-CoV-2). Phyther. Res. 2020, 35, 743–750. [Google Scholar] [CrossRef]
- Maruta, H.; He, H. PAK1-blockers: Potential therapeutics against COVID-19. Med. Drug Discov. 2020, 6, 100039. [Google Scholar] [CrossRef]
- Chiow, K.H.; Phoon, M.C.; Putti, T.; Tan, B.K.H.; Chow, V.T. Evaluation of antiviral activities of Houttuynia cordata Thunb. extract, quercetin, quercetrin and cinanserin on murine coronavirus and dengue virus infection. Asian Pac. J. Trop. Med. 2016, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Colunga Biancatelli, R.M.L.; Berrill, M.; Catravas, J.D.; Marik, P.E. Quercetin and vitamin c: An experimental, synergistic therapy for the prevention and treatment of sars-cov-2 related disease (COVID-19). Front. Immunol. 2020, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Sforcin, J.M. Propolis and the immune system: A review. J. Ethnopharmacol. 2007, 113, 1–14. [Google Scholar] [CrossRef]
- Piñeros, A.R.; de Lima, M.H.F.; Rodrigues, T.; Gembre, A.F.; Bertolini, T.B.; Fonseca, M.D.; Berretta, A.A.; Ramalho, L.N.Z.; Cunha, F.Q.; Hori, J.I.; et al. Green propolis increases myeloid suppressor cells and CD4+Foxp3+ cells and reduces Th2 inflammation in the lungs after allergen exposure. J. Ethnopharmacol. 2020, 252, 112496. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. Asthma: The importance of dysregulated barrier immunity. Eur. J. Immunol. 2013, 43, 3125–3137. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.S.; Hamid, Q.; Ying, S.; Tsicopoulos, A.; Barkans, J.; Bentley, A.M.; Corrigan, C.; Durham, S.R.; Kay, A.B. Predominant TH2-like bronchoalveolar T-lymphocyte population in atopic asthma. N. Engl. J. Med. 1992, 326, 298–304. [Google Scholar] [CrossRef]
- Sy, L.B.; Wu, Y.-L.; Chiang, B.-L.; Wang, Y.-H.; Wu, W.-M. Propolis extracts exhibit an immunoregulatory activity in an OVA-sensitized airway inflammatory animal model. Int. Immunopharmacol. 2006, 6, 1053–1060. [Google Scholar] [CrossRef]
- Athari, S.S. Targeting cell signaling in allergic asthma. Signal Transduct. Target. Ther. 2019, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Okamoto, Y.; Fukui, T.; Masuzawa, T. Suppression of interleukin 17 production by Brazilian propolis in mice with collagen-induced arthritis. Inflammopharmacology 2012, 20, 19–26. [Google Scholar] [CrossRef]
- Van den Berg, W.B.; Miossec, P. IL-17 as a future therapeutic target for rheumatoid arthritis. Nat. Rev. Rheumatol. 2009, 5, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Roeleveld, D.M.; Koenders, M.I. The role of the Th17 cytokines IL-17 and IL-22 in Rheumatoid Arthritis pathogenesis and developments in cytokine immunotherapy. Cytokine 2015, 74, 101–107. [Google Scholar] [CrossRef]
- Okamoto, Y.; Tanaka, M.; Fukui, T.; Masuzawa, T. Brazilian propolis inhibits the differentiation of Th17 cells by inhibition of interleukin-6-induced phosphorylation of signal transducer and activator of transcription 3. Immunopharmacol. Immunotoxicol. 2012, 34, 803–809. [Google Scholar] [CrossRef]
- Choi, J.-H.; Roh, K.-H.; Oh, H.; Park, S.-J.; Ha, S.-M.; Kang, M.S.; Lee, J.-H.; Jung, S.Y.; Song, H.; Yang, J.W.; et al. Caffeic acid phenethyl ester lessens disease symptoms in an experimental autoimmune uveoretinitis mouse model. Exp. Eye Res. 2015, 134, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Takeda, K.; Kaisho, T. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2001, 2, 675–680. [Google Scholar] [CrossRef]
- Orsatti, C.L.; Sforcin, J.M. Propolis immunomodulatory activity on TLR-2 and TLR-4 expression by chronically stressed mice. Nat. Prod. Res. 2012, 26, 446–453. [Google Scholar] [CrossRef]
- Búfalo, M.C.; Bordon-Graciani, A.P.; Conti, B.J.; de Assis Golim, M.; Sforcin, J.M. The immunomodulatory effect of propolis on receptors expression, cytokine production and fungicidal activity of human monocytes. J. Pharm. Pharmacol. 2014, 66, 1497–1504. [Google Scholar] [CrossRef] [PubMed]
- Conti, B.J.; Búfalo, M.C.; Golim, M.d.A.; Bankova, V.; Sforcin, J.M. Cinnamic acid is partially involved in propolis immunomodulatory action on human monocytes. Evid. Based Complement. Altern. Med. 2013, 2013, 109864. [Google Scholar] [CrossRef]
- Orsatti, C.L.; Missima, F.; Pagliarone, A.C.; Bachiega, T.F.; Búfalo, M.C.; Araújo, J.P., Jr.; Sforcin, J.M. Propolis immunomodulatory action in vivo on Toll-like receptors 2 and 4 expression and on pro-inflammatory cytokines production in mice. Phyther. Res. 2010, 24, 1141–1146. [Google Scholar] [CrossRef]
- Conti, B.J.; Santiago, K.B.; Cardoso, E.O.; Freire, P.P.; Carvalho, R.F.; Golim, M.A.; Sforcin, J.M. Propolis modulates miRNAs involved in TLR-4 pathway, NF-κB activation, cytokine production and in the bactericidal activity of human dendritic cells. J. Pharm. Pharmacol. 2016, 68, 1604–1612. [Google Scholar] [CrossRef]
- Búfalo, M.C.; Sforcin, J.M. The modulatory effects of caffeic acid on human monocytes and its involvement in propolis action. J. Pharm. Pharmacol. 2015, 67, 740–745. [Google Scholar] [CrossRef]
- Pontin, K.; Da Silva Filho, A.A.; Santos, F.F.; e Silva, M.L.A.; Cunha, W.R.; Nanayakkara, N.P.D.; Bastos, J.K.; de Albuquerque, S. In vitro and in vivo antileishmanial activities of a Brazilian green propolis extract. Parasitol. Res. 2008, 103, 487–492. [Google Scholar] [CrossRef]
- Da Silva, S.S.; Thomé, G.d.S.; Cataneo, A.H.D.; Miranda, M.M.; Felipe, I.; Andrade, C.G.T.d.J.; Watanabe, M.A.E.; Piana, G.M.; Sforcin, J.M.; Pavanelli, W.R.; et al. Brazilian propolis antileishmanial and immunomodulatory effects. Evidence-Based Complement. Altern. Med. 2013, 2013, 673058. [Google Scholar] [CrossRef] [Green Version]
- Amarante, M.K.; Watanabe, M.A.E.; Conchon-Costa, I.; Fiori, L.L.; Oda, J.M.M.; Búfalo, M.C.; Sforcin, J.M. The effect of propolis on CCL5 and IFN-γ expression by peripheral blood mononuclear cells from leishmaniasis patients. J. Pharm. Pharmacol. 2012, 64, 154–160. [Google Scholar] [CrossRef]
- Dos Santos Thomazelli, A.P.F.; Tomiotto-Pellissier, F.; da Silva, S.S.; Panis, C.; Orsini, T.M.; Cataneo, A.H.D.; Miranda-Sapla, M.M.; Custódio, L.A.; Tatakihara, V.L.H.; Bordignon, J.; et al. Brazilian propolis promotes immunomodulation on human cells from American Tegumentar Leishmaniasis patients and healthy donors infected with L. braziliensis. Cell. Immunol. 2017, 311, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Benkovic, V.; Horvat Knezevic, A.; Brozovic, G.; Knezevic, F.; Đikic, D.; Bevanda, M.; Basic, I.; Orsolic, N. Enhanced antitumor activity of irinotecan combined with propolis and its polyphenolic compounds on Ehrlich ascites tumor in mice. Biomed. Pharmacother. 2007, 61, 292–297. [Google Scholar] [CrossRef]
- Oršolić, N.; Benković, V.; Lisičić, D.; Đikić, D.; Erhardt, J.; Horvat Knežević, A. Protective effects of propolis and related polyphenolic/flavonoid compounds against toxicity induced by irinotecan. Med. Oncol. 2010, 27, 1346–1358. [Google Scholar] [CrossRef]
- Oršolić, N.; Bašić, I. Immunomodulation by water-soluble derivative of propolis: A factor of antitumor reactivity. J. Ethnopharmacol. 2003, 84, 265–273. [Google Scholar] [CrossRef]
- Oršolić, N.; Bašić, I. Water-soluble derivative of propolis and its polyphenolic compounds enhance tumoricidal activity of macrophages. J. Ethnopharmacol. 2005, 102, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Oršolic, N.; Terzic, S.; Šver, L.; Bašic, I. Polyphenolic compounds from propolis modulate immune responses and increase host resistance to tumour cells. Food Agric. Immunol. 2005, 16, 165–179. [Google Scholar] [CrossRef]
- Oršolic, N.; Šver, L.; Terzić, S.; Bašić, I. Peroral application of water-soluble derivative of propolis (WADP) and its related polyphenolic compounds and their influence on immunological and antitumour activity. Vet. Res. Commun. 2005, 29, 575–593. [Google Scholar] [CrossRef]
- Fischer, G.; Conceição, F.R.; Leite, F.P.L.; Dummer, L.A.; Vargas, G.D.; de Hübner, S.O.; Dellagostin, O.A.; Paulino, N.; Paulino, A.S.; Vidor, T. Immunomodulation produced by a green propolis extract on humoral and cellular responses of mice immunized with SuHV-1. Vaccine 2007, 25, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.C.-F.; Cheung, K.-W.; Sze, D.M.-Y. The immunomodulatory and anticancer properties of propolis. Clin. Rev. Allergy Immunol. 2013, 44, 262–273. [Google Scholar] [CrossRef]
- Medjeber, O.; Touri, K.; Rafa, H.; Djeraba, Z.; Belkhelfa, M.; Boutaleb, A.F.; Arroul-Lammali, A.; Belguendouz, H.; Touil-Boukoffa, C. Ex vivo immunomodulatory effect of ethanolic extract of propolis during Celiac Disease: Involvement of nitric oxide pathway. Inflammopharmacology 2018, 26, 1469–1481. [Google Scholar] [CrossRef]
- Touzani, S.; Embaslat, W.; Imtara, H.; Kmail, A.; Kadan, S.; Zaid, H.; ElArabi, I.; Badiaa, L.; Saad, B. In vitro evaluation of the potential use of propolis as a multitarget therapeutic product: Physicochemical properties, chemical composition, and immunomodulatory, antibacterial, and anticancer properties. Biomed Res. Int. 2019, 2019, 4836378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosravi, A.R.; Shokri, H.; Darvishi, S.; Taghavi, M. Immunomodulatory efficacy of ethanol extract of propolis on tumor-bearing mice with disseminated candidiasis. J. Mycol. Med. 2014, 24, e143–e148. [Google Scholar] [CrossRef]
- Asgharpour, F.; Moghadamnia, A.A.; Motallebnejad, M.; Nouri, H.R. Propolis attenuates lipopolysaccharide-induced inflammatory responses through intracellular ROS and NO levels along with downregulation of IL-1β and IL-6 expressions in murine RAW 264.7 macrophages. J. Food Biochem. 2019, 43, e12926. [Google Scholar] [CrossRef]
- Conti, B.J.; Santiago, K.B.; Búfalo, M.C.; Herrera, Y.F.; Alday, E.; Velazquez, C.; Hernandez, J.; Sforcin, J.M. Modulatory effects of propolis samples from Latin America (Brazil, Cuba and Mexico) on cytokine production by human monocytes. J. Pharm. Pharmacol. 2015, 67, 1431–1438. [Google Scholar] [CrossRef]
- Ivanovska, N.D.; Dimov, V.B.; Bankova, V.S.; Popov, S.S. Immunomodulatory action of propolis. VI. Influence of a water soluble derivative on complement activity in vivo. J. Ethnopharmacol. 1995, 47, 145–147. [Google Scholar] [CrossRef]
- Girgin, G.; Baydar, T.; Ledochowski, M.; Schennach, H.; Bolukbasi, D.N.; Sorkun, K.; Salih, B.; Sahin, G.; Fuchs, D. Immunomodulatory effects of Turkish propolis: Changes in neopterin release and tryptophan degradation. Immunobiology 2009, 214, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Bachiega, T.F.; Orsatti, C.L.; Pagliarone, A.C.; Sforcin, J.M. The effects of propolis and its isolated compounds on cytokine production by murine macrophages. Phyther. Res. 2012, 26, 1308–1313. [Google Scholar] [CrossRef]
- Búfalo, M.C.; Ferreira, I.; Costa, G.; Francisco, V.; Liberal, J.; Cruz, M.T.; Lopes, M.C.; Batista, M.T.; Sforcin, J.M. Propolis and its constituent caffeic acid suppress LPS-stimulated pro-inflammatory response by blocking NF-κB and MAPK activation in macrophages. J. Ethnopharmacol. 2013, 149, 84–92. [Google Scholar] [CrossRef]
- Machado, J.L.; Assunção, A.K.M.; da Silva, M.C.P.; dos Reis, A.S.; Costa, G.C.; Arruda, D.d.S.; Rocha, B.A.; Vaz, M.M.d.O.L.L.; Paes, A.M.d.A.; Guerra, R.N.M.; et al. Brazilian green propolis: Anti-inflammatory property by an immunomodulatory activity. Evidence-Based Complement. Altern. Med. 2012, 2012, 157652. [Google Scholar] [CrossRef] [Green Version]
- Pagliarone, A.C.; Missima, F.; Orsatti, C.L.; Bachiega, T.F.; Sforcin, J.M. Propolis effect on Th1/Th2 cytokines production by acutely stressed mice. J. Ethnopharmacol. 2009, 125, 230–233. [Google Scholar] [CrossRef]
- Missima, F.; Pagliarone, A.C.; Orsatti, C.L.; Araújo Jr, J.P.; Sforcin, J.M. The Effect of propolis on Th1/Th2 cytokine expression and production by melanoma-bearing mice submitted to stress. Phyther. Res. 2010, 24, 1501–1507. [Google Scholar] [CrossRef]
- Pagliarone, A.C.; Orsatti, C.L.; Búfalo, M.C.; Missima, F.; Bachiega, T.F.; Júnior, J.P.A.; Sforcin, J.M. Propolis effects on pro-inflammatory cytokine production and Toll-like receptor 2 and 4 expression in stressed mice. Int. Immunopharmacol. 2009, 9, 1352–1356. [Google Scholar] [CrossRef]
- Orsatti, C.L.; Missima, F.; Pagliarone, A.C.; Sforcin, J.M. Th1/Th2 cytokines’ expression and production by propolis-treated mice. J. Ethnopharmacol. 2010, 129, 314–318. [Google Scholar] [CrossRef]
- Dimov, V.; Ivanovska, N.; Bankova, V.; Popov, S. Immunomodulatory action of propolis: IV. Prophylactic activity against Gram-negative infections and adjuvant effect of the water-soluble derivative. Vaccine 1992, 10, 817–823. [Google Scholar] [CrossRef]
- Oršolić, N.; Knežević, A.H.; Šver, L.; Terzić, S.; Bašić, I. Immunomodulatory and antimetastatic action of propolis and related polyphenolic compounds. J. Ethnopharmacol. 2004, 94, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Sá-Nunes, A.; Faccioli, L.H.; Sforcin, J.M. Propolis: Lymphocyte proliferation and IFN-γ production. J. Ethnopharmacol. 2003, 87, 93–97. [Google Scholar] [CrossRef]
- Oršolić, N.; Šaranović, A.B.; Bašić, I. Direct and indirect mechanism(s) of antitumour activity of propolis and its polyphenolic compounds. Planta Med 2006, 72, 20–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Missima, F.; Sforcin, J.M. Green brazilian propolis action on macrophages and lymphoid organs of chronically stressed mice. Evid. Based Complement. Altern. Med. 2008, 5, 973715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimov, V.; Ivanovska, N.; Manolova, N.; Bankova, V.; Nikolov, N.; Popov, S. Immunomodulatory action of propolis. Influence on anti-infectious protection and macrophage function. Apidologie 1991, 22, 155–162. [Google Scholar] [CrossRef]
- Orsolic, N.; Sver, L.; Terzic, S.; Tadic, Z.; Basic, I. Inhibitory effect of water-soluble derivative of propolis and its polyphenolic compounds on tumor growth and metastasizing ability: A possible mode of antitumor action. Nutr. Cancer 2003, 47, 156–163. [Google Scholar] [CrossRef]
- Dantas, A.P.; Olivieri, B.P.; Gomes, F.H.M.; De Castro, S.L. Treatment of Trypanosoma cruzi-infected mice with propolis promotes changes in the immune response. J. Ethnopharmacol. 2006, 103, 187–193. [Google Scholar] [CrossRef]
- Münstedt, K. Bee products and the treatment of blister-like lesions around the mouth, skin and genitalia caused by herpes viruses—A systematic review. Complement. Ther. Med. 2019, 43, 81–84. [Google Scholar] [CrossRef]
- Vynograd, N.; Vynograd, I.; Sosnowski, Z. A comparative multi-centre study of the efficacy of propolis, acyclovir and placebo in the treatment of genital herpes (HSV). Phytomedicine 2000, 7, 1–6. [Google Scholar] [CrossRef]
- Tomanova, D.; Holcova, S.; Hladikova, M. Clinical study: Lotion containing propolis special extract GH 2002 0.5% vs. placebo as on-top treatment of herpes zoster. Health (Irvine. Calif) 2017, 9, 1337–1347. [Google Scholar] [CrossRef] [Green Version]
- Adis Insight. GH 2002; Propolis Extract; Propolis Special Extract. Available online: https://adisinsight.springer.com (accessed on 20 June 2021).
- Holcová, S.; Hladiková, M. Efficacy and tolerability of propolis special extract GH 2002 as a lip balm against herpes labialis: A randomized, double-blind three-arm dose finding study. Health (Irvine. Calif) 2011, 3, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Arenberger, P.; Arenbergerova, M.; Hladíková, M.; Holcova, S.; Ottillinger, B. Comparative study with a lip balm containing 0.5% Propolis special extract GH 2002 versus 5% aciclovir cream in patients with herpes labialis in the papular/erythematous stage: A single-blind, randomized, two-arm study. Curr. Ther. Res. 2018, 88, 1–7. [Google Scholar] [CrossRef]
- Jautová, J.; Zelenková, H.; Drotarová, K.; Nejdková, A.; Grünwaldová, B.; Hladiková, M. Lip creams with propolis special extract GH 2002 0.5% versus aciclovir 5.0% for herpes labialis (vesicular stage). Wien. Med. Wochenschr. 2019, 169, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Scully, C. Propolis: A background. Br. Dent. J. 2006, 200, 359–360. [Google Scholar] [CrossRef] [PubMed]
- Kandeel, M.; Abdelrahman, A.H.M.; Oh-Hashi, K.; Ibrahim, A.; Venugopala, K.N.; Morsy, M.A.; Ibrahim, M.A.A. Repurposing of FDA-approved antivirals, antibiotics, anthelmintics, antioxidants, and cell protectives against SARS-CoV-2 papain-like protease. J. Biomol. Struct. Dyn. 2020, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, T.A.; Elshamy, A.I.; Ibrahim, M.A.A.; Zellagui, A.; Moustafa, M.F.; Abdelrahman, A.H.M.; Ohta, S.; Pare, P.W.; Hegazy, M.-E.F. Carotane sesquiterpenes from Ferula vesceritensis: In silico analysis as SARS-CoV-2 binding inhibitors. RSC Adv. 2020, 10, 34541–34548. [Google Scholar] [CrossRef]
- Ibrahim, M.A.A.; Mohamed, E.A.R.; Abdelrahman, A.H.M.; Allemailem, K.S.; Moustafa, M.F.; Shawky, A.M.; Mahzari, A.; Hakami, A.R.; Abdeljawaad, K.A.A.; Atia, M.A.M. Rutin and flavone analogs as prospective SARS-CoV-2 main protease inhibitors: In silico drug discovery study. J. Mol. Graph. Model. 2021, 105, 107904. [Google Scholar] [CrossRef]
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Hussien, T.A.; Badr, E.A.A.; Mohamed, T.A.; El-Seedi, H.R.; Pare, P.W.; Efferth, T.; Hegazy, M.-E.F. In silico drug discovery of major metabolites from spices as SARS-CoV-2 main protease inhibitors. Comput. Biol. Med. 2020, 126, 104046. [Google Scholar] [CrossRef]
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Hegazy, M.-E.F. In-silico Drug Repurposing and Molecular Dynamics Puzzled out Potential SARS-CoV-2 Main Protease Inhibitors. Available online: https://www.tandfonline.com/doi/full/10.1080/07391102.2020.1791958 (accessed on 29 July 2021).
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Allemailem, K.S.; Almatroudi, A.; Moustafa, M.F.; Hegazy, M.-E.F. In silico evaluation of prospective anti-Covid-19 drug candidates as potential SARS-COV-2 main protease inhibito. Protein J. 2021, 40, 296–309. [Google Scholar] [CrossRef]
- Ibrahim, M.A.A.; Abdeljawaad, K.A.A.; Abdelrahman, A.H.M.; Hegazy, M.-E.F. Natural-like products as potential SARS-CoV-2 Mpro inhibitors: In-silico drug discovery. J. Biomol. Struct. Dyn. 2020, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.A.; Abdelrahman, A.H.M.; Mohamed, T.A.; Atia, M.A.M.; Al-Hammady, M.A.M.; Abdeljawaad, K.A.A.; Elkady, E.M.; Moustafa, M.F.; Alrumaihi, F.; Allemailem, K.S.; et al. In silico mining of terpenes from red-sea invertebrates for sars-COV-2 main protease (Mpro) inhibitors. Molecules 2021, 26, 2082. [Google Scholar] [CrossRef]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Yan, L.; Huang, Y.; Liu, F.; Zhao, Y.; Cao, L.; Wang, T.; Sun, Q.; Ming, Z.; Zhang, L.; et al. Structure of the RNA-dependent RNA polymerase from COVID-19 virus. Science (80-) 2020, 368, 779–782. [Google Scholar] [CrossRef] [Green Version]
- Osipiuk, J.; Azizi, S.-A.; Dvorkin, S.; Endres, M.; Jedrzejczak, R.; Jones, K.A.; Kang, S.; Kathayat, R.S.; Kim, Y.; Lisnyak, V.G.; et al. Structure of papain-like protease from SARS-CoV-2 and its complexes with non-covalent inhibitors. Nat. Commun. 2021, 12, 743. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Newman, J.A.; Douangamath, A.; Yazdani, S.; Yosaatmadja, Y.; Aimon, A.; Brandão-Neto, J.; Dunnett, L.; Gorrie-stone, T.; Skyner, R.; Fearon, D.; et al. Structure, Mechanism and Crystallographic Fragment Screening of the SARS-COV-2 NSP13 Helicase. bioRxiv 2021. Available online: https://www.biorxiv.org/content/10.1101/2021.03.15.435326v1.abstract (accessed on 29 July 2021).
- Martí-Renom, M.A.; Stuart, A.C.; Fiser, A.; Sánchez, R.; Melo, F.; Šali, A. Comparative protein structure modeling of genes and genomes. Annu. Rev. Biophys. Biomol. Struct. 2000, 29, 291–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordon, J.C.; Myers, J.B.; Folta, T.; Shoja, V.; Heath, L.S.; Onufriev, A. H++: A server for estimating p Ka s and adding missing hydrogens to macromolecules. Nucleic Acids Res. 2005, 33, W368–W371. [Google Scholar] [CrossRef]
- Forli, S.; Huey, R.; Pique, M.E.; Sanner, M.F.; Goodsell, D.S.; Olson, A.J. Computational protein–ligand docking and virtual drug screening with the AutoDock suite. Nat. Protoc. 2016, 11, 905–919. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, P.C.D.; Skillman, A.G.; Warren, G.L.; Ellingson, B.A.; Stahl, M.T. Conformer generation with OMEGA: Algorithm and validation using high quality structures from the protein databank and cambridge structural database. J. Chem. Inf. Model. 2010, 50, 572–584. [Google Scholar] [CrossRef]
- OMEGA 2.5.1.4 OpenEye Scientific Software. Available online: https://www.yumpu.com/en/document/view/19633413/omega-release-2514-openeye-scientific-software-inc (accessed on 29 July 2021).
- Halgren, T.A. MMFF VI. MMFF94s option for energy minimization studies. J. Comput. Chem. 1999, 20, 720–729. [Google Scholar] [CrossRef]
- SZYBKI 1.9.0.3; OpenEye Scientific Software: Santa Fe, NM, USA, 2016.
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, J.; Marsili, M. Iterative partial equalization of orbital electronegativity—A rapid access to atomic charges. Tetrahedron 1980, 36, 3219–3228. [Google Scholar] [CrossRef]
- Cheng, F.; Li, W.; Zhou, Y.; Shen, J.; Wu, Z.; Liu, G.; Lee, P.W.; Tang, Y. AdmetSAR: A comprehensive source and free tool for assessment of chemical ADMET properties. J. Chem. Inf. Model. 2012, 52, 3099–3105. [Google Scholar] [CrossRef] [PubMed]









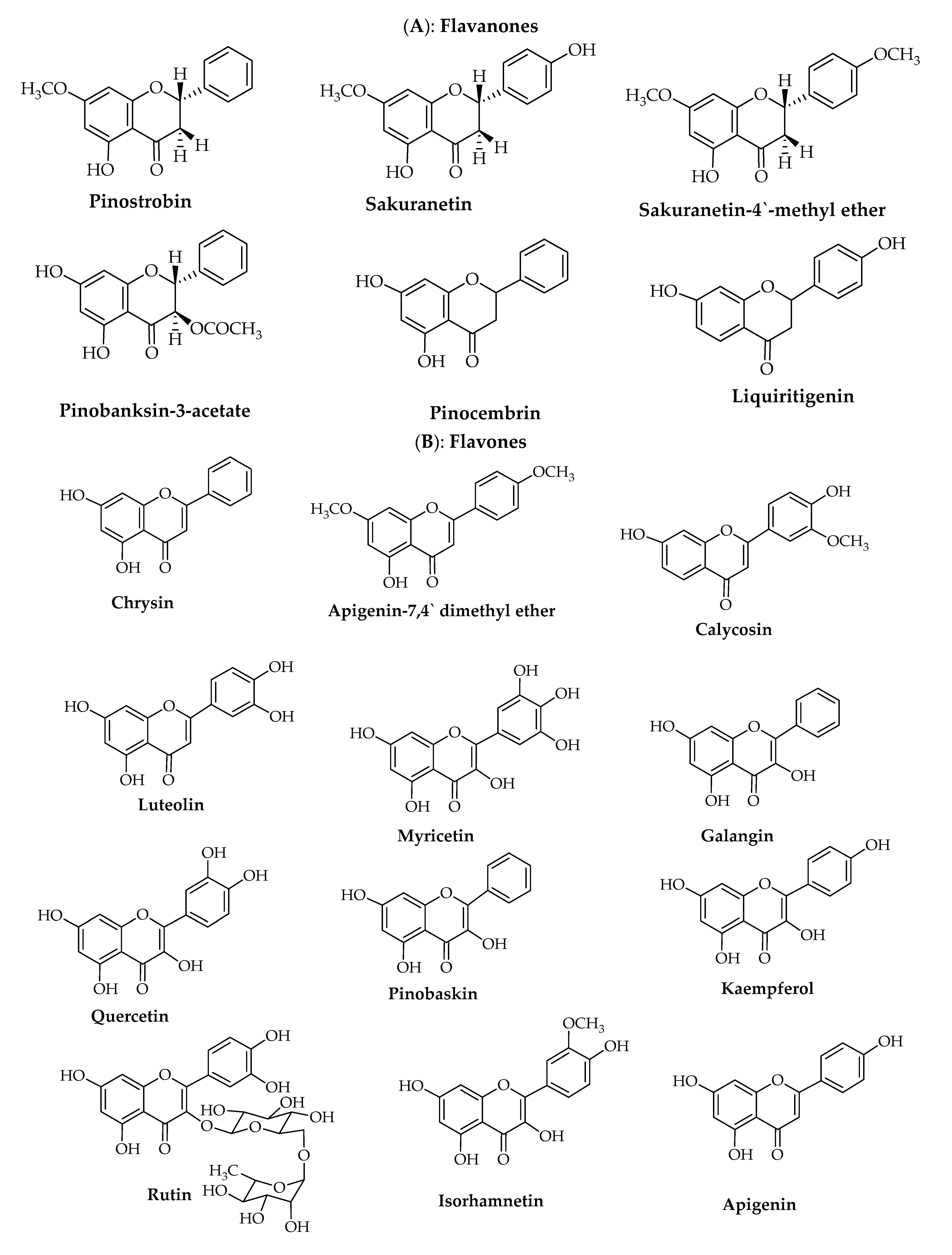{kind=link}
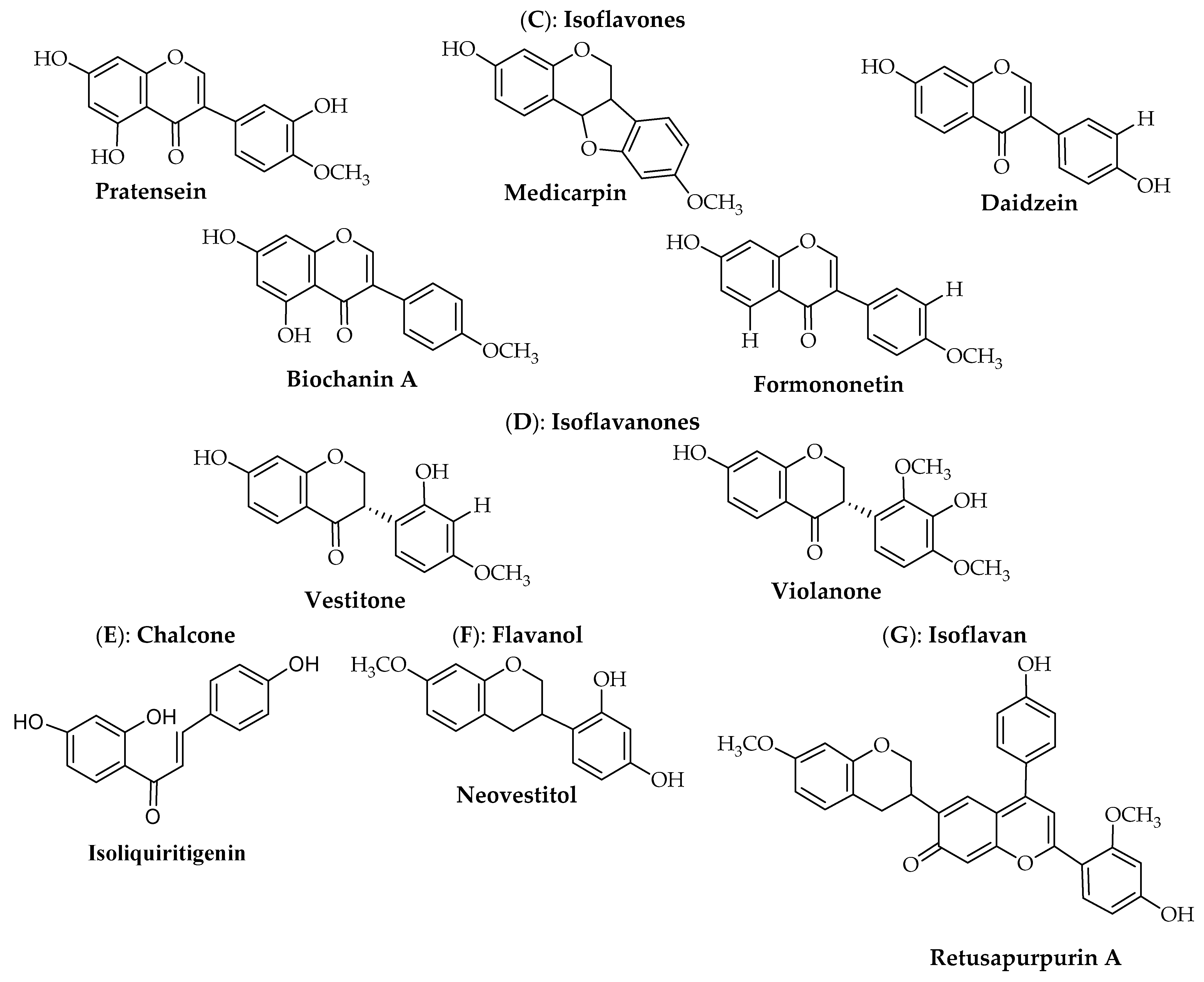{kind=link}
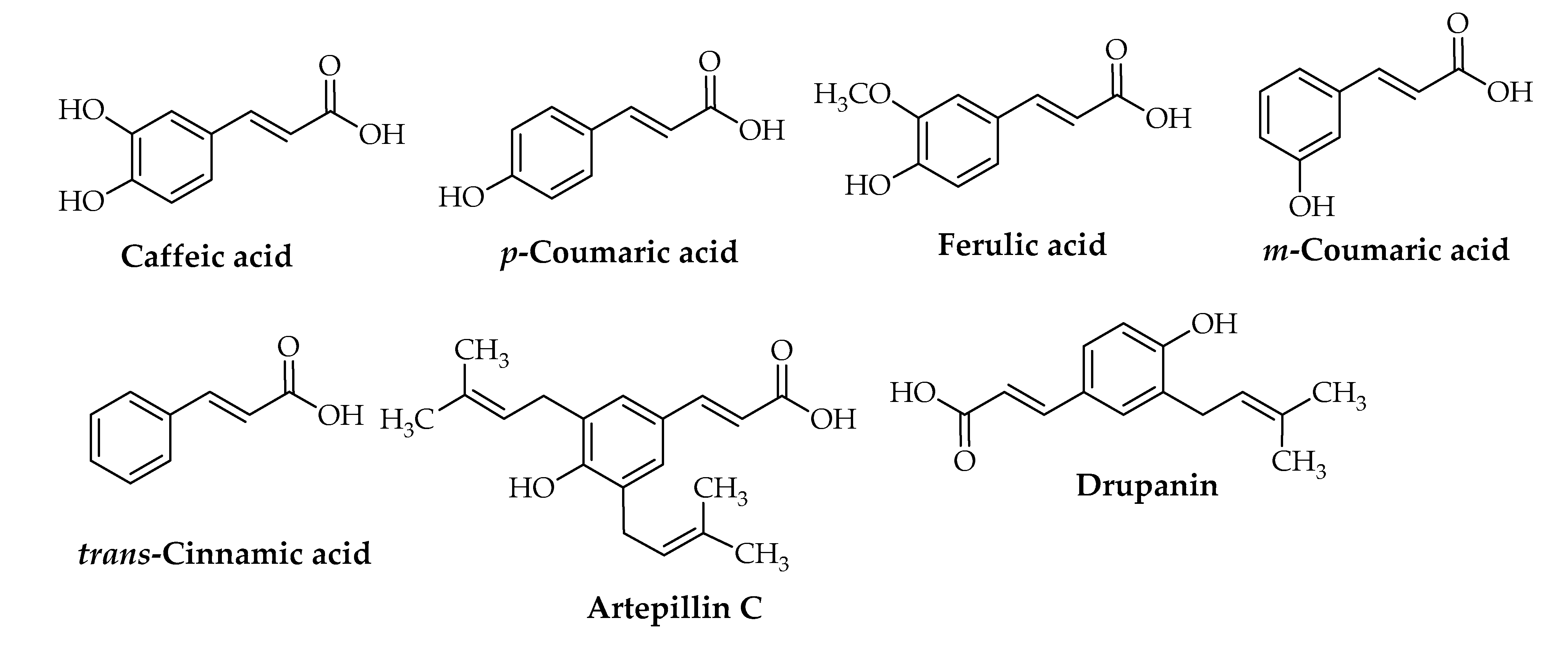{kind=link}
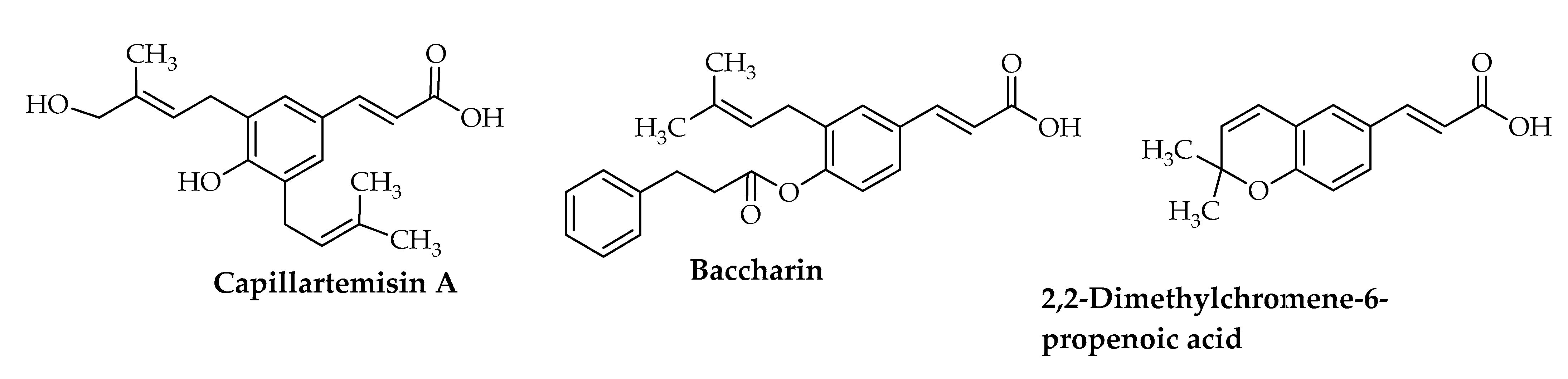{kind=link}
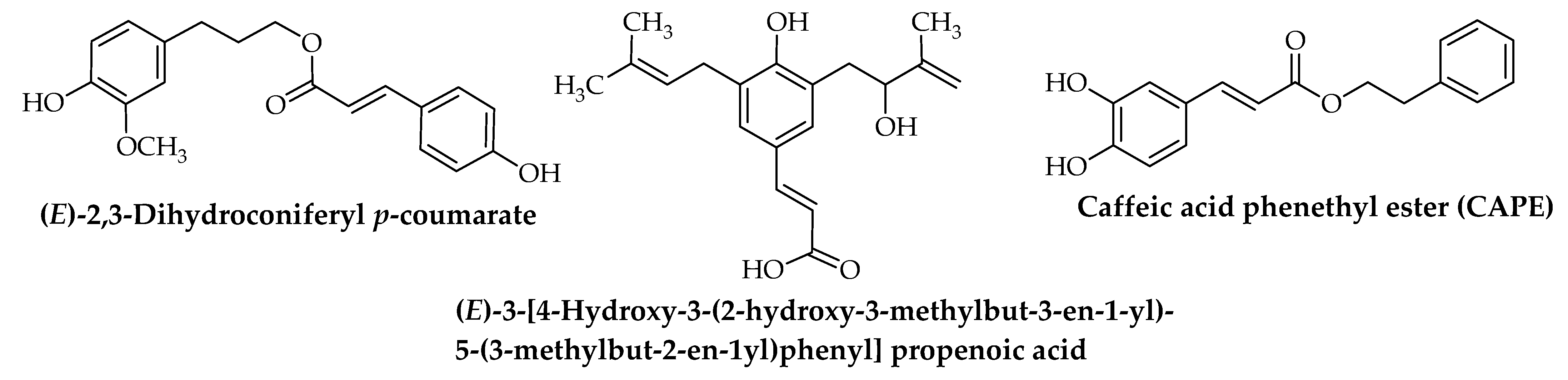{kind=link}
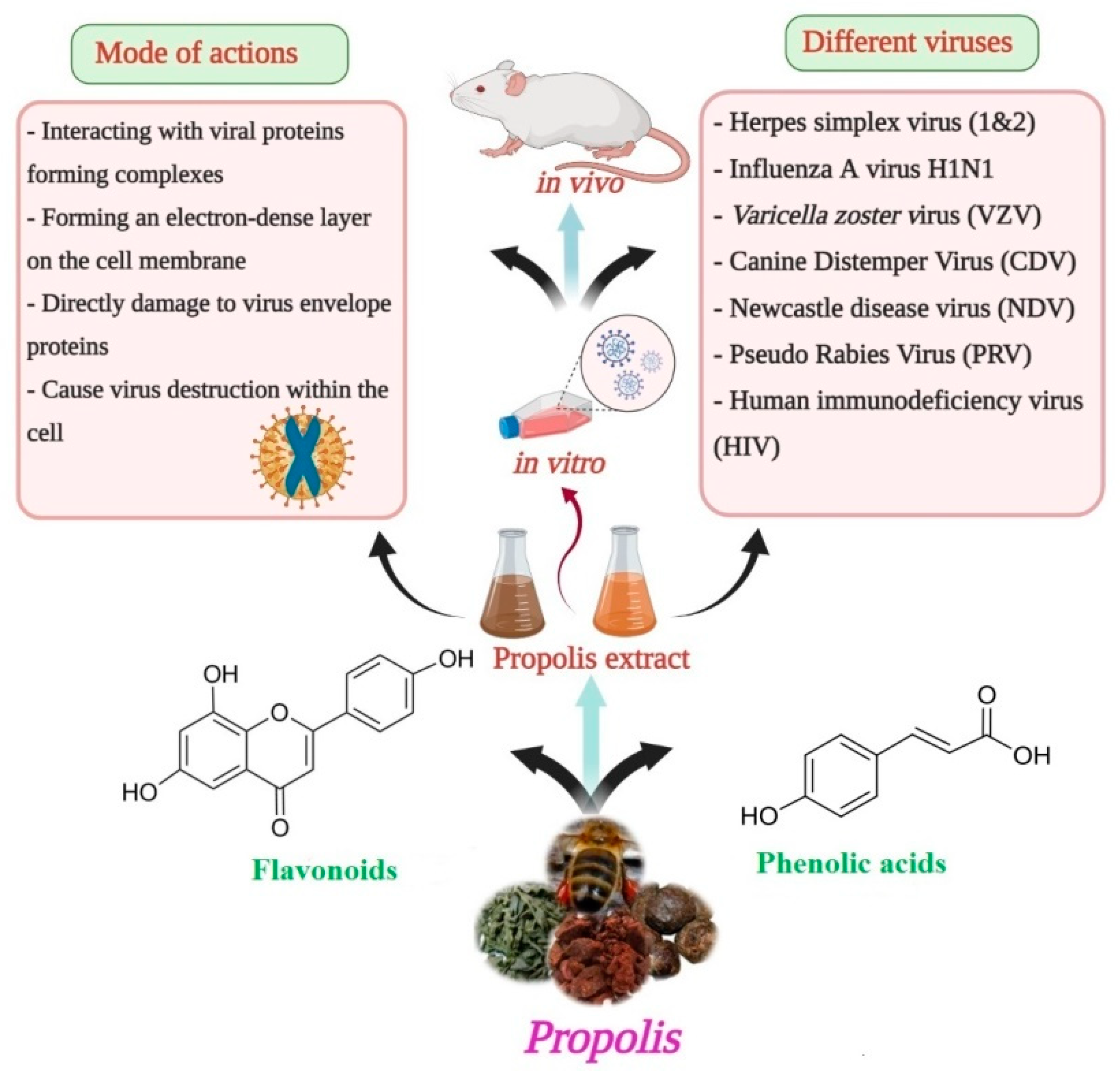{kind=link}
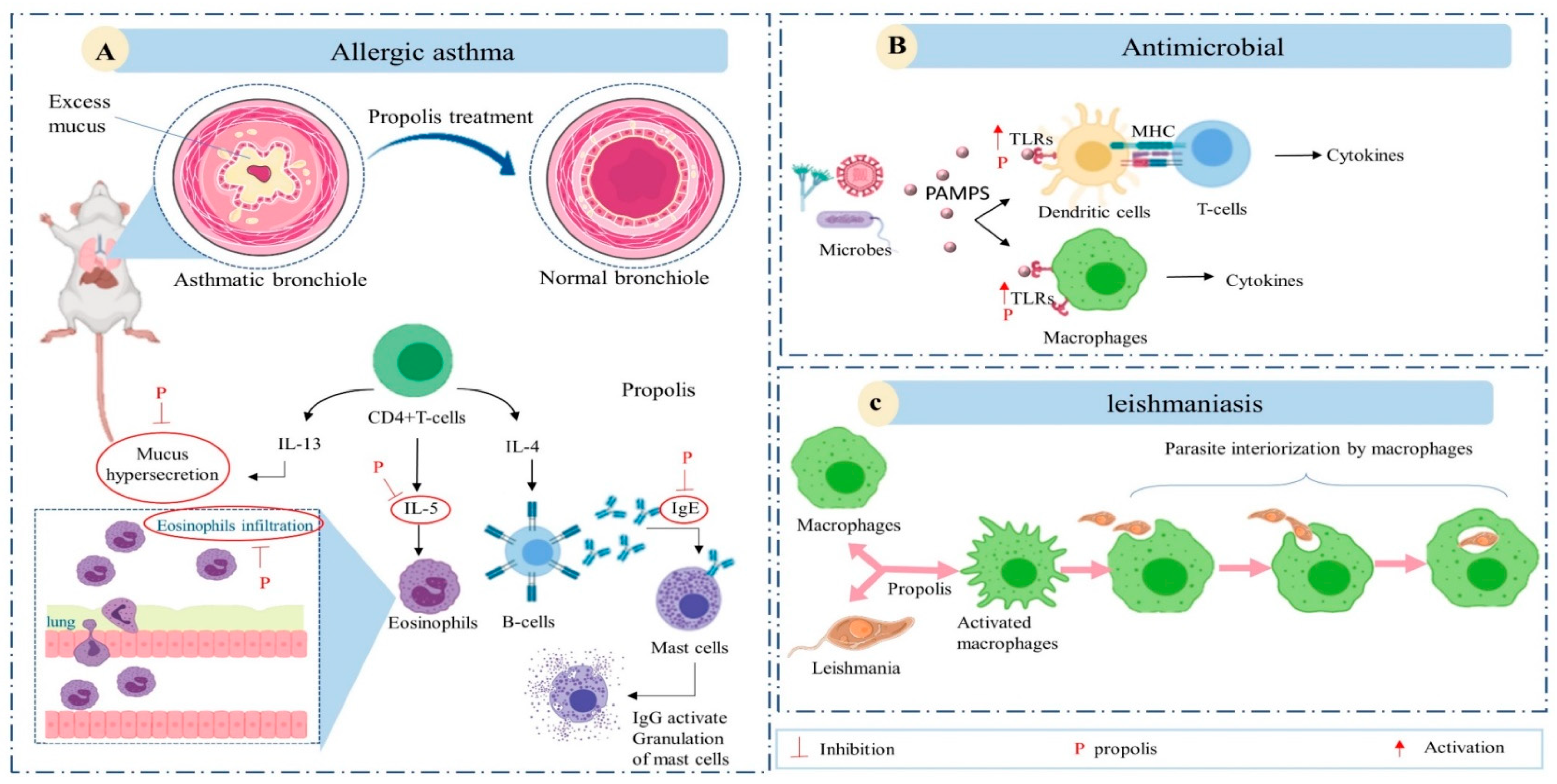{kind=link}
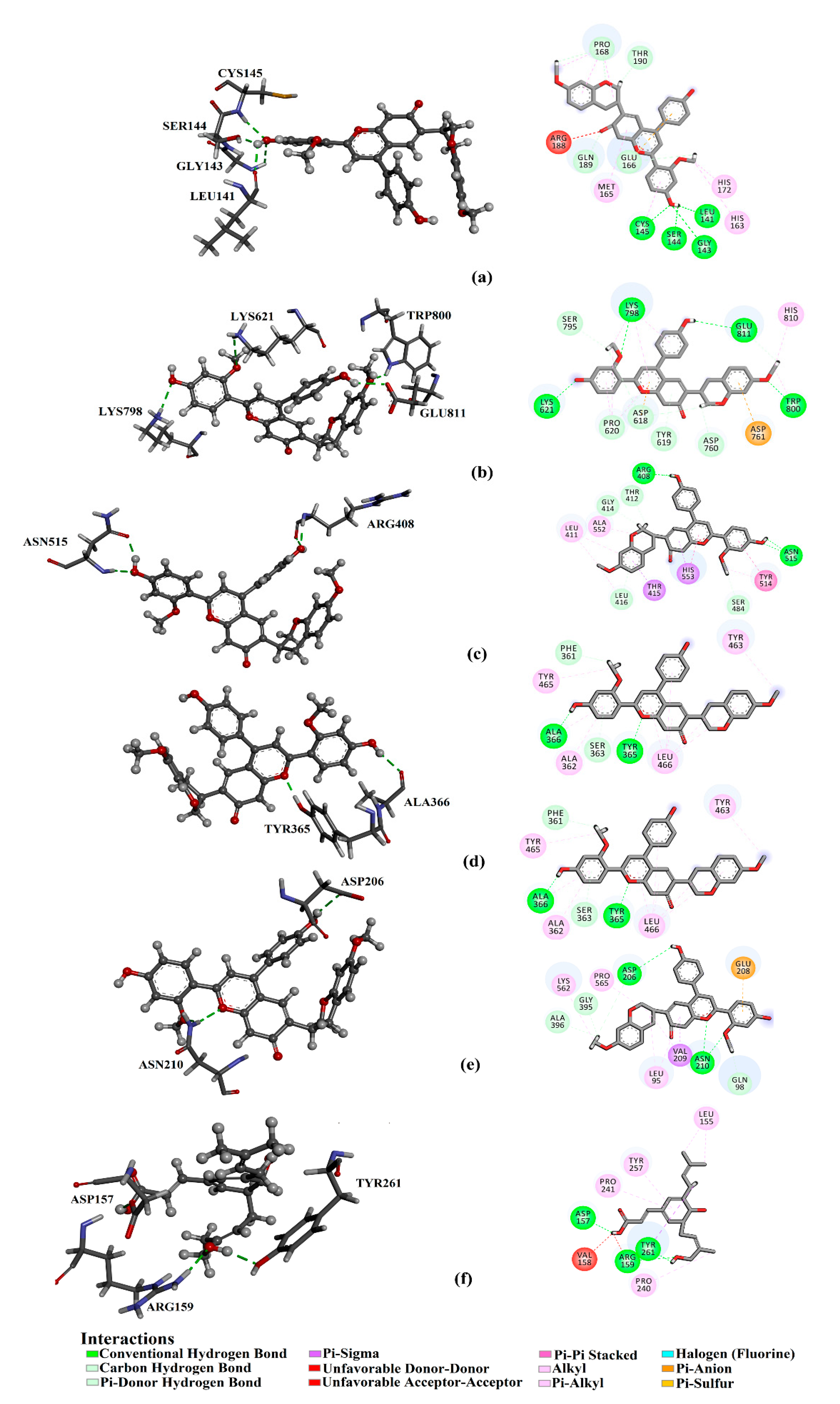{kind=link}
| Propolis Origin | Type of Extract | Antiviral Activities | References |
|---|---|---|---|
| South of Turkey (Hatay region) | 70% Ethanol | Herpes simplex virus (HSV-1) Assay: MTT colorimetric and Real-Time PCR Model: HEp-2 cell culture MIC = 130 μg/mL Tested doses: 25, 50, 75, 100, 200, 400, 800, 1600 and 3200 μg/mL PC: Acyclovir MIC = 65 μg/mL NC: Cell cultures without propolis or acyclovir Mode of action: Inhibits the replication after 24 h. (in vitro) | [43] |
| Herpes simplex virus (HSV-2) Assay: MTT colorimetric and Real-Time PCR Model: HEp-2 cell culture MIC = 250 μg/mL Tested doses: 25, 50, 75, 100, 200, 400, 800, 1600 and 3200 μg/mL PC: Acyclovir NC: Cell cultures without propolis or acyclovir Mode of action: Inhibits the replication after 48 h. (in vitro) | [43] | ||
| Northwestern Parana state, Brazil | 70% Ethanol | Herpes simplex virus 1 (HSV-1) Assay: Attachment and penetration; attachment, virucidal, and plaque formation Model: Vero cells EC50 = 3.20 ± 0.14 µg/mL EC50 for attachment and penetration: 1.21 ± 0.14 µg/mL EC50 for attachment: 0.40 ± 0.07 µg/mL EC50 for virucidal: 3.84 ± 0.15 µg/mL PC: Acyclovir EC50 = 1.33 ± 0.08 µg/mL NC: Untreated cells Mode of action: Inhibits viral infection and induces virion damage. (in vitro) | [44] |
| Santa Flora City (RS-Brazil) | 70% Ethanol | Herpes simplex virus 2 (HSV-2) Assay: Plaque reduction Model: Female BALB/c mice Dose of Pre-treatment: 50 mg/kg, once a day Dose of Post-treatment: 50 mg/kg for 5 days more PC: Not reported NC: Untreated cells Mode of action: Reduces extravaginal lesions and the histological damage caused by HSV-2 infection in vaginal tissues of animals. (in vivo and ex vivo) | [45] |
| Moravia, Czech Republic |
| Herpes simplex virus 1 (HSV-1) Assay: Plaque reduction Model: RC-37 cells Aqueous extract: TC50 (%): 0.04; IC50 (%): 0.0004; SI: 100 Ethanol extract: TC50 (%):0.0017; IC50 (%): 0.000035; SI: 485 PC: Heparin-Na and acyclovir NC: Untreated cells Mode of action: Mask viral compounds which are necessary for adsorption or entry into host cells. (in vitro) | [13] |
| Moravia, Czech Republic |
| Herpes simplex virus type 2 (HSV-2) Assay: Plaque reduction Model: RC-37 cells IC50% for aqueous extract: 0.0005; SI: 80 IC50% for ethanolic extract: 0.0004; SI: 42.5 PC: Acyclovir Inhibits replication of 98.8% NC: Untreated cells without drugs Mod of action:
| [46] |
| Canada | 70% Ethanol | Herpes simplex virus 1 and 2 Assay: Virucidal assay Model: MDBK cell PPE marked effect: 3.2 mg/mL PC: Acyclovir NC: Not reported Mode of action:
| [47] |
| Botucatu, Brazil | 70% Ethanol | Poliovirus type 1 (PV1) Assay: Real-time PCR Model: HEp-2 cells
PC: Cells with virus but without propolis NC: Not reported Mode of action: Causes RNA degradation before the virus entry into cells; also affects the steps of viral cycle replication into cells. (in vitro) | [48] |
| Rennes (France) | 80% Ethanol | Poliovirus type 2 (PV), vesicular stomatitis virus (VSV), adenovirus type 2 (Adeno-2), herpes simplex Virus (HSV-1, HSV1-R, and HSV-2) Assay: Plaque reduction Model: Vero cells Dose: 30 µg/mL PC: NR NC: Without propolis Mode of action: NR (In vitro) | [12] |
| Moravia, Czech Republic | 90% Ethanol | Varicella zoster virus (VZV) Assay: Plaque reduction and PCR Model: Cell line of human embryonic lung fibroblasts (LEP) IC50: 64 μg/mL Tested doses: 100 μg/mL PC: Acyclovir (conc: 50 μg/mL) NC: Untreated culture Mode of action: Masking viral compounds which are necessary for entry into host cells. (in vitro) | [49] |
| Paraná state and Alagoas state, Brazil | 80% Ethanol | Enterovirus surrogates MS2 and Av-08 bacteriophage Assay: Plaque formation Tested conc.: 100, 500, and 1000 μg/mL PC: Not reported NC: Bacteriophage with 1 mL of E. coli O157 bacteria without the addition of the propolis extracts Mode of action: Inhibits viral polymerase and the binding of viral nucleic acid or capsid proteins. (in vitro) | [50] |
| Tavarnelle Val di Pesa, Firenze, Italy | 80% Ethanol | Influenza A virus H1N1 Model: Madin-Darby canine kidney cells (MDCK) Anti-neuraminidase IC50 (µg/mL): 35.29 ± 4.08 PC: Oseltamivir IC50 (µg/mL): 5.88 ± 0.89 NC: Not reported Mode of action:
| [31] |
| Brazil and China | 95% Ethanol | Human immunodeficiency virus type 1 (HIV-1) Model: CD4+ lymphocytes and microglial cell cultures At 66.6 µg/mL give inhibition 85 and 98% PC: Zidovudine (AZT) or indinavir NC: Culture medium alone Mode of action: Inhibits viral entry. (in vitro) | [51] |
| Southern Brazil | Ethanol | Influenza A/PR/8/34 (H1N1) Assay: Plaque reduction assay Model: Mice Dose: 10 mg/kg Propolis (EC50 µg/mL):
EC50 = 20.2 ± 11.7 µg/mL NC: 1% ethanol solution Mode of action: Not reported. (in vitro and in vivo) | [52] |
| Cuautitlan Izcalli, State of Mexico | 70% Ethanol | Pseudo Rabies Virus (PRV) Model: Monolayers of Madin-Darby bovine kidney (MDBK) cells Assay: Plaque assay Tested dose: 0.5 mg /mL PC: Not reported NC: Not infected MDBK cell culture Plaque forming unit: 8.7 Mode of action:
| [53] |
| Origin | Compound Names | Antiviral Activities/Species | References |
|---|---|---|---|
| Southern Brazil | Melliferone | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 0.205 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] |
| Moronic acid | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 18.6 µg/mL EC50 < 0.1 µg/mL
EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] | |
| Anwuweizonic acid | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 2.14 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] | |
| Betulonic acid | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 1.8 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication (in vitro) | [55] | |
| 4-Hydroxy-3-methoxypropiophenone | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 18.8 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] | |
| 4-Hydroxy-3 methoxybenzaldehyde | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: >100 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] | |
| 3-(3,4-Dimethoxyphenyl)-2-propenal | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 18.9 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] | |
| 12-Acetoxytremetone | Human immunodeficiency virus (HIV) Model: H9 lymphocytes IC50: 2.07 µg/mL PC: AZT IC50: 500 µg/mL EC50: 0.00289 µg/mL NC: Infected cells with culture medium Mode of action: Inhibits viral replication. (in vitro) | [55] | |
| Uniflora Apicultores Associados (Olimpia, Brazil)/ Moravia, Czech Republic | Kaempferol | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2: IC50 = 7.3 ± 4.54 µM PC: Ribavirin IC50 =270.1 ± 35.94 µM HRV-3: IC50 = 11.9 ± 0.42 µM PC: Ribavirin IC50 =307.9 ± 5.53.94 µM HRV-4 IC50 = 12.9 ± 1.15 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] |
| p-Coumaric acid | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 371.2 ± 7.74 µM PC: Ribavirin IC50 = 270.1 ± 35.94 µM HRV-3 IC50 = 454.5 ± 3.16 µM PC: Ribavirin: IC50 = 307.9 ± 5.53.94 µM HRV-4 IC50 = 604.3 ± 50.93 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Galangin | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 20.0 ± 8.07 µM PC: Ribavirin IC50 =270.1 ±35.94 µM HRV-3 IC50 = 116.2 ± 0.85 µM PC: Ribavirin IC50 = 307.9 ± 5.53.94 µM HRV-4 IC50 = 88.1 ± 28.71 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Herpes simplex virus 1 (HSV-1) Assay: Plaque reduction Model: RC-37 cells IC50 (%): 0.00045; SI: 3.3 PC: Heparin-Na and acyclovir NC: Untreated cells Mode of action: Masks viral compounds which are necessary for adsorption or entry into host cells. (in vitro) | [13,15] | ||
| Quercetin | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 34.1 ± 10.33 µM PC: Ribavirin IC50 =270.1 ± 35.94 µM HRV-3 IC50 = 15.5 ± 2.29 µM PC: Ribavirin: IC50 =307.9 ± 5.53.94 µM HRV-4 IC50 = 18.2 ± 4.14 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Fisetin | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 40.9 ± 15.20 µM PC: Ribavirin IC50 =270.1 ± 35.94 µM HRV-3 IC50 = 67.1 ± 3.50 µM PC: Ribavirin IC50 =307.9 ± 5.53.94 µM HRV-4 IC50 = 66.4 ± 13.28 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Chrysin | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 17.3 ± 9.83 µM PC: Ribavirin: IC50 =270.1 ± 35.94 µM HRV-3 IC50 = 16.1 ± 4.80 µM PC: Ribavirin IC50 =307.9 ± 5.53.94 µM HRV-4 IC50 = 24.4 ± 5.27 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Herpes simplex virus 1 (HSV-1) Assay: Plaque reduction Model: RC-37 cells IC50 (%): 0.00003; SI: 20 PC: Heparin-Na and acyclovir NC: Untreated cells Mode of action: Masks viral compounds which are necessary for adsorption or entry into host cells. (in vitro) | [13,15] | ||
| Luteolin | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 37.4 ± 2.10 µM PC: Ribavirin IC50 =270.1 ± 35.94 µM HRV-3 IC50 = 20.4 ± 2.63 µM PC: Ribavirin IC50 =307.9 ± 5.53.94 µM HRV-4 IC50 = 14.7 ± 7.86 µM PC: Ribavirin: IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Acacetin | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2: IC50 = 163.2 ± 18.97 µM PC: Ribavirin: IC50 = 270.1 ±35.94 µM HRV-3: IC50 = 107.6 ± 18.30 µM PC: Ribavirin: IC50 = 307.9 ± 5.53.94 µM HRV-4: IC50 = 102.3 ± 3.59 µM PC: Ribavirin: IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Caffeic acid | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 67.2 ± 5.89 µM PC: Ribavirin IC50 =270.1 ± 35.94 µM HRV-3 IC50 = 52.2 ± 2.61 µM PC: Ribavirin IC50 =307.9 ± 5.53.94 µM HRV-4 IC50 = 66.1 ± 15.43 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Ferulic acid | Human rhinovirus (HRV)-2, HRV-3, and HRV-4 Assay: Ulforhodamine B and real-time reverse transcription PCR Model: HeLa cells HRV-2 IC50 = 175.1 ± 29.10 µM PC: Ribavirin IC50 = 270.1 ±35.94 µM HRV-3: IC50 = 248.7 ± 22.30 µM PC: Ribavirin IC50 = 307.9 ± 5.53.94 µM HRV-4 IC50 = 232.3 ± 5.05 µM PC: Ribavirin IC50 = 323.9 ± 31.16 µM NC: DMSO Mode of action: Blocks or reduces the entrance of the viruses into the cells to protect the cells from virus destruction and abate virus replication. (in vitro) | [13,15] | |
| Brazil | Apigenin | Anti-influenza virus Assay: Plaque reduction Model: MDCK A/PR/8/34(H1N1) EC50 = 15.3 ± 3.0 µg/mL A/Toyama/129/2011(H1N1) EC50 = 17.8 ± 8.7 µg/mL A/Toyama/26/2011(H1N1 EC50 = 8.1 ± 4.7 µg/mL PC: Ribavirin EC50 = 19.2 ± 7.5µg/mL NC: Distilled water Mode of action: Suppresses the stage of virus replication after adsorption and/or invasion. (in vitro) | [56] |
| Artepillin C | Anti-influenza virus Assay: Plaque reduction Model: MDCK A/PR/8/34(H1N1) EC50 ˃ 40 µg/mL A/Toyama/129/2011(H1N1) EC50 ˃ 40 µg/mL A/Toyama/26/2011(H1N1 EC50 ˃ 40 µg/mL PC: Ribavirin EC50 = 19.2 ± 7.5µg/mL NC: Distilled water Mode of action: Suppresses the stage of virus replication after adsorption and/or invasion. (in vitro) | [56] | |
| Kaempferol | Anti-influenza virus Assay: Plaque reduction Model: MDCK A/PR/8/34(H1N1) EC50 = 38.2 ± 17.1 µg/mL A/Toyama/129/2011(H1N1) EC50 = 21.7 ± 5.5 µg/mL A/Toyama/26/2011(H1N1 EC50 = 24.8 ± 4.3 µg/mL PC: Ribavirin EC50 = 19.2 ± 7.5µg/mL NC: Distilled water Mode of action: Suppresses the stage of virus replication after adsorption and/or invasion. (in vitro) | [56] | |
| Caffeic acid | Anti-influenza virus Assay: Plaque reduction Model: MDCK A/PR/8/34(H1N1) EC50 >100 µg/mL A/Toyama/129/2011(H1N1) EC50 = 49.7 ± 5.0 µg/mL A/Toyama/26/2011(H1N1 EC50 > 100 µg/mL PC: Ribavirin EC50 = 19.2 ± 7.5µg/mL NC: Distilled water Mode of action: Suppresses the stage of virus replication after adsorption and/or invasion. (in vitro) | [56] | |
| Coumaric acid | Anti-influenza virus Assay: Plaque reduction Model: MDCK A/PR/8/34(H1N1) EC50 = 31.5 ± 1.3µg/mL A/Toyama/129/2011(H1N1) EC50 = 16.4 ± 6.6 µg/mL A/Toyama/26/2011(H1N1 EC50 = 27.0 ± 4.9 µg/mL PC: Ribavirin EC50 = 19.2 ± 7.5µg/mL NC: Distilled water Mode of action: Suppresses the stage of virus replication after adsorption and/or invasion. (in vitro) | [56] | |
| Caffeic acid phenethyl ester (CAPE) | Herpes simplex viruses (HSV-1 and HSV-2) Assay: Microscopic Fourier transform infrared spectroscopy (FTIR) Model: Mouse embryo fibroblasts (MEF) Tested doses: 10 and 50 µM PC: NR NC: Untreated cells (in vitro) | [57] |
| Place of Propolis Collection | Type of Extract | Immunomodulatory Effect | References |
|---|---|---|---|
| Northeast of Algeria | 85% Ethanol | Celiac Disease (immune-mediated enteropathy) Assay: Griess method, ELIZA, and immunofluorescence assay Model: Peripheral blood mononuclear cells (PBMCs) Tested doses: 1, 50, and 100 µg/mL Mode of action:
| [99] |
| Brazil | 70% Ethanol | Antifungal immunity Assay: Flow-cytometry and ELISA Model: Human monocytes Tested doses: 5, 10, 25, 50, and 100 μg/mL Mode of action:
| [82] |
| Brazil | Ethanol | Antileishmaniasis Assay: Examination by SEM spectroscopy, Phagocytic Assay, ELISA Model: Human urine (in vitro), macrophages (in vivo) Tested doses: 5, 10, 25, 50, and 100 μg/mL (in vitro) 2.5, 5, or 10 mg/kg (in vivo) Mode of action:
| [88] |
| Brazil | Propolis standard extract (dry extract) | Allergic asthma Assay: Flow cytometry, real time-PCR, and ELIZA Model: Bronchoalveolar lavage fluid (BALF) of allergic mice Tested doses: 150 mg/Kg every day for 17 days Mode of action:
| [70] |
| Northern Morocco | Ethanol | Immunomodulation Assay: Cytotoxic and cytostatic assays, MTT assay, and ELISA Model: MCF-7, HCT, THP-1, and PBMNCs cell lines IC50: 479.22, 108.88, and 50.54 μg/mL Tested doses: 125 and 250 μg/mL Mode of action:
| [100] |
| Brazil | 0.1% Ethanol | Leishmaniasis Assay: Cytometric bead array assay, indirect immunofluorescence assay Model: PBMNCs Tested doses: 5 and 25 µg/mL Mode of action: Increases IL-4 and IL-17 and decreases IL-10. (in vitro) | [90] |
| Iran | 30% Ethanol | Immunomodulation on tumor-bearing mice with disseminated candidiasis Assay: ELISA Model: Mouse mammary tumor Tested doses: 100 mg/kg Mode of action: Decreases IL-4 and IL-10 levels and increases TNF-α and IFN-γ levels. (in vitro) | [101] |
| Iran | Ethanol | Lipopolysaccharide-induced inflammation Assay: MTT assay, the Griess method, flow cytometry real-time PCR, and MTT assays Model: Murine macrophage (RAW 264.7) IC50: 15 ± 3.2 µg/mL Tested doses: 15, 1.5, 0.15 µg/mL Mode of action: Inhibits NO and ROS production and then decreases COX-2, IL-1β, and IL-6 gene expression. (in vitro) | [102] |
| Brazil | 70% Ethanol | Immunomodulation Assay: MTT assay, ELISA, RT-qPCR, flow cytometry Model: Human DCs Tested doses: (5, 10, 20 and 40 μg/mL) Mode of action: Activates human DCs; induces the NF-kB signaling pathway and TNF-α, IL-6, and IL-10 production; inhibits the expression of hsamiR-148a and hsa-miR-148b; and increases of miR-155 expression. (in vitro) | [85] |
| Brazil | 70% Ethanol | Immunomodulation Assay: ELISA, real-time PCR Model: Peritoneal macrophages and spleen cells in BALB/c mice Tested doses: 200 mg/kg, 0.1 mL for 3 consecutive days by gavage Mode of action:
| [84] |
| Brazil Cuba Mexico | 70% Ethanol | Immunomodulation Assay: MTT assay and ELISA Model: PBMNCS Tested doses: 0.2, 1.0, 2.0, 10.0, and 20.0 μg/mL Mode of action: Stimulates both TNF-α and IL-10 production by monocytes. (in vitro) | [103] |
| NR | Aqueous | Immunomodulatory activity in Zymosan-induced paw oedema Assay: AP complement assay, microtiter assay Model: Zymosan-induced paw oedema in mice strain ICR Tested doses: 150 mg/kg Mode of action: Inhibits the formation of edema by activation of alternative pathway (AP) complement. (in vivo) | [104] |
| Turkey | 96% Ethanol | Immunomodulation Assay: MTT, high-pressure liquid chromatography, ELISA Model: PBMNCS Tested doses: NR Mode of action: Dose-dependent downregulation by induction of neopterin production and tryptophan degradation and inhibition of TNF-α and IFN-γ levels. (in vitro) | [105] |
| Brazil | 70% Ethanol | Immunomodulation Assay: Cytotoxicity assay, ELISA Model: Peritoneal macrophages of BALB/c mice Tested doses: 5, 50, and 100 mg/well Mode of action:
| [106] |
| Brazil | 70% Ethanol | Immunomodulation Assay: Radioimmunoassay, quantitative real time PCR, ELISA Model: C57BL/6 mice Tested doses: 200 mg/kg Mode of action: Increases the gene expression of TLR-2 and TLR-4. (in vivo) | [81] |
| Brazil | 70% Ethanol | Ant-inflammatory Assay: 2,2-Diphenyl-1-picrylhydrazyl free radical (DPPH) scavenging method Griess reaction, MTT assay, ELISA Model: Raw 264.7 cells and a mouse leukemic monocyte macrophage cell line (ATCC TIB-71) Tested doses: 5, 10, 25, 50, and 100 μg/mL Mode of action: Inhibits NO production in macrophages and suppresses p38 MAPK, JNK1/2 pathways. (in vitro) | [107] |
| Brazil | Lyophilized samples | Immunomodulation Assay: Cotton Pellet Granuloma, ELISA, histopathology analysis Model: Swiss and Balb/c mice Tested doses: 5 mg/kg Mode of action: Decreases the concentration of TNF-α and IL-6 and increases TGF-β and IL-10. (in vivo) | [108] |
| Brazil | 70% Ethanol | Antifungal immunity Assay: MTT, Flow Cytometry, ELISA Model: PBMNCS Tested doses: 5, 10, 25, 50, and 100 μg/mL Mode of action:
| [83] |
| Brazil | 70% Ethanol | Immunomodulatory in acutely stressed mice Assay: ELISA Model: BALB/c male mice Tested doses: 200 mg/kg/day, 0.1 mL Mode of action: Increases IL-4 production in stressed mice. (in vivo) | [109] |
| Brazil | 70% Ethanol | Immunomodulatory activity melanoma-bearing mice submitted to stress Assay: ELISA, Real-time PCR Model: C57BL/6 male mice Tested doses: 200 mg/kg Mode of action: Upregulates the expression of TLR-2, IL-10, and IFN-γ. (in vivo) | [110] |
| Brazil | 70% Ethanol | Immunomodulation Assay: Real Time PCR, ELISA Model: BALB/c male mice Tested doses: 200 mg/kg/day, 0.1 mL Mode of action: Immunorestorative role in TLR-4 expression. (in vivo) | [111] |
| Brazilian propolis provided by Yamada Apiculture Center, Japan | Ethanol | Rheumatoid arthritis Assay: RT-PCR, ELISA, flow cytometry Model: DBA/1J mice Tested doses: (6.7 mg/g diet) and high dose (20 mg/g diet) propolis Mode of action: Inhibits production of IL-17 and the differentiation of Th17 cells. (in vivo) | [75] |
| Brazil | 70% Ethanol | Fungicidal activity Assay: Flow cytometry, ELISA Model: PBMNCS Tested doses: 5, 10, 25, 50, and 100 μg/mL Mode of action: Downregulates the expression of TLR-2 and HLA-DR, inhibits TNF-α and IL-10 production, and increases fungicidal activity. (in vitro) | [86] |
| Brazilian propolis provided by Yamada Apiculture Center, Japan | Ethanol | Rheumatoid arthritis Assay: Cytotoxicity assays, Flowcytometry, Western blot analysis Model: splenocytes from Balb/c mice Tested doses: 12, 48 μg/mL Mode of action:
| [78] |
| Beekeeping Section, UNESP | 70% Ethanol | Immunomodulation Assay: Real-time PCR, ELISA Model: Male BALB/c mice Tested doses: 200 mg/kg, 0.1 mL Mode of action: Inhibits the production of IFN-γ. (in vivo) | [112] |
| Bulgaria | Ethanol | Prophylactic activity against Gram-negative bacteria Assay: Negative Limulus amoebocyte lysate assay Model: Strain ICR mice Tested doses: 150 mg g-1 Mode of action: Production of Clq Macrophages, and change in the alternative complement pathway hemolysis. (in vivo) | [113] |
| Croatia |
| Immunomodulatory effect against irinotecan-induced toxicity and genotoxicity Assay: Hematological analysis, peripheral blood micronucleus (MN) assay Model: Male albino mice of the Swiss strains Tested doses: 100 mg/kg Mode of action: Inhibits the growth of Ehrlich ascites tumors (EAT) by activation of macrophages and neutrophils, which inhibits Irinotecan induced toxicity. (in vivo) | [92] |
| Croatia | WSDP | Antimetastatic effect against lung cancer Assay: Flow cytometry, colorimetric Griess reaction Model: CBA inbred mice Tested doses: 50 or 150 mg/kg Mode of action: Suppression of metastasis by activation of macrophages and production of TNF-α, H2O2, and nitric oxide NO. (in vivo) | [114] |
| Brazil | 70% Ethanol | Immunomodulation Assay: ELISA, spleen cells proliferation assay Model: Spleen cells of BALB/c male Tested doses: 2.5, 5, and 10 mg/kg, for 3 days Mode of action: Decreases splenocyte proliferation and stimulates IFN-γ production. (in vitro) | [115] |
| Brazil Croatia | WSDP | Antimetastatic effect against lung cancer Assay: Flow cytometry, Plaque-forming cells assay, hematological analysis Model: Male and female CBA inbred mice Tested doses: 50 or 150 mg/kg and 50 mg/mL Mode of action: Suppression of metastasis and increased level of LAF produced macrophages. (in vitro, in vivo) | [93] |
| NR | WSDP | Transplantable mammary carcinoma Assay: Colorimetric Griess reaction, hematological analysis Model: Male and female CBA inbred mice Tested doses: 50 or 150 µg/kg Mode of action: Inhibits tumor by macrophages activation, induces NO production, increases, the response of splenocytes to monoclonal antibodies, and inhibits [3 H]TdR incorporation into HeLa cells. (in vitro) | [116] |
| Brazil | 70% Ethanol | Antileishmanial activity Assay: MTT assay, real-time PCR Model: PBMNCs Tested doses: 5, 10, 25 µg/mL Mode of action: Reduces the expression of CCL5 and IFN-γ. (in vitro) | [89] |
| Brazil | Hydroalcoholic extract | Antileishmanial activity Assay: Neutral red method Model: Male Balb/C albino mice (lineage of Mus musculus), Tested doses: 1.5 mg/kg/day Mode of action: NR (in vitro) | [87] |
| Brazil | 70% Ethanol | Immunomodulatory activity in chronically stressed mice Assay: Griess Reaction, histopathological analysis Model: Male BALB/c mice Tested doses: 200 mg/kg Mode of action: Increases production of H2O2 and inhibits NO. (in vivo) | [117] |
| Bulgaria | WSDP | Immunomodulatory activity Assay: Popliteal lymph node assay Model: Macrophage Tested doses: 50 and 150 mg/kg Mode of action: Activation of macrophages, which induces the production of IL-1. (in vitro) | [118] |
| Croatia Brazil | WSDP | Immunomodulatory activity against Ehrlich ascites tumor Assay: Differential cell counts Model: Male albino mice of the Swiss strain Tested doses: 50 mg/kg Mode of action: Increases macrophage spreading activity. (in vivo) | [94] |
| Croatia | WSDP | Ehrlich ascites tumor Assay: Macrophage spreading assay, colorimetric Griess reaction, plaque-forming cells (PFC) assay, biuret method Model: Male and female CBA and Swiss albino inbred mice Tested doses: 50 mg/kg Mode of action: Increases cytotoxic T-cell, NK, and B cells activity. (in vivo) | [95] |
| Croatia Brazil | WSDP | Immunomodulatory activity in lung metastases Assay: Flow cytometry Model: Male and female CBA inbred mice Tested doses: 50 or 150 mg/kg Mode of action: Reduces metastases, delays tumor formation, and increases survival of treated animals. (in vivo) | [119] |
| Croatia Brazil | WSDP | Immunomodulatory activity in metastasis mammary carcinoma Assay: Griess method, plaque-forming cells assay, flow cytometry Model: Human cervical carcinoma cells (HeLa) Tested doses: 50 and 150 mg/kg Mode of action: Inhibits metastasis of mammary carcinoma, induces apoptosis, activates macrophages, increases CD4+ and CD8+ T cells, and induces the production of NO. (in vivo) | [96] |
| Bulgaria | 70% Ethanol | (Parasitemia) Trypanosoma cruzi Assay: Flow cytometry Model: Swiss mice Tested doses: 25 to 100 mg/kg Mode of action: Preferential expansion of CD8+, inhibits CD4+ CD69+ and CD8+ CD69+ in CD4+ CD44+ and CD8+ CD44+, and decreases CD8+CD62L in Trypanosoma cruzi-infected mice. (in vivo) | [120] |
| Activity/Disease | Therapeutic Effect | Preclinical/Clinical Trials/ Number of Participants | Dose/Administration Route | References |
|---|---|---|---|---|
| Anti-viral/genital herpes (HSV-2) | Heal genital herpetic lesions and reduce local symptoms | Randomized controlled trials/90 p | Four times daily for 10 days/topical | [122] |
| Anti-viral/HSV-1 | Treat herpetic skin lesions | NR | Propolis 3%/topical | [121] |
| Anti-viral/Herpes zoster | Heal skin lesion and reduce pain | Clinical trial/60 p | Propolis lotion (3 times/day topical) + Acyclovir (400 or 800 mg oral) for 28 days | [123,124] |
| Anti-viral/Herpes labialis | Reduce pain, short healing time with painless incrustation | Randomized, double-blind trial/150 p | Propolis (0.1%, 0.5% and 1%)/topical | [125] |
| Anti-viral/Herpes labialis | Reduce pain, burning, itching, tension, and swelling | Randomized, single-blind trial/379 p | Propolis 0.5%/5 times per 5 days/topical | [126] |
| Anti-viral/Herpes labialis | Treat episodes of herpes labialis | Randomized, controlled double-blind study/397 p | Propolis 0.5%/(0.2 g) 5 times per 5 days/topical | [127] |
| Anti-viral/dengue hemorrhagic fever virus | Reduce TNF-α levels and improve platelet counts | Randomized, double-blind, placebo-controlled trial/63 p | Propolis 200 mg three times a day for 7 days/oral | [3] |
| No. | Compound Name | 3CLpro | PLpro | RdRp | NSP13 | RBD | ACE2 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Docking Score (kcal/mol) | Binding Features (Hydrogen Bond Length in Å) | Docking Score (kcal/mol) | Binding Features (Hydrogen Bond Length in Å) | Docking Score (kcal/mol) | Binding Features (Hydrogen Bond Length in Å) | Docking Score (kcal/mol) | Binding Features (Hydrogen Bond Length in Å) | Docking Score (kcal/mol) | Binding Features (Hydrogen Bond Length in Å) | Docking Score (kcal/mol) | Binding Features (Hydrogen Bond Length in Å) | ||
| Darunavir | −8.2 | GLU166 (1.94, 2.88 Å), LEU167 (1.96 Å) | −3.8 | GLU196 (2.18, 2.30 Å), MET199 (2.11 Å) | −4.4 | TYR619 (2.11 Å), ASP760 (1.83 Å), GLU811 (2.24, 2.26 Å) | −7.1 | LEU411 (2.93 Å), THR412 (2.18 Å), GLY414 (2.89 Å), LEU416 (2.12 Å), ASN556 (2.11 Å), ARG559 (1.97 Å) | −3.3 | GLU498 (1.89 Å), GLN507 (2.17 Å), SER508 (1.92 Å) | −8.5 | TYR202 (1.97 Å), GLU208 (3.02 Å), LYS562 (2.81 Å) | |
| Favipiravir | −4.3 | GLU166 (1.96, 2.26 Å), ARG188 (1.89 Å), THR190 (2.15 Å) | −4.0 | GLU196 (1.99 Å) | −4.8 | TRP617 (1.88 Å), ASP761 (1.96 Å), ALA762 (1.80 Å), TRP800 (2.20 Å) | −4.3 | PRO513 (2.18 Å), ASN515 (1.89 Å), THR531 (1.76 Å), ASP533 (2.19 Å) | −4.2 | LYS472 (2.05 Å), SER508 (2.07 Å), GLU498 (2.20 Å) | −4.4 | ASP206 (2.07 Å), GLU208 (1.98 Å), ALA398 (1.80 Å) | |
| 1 | Retusapurpurin A | −9.4 | LEU141 (2.22 Å) GLY143 (2.93 Å), SER144 (1.90 Å), CYS145 (2.25 Å) | −8.0 | ARG157 (2.17 Å), GLU196 (2.70 Å), MET201 (2.20 Å) | −7.5 | LYS621 (2.42 Å), LYS798 (2.89 Å), TRP800 (2.29 Å), GLU811 (2.23 Å) | −9.4 | ARG408 (1.88, 2.11 Å) ASN515 (1.99, 2.15 Å), | −7.2 | TYR365 (1.71 Å), ALA366 (2.14 Å) | −10.4 | ASP206 (2.23 Å), ASN210 (2.16, 2.24 Å) |
| 2 | Capillartemisin A | −8.9 | HIS163 (1.99 Å), HIS164 (2.02 Å), THR190 (1.86 Å), GLN192 (2.32 Å) | −7.4 | ASP157 (1.81 Å), ASN260 (1.94 Å) | −6.7 | TYR619 (1.99 Å), ASP760 (2.08 Å), GLU811 (1.93 Å) | −7.0 | ARG177 (2.37 Å), ASN178 (2.91Å), ASN515 (1.83, 1.98, 2.02 Å), THR531 (2.10 Å), HIS553 (2.40 Å), | −4.8 | TYR463 (1.91 Å), GLU498 (2.33 Å), GLN507 (2.01, 2.22 Å) | −8.9 | GLU208 (2.35 Å), ALA396 (1.84 Å), GLU564 (1.75 Å) |
| 3 | Artepillin C | −8.8 | TYR54 (2.12, 2.34 Å), CYS44 (2.46 Å), GLU166 (2.25 Å), ASP187 (2.02 Å) | −8.0 | ASP157 (1.87 Å), ASN260 (1.87 Å) | −7.0 | ASP760 (2.09 Å), SER814 (1.89 Å) | −6.6 | ARG408 (2.02 Å), LEU411 (2.84 Å), LEU416 (2.11 Å) | −4.4 | TYR463 (2.08 Å) | −9.0 | GLU208 (2.35 Å), GLU564 (1.78 Å) |
| 4 | (E)-3-[4−Hydroxy-3-(2-hydroxy-3-methylbut-3-en-l-yl)-5-(3-methybut-2-en-l-yl)phenyl] propenoic acid | −8.7 | MET49 (2.01 Å), TYR54 (2.24 Å), GLU166 (1.81, 2.13 Å) | −7.9 | ASP157 (1.89 Å), TYR257 (2.02 Å), ASN260 (1.91 Å) | −6.3 | SER759 (2.99 Å), ASP760 (1.74, 1.88 Å), TRP800 (1.86 Å), GLU811 (2.08 Å) | −6.4 | ARG408 (1.95Å), LEU411 (2.12 Å), LEU416 (1.72 Å) | −2.7 | SER508 (1.92 Å) | −8.8 | GLN98 (1.86 Å), GLU208 (2.19 Å), GLU564 (2.06 Å) |
| 5 | Baccharin | −8.7 | TYR54 (2.06, 2.33 Å), CYS44 (2.19 Å), LEU141 (2.32 Å), GLY143 (2.82 Å), ASP187 (2.45 Å), GLN189 (1.90 Å) | −8.2 | ASP157 (1.87 Å), ARG159 (2.01 Å), TYR261 (2.00 Å) | −6.8 | ASP618 (2.19 Å), ASP760 (1.99 Å), ASP761 (2.15 Å), SER814 (2.07 Å) | −6.5 | ARG177 (2.48 Å), ASN178 (2.80 Å), SER485 (2.01 Å), ASN515 (1.93 Å), THR531 (1.92 Å) | −5.5 | TYR463 (1.89 Å), GLU498 (1.88 Å), SER508 (1.87 Å), LEU506 (2.13 Å), GLN507 (2.17, 2.60 Å) | −8.5 | GLN98 (2.23 Å), GLU208 (3.07 Å), SER563 (2.05 Å), TRP566 (2.19 Å) |
| 6 | (E)-2,3-Dihydroconiferyl p-coumarate | −8.6 | LEU141 (1.96 Å), GLY143 (3.06 Å), SER144 (2.14 Å), CYS145 (2.96 Å), THR190 (2.05 Å) | −7.6 | LYS150 (1.97 Å), LEU155 (2.01 Å), ASP259 (1.94 Å) | −5.7 | ASP760 (2.14 Å), CYS813 (2.63 Å), SER814 (1.96 Å) | −6.8 | ASN515 (2.52 Å), ASP533 (2.07Å) | −3.0 | GLU498 (1.83 Å), GLN507 (2.38 Å) | −8.0 | LEU95 (1.91 Å), GLN98 (1.97 Å), ASN210 (1.94 Å), TRP566 (2.23 Å) |
| 7 | Quercetin | −8.5 | HIS164 (2.23 Å), ASP187 (1.91 Å), THR190 (2.08, 2.08 Å), GLN192 (1.51 Å) | −6.8 | LYS150 (2.36 Å), GLU160 (2.28 Å), ASN260 (1.79 Å), TYR266 (2.98 Å), THR294 (2.09 Å), ALA239 (3.01 Å) | −6.6 | TYR619 (1.99, 2.03 Å), ASP761 (2.10 Å), GLU811 (2.33 Å) | −6.9 | ASN178 (2.25 Å), LEU416 (1.94 Å) | −4.7 | PHE361 (2.00, 2.08 Å), TYR365 (2.89 Å), TYR463 (2.16 Å) | −9.1 | GLU208 (1.88 Å), ASN210 (2.06, 2.16, 2.85 Å), SER563 (2.00 Å), GLU564 (2.92 Å) |
| 8 | Sakuranetin | −8.5 | HIS164 (2.22 Å), THR190 (2.01 Å) | −6.9 | ASP295 (1.65 Å), | −6.7 | TYR619 (1.86 Å), ASP761 (2.43 Å) | −6.9 | PRO405 (1.84 Å), ASN515 (1.96 Å) | −4.9 | SER363 (1.93 Å), TYR365 (1.77 Å), ASN364 (1.99, 2.73 Å) | −8.8 | ASP206 (1.99 Å), ASN210 (1.87, 2.00 Å) |
| 9 | Kaempferol | −8.4 | HIS164 (2.04 Å), ASP187 (1.99 Å), THR190 (1.81 Å) | −7.2 | LEU255 (1.91 Å), ASN260 (1.96 Å), THR294 (1.83 Å) | −6.4 | TRP617 (2.45 Å), TYR619 (1.84 Å), ASP761 (2.27 Å), TRP800 (2.12 Å) | −6.9 | ASN178 (2.03 Å), LEU416 (1.89 Å), ASN556 (1.87 Å), ARG559 (2.70 Å) | −4.9 | SER363 (2.08 Å), TYR365 (1.69 Å), ASN464 (1.92, 2.67 Å) | −8.9 | ASP206 (1.86 Å), GLU208 (1.89 Å), ASN210 (2.02, 2.15, 2.83 Å) SER563 (2.02 Å), GLU564 (2.99 Å) |
| 10 | Isorhamnetin | −8.3 | HIS164 (2.36 Å), ASP187 (1.98 Å), THR190 (2.07 Å) | −6.8 | GU160 (1.99 Å), ASN260 (1.89 Å), TYR266 (2.16 Å), THR294 (1.84 Å) | −6.5 | LYS621 (2.32 Å), CYS622 (2.19 Å), ASP760 (1.77 Å), ASP761 (1.90 Å), GLU811 (2.27 Å) | −6.6 | ASN178 (2.15 Å), ARG408 (2.18 Å), PRO513 (1.83 Å), THR531 (1.76 Å), ASN515 (2.89 Å) | −4.9 | SER363 (2.05 Å), TYR365 (1.75 Å), ASN464 (2.02, 2.63 Å) | −9.1 | ASP206 (2.23 Å), GLU208 (1.83 Å), ASN210 (2.01, 2.09, 3.03 Å), LYS562 (2.25 Å), SER563 (1.95 Å), GLU564 (2.98 Å) |
| 11 | Sakuranetin- 4′-methylether | −8.2 | HIS164 (2.25 Å), GLN192 (2.21 Å) | −7.0 | ARG159 (1.93 Å), THR294 (1.87 Å) | −6.5 | ASP761 (2.40 Å) | −7.0 | ARG408 (2.13 Å), ASN515 (2.12, 2.30 Å) | −4.9 | TYR365 (2.21 Å), ASN464 (2.22, 2.26 Å) | −8.8 | ASN210 (1.96, 1.98 Å) |
| 12 | Pinobanksin- 3-acetate | −8.1 | HIS164 (2.21 Å), GLU166 (1.96 Å), ASP187 (1.87 Å) | −6.8 | TYR266 (2.92 Å), ASP295 (2.08 Å) | −6.0 | LYS621 (2.45, 2.77 Å), CYS622 (1.89 Å), LYS798 (2.21 Å) | −6.7 | ASN178 (2.22 Å), THR409 (2.00 Å), ARG559 (2.29 Å) | −5.4 | PHE361 (1.71 Å), TYR365 (2.09 Å), ASN464 (2.14 Å) | −8.8 | ASN210 (2.05, 2.16, 2.92 Å) SER563 (2.21 Å) |
| 13 | Calycosin | −8.1 | ASP187 (2.0 Å), THR190 (2.15 Å) | −7.4 | LYS150 (2.42 Å), GLU160 (2.29 Å), THR294 (1.92 Å) | −6.2 | LYS619 (2.59 Å), LYS621 (2.22 Å), ASP623 (2.08 Å), LYS798 (2.33 Å) | -7.0 | ASN176 (1.97, 2.90 Å), TYR197 (1.78 Å), ASP533 (1.84 Å) | -5.1 | PHE361 (2.10 Å), TYR365 (1.65 Å) | −8.7 | ASP206 (2.10 Å), ASN210 (1.73 Å), LYS562 (2.10 Å), SER563 (2.08 Å), GLU564 (3.04 Å) |
| 14 | Myricetin | −8.0 | HIS164 (2.23 Å), ASP187 (1.91 Å), THR190 (2.02, 2.09 Å), GLN192 (2.52 Å) | −7.0 | ARG159 (3.01, 3.02 Å), ALA239 (2.84 Å), TYR257 (2.79 Å), ASN260 (1.83 Å), TYR261 (2.63 Å), THR294 (2.04 Å) | −6.8 | TYR619 (2.22 Å), LYS621 (2.83 Å), ASP760 (1.92 Å), ASP761 (1.95 Å), GLU811 (2.36 Å) | −7.0 | ASN178 (2.09 Å), PRO405 (1.80 Å), LEU416 (1.85 Å), ASN556 (1.91 Å), ARG559 (2.77Å) | −4.8 | ARG360 (2.95 Å), PHE361 (1.98, 2.32 Å), TYR363 (2.17 Å), ASN464 (1.84 Å) | −8.6 | ASP206 (1.88 Å), ASN210 (1.99,2.16 ALA396 (1.82, 1.84 Å), SER563 (2.09 Å), GLU564 (3.02 Å) |
| 15 | Apigenin | −8.0 | HIS164 (2.39 Å), ASP187 (1.96 Å), THR190 (1.89 Å) | −6.5 | LYS150 (1.96 Å), TYR261 (2.71 Å), THR294 (1.97 Å) | −6.8 | TRP617 (2.48 Å), TYR619 (1.86 Å), ASP761 (2.26 Å), TRP800 (2.10 Å) | −6.8 | LEU416 (1.95 Å), ASN556 (1.86, 2.19 Å) | −5.3 | SER363 (2.06 Å), TYR365 (1.70 Å), ASN464 (1.96, 2.65 Å) | −8.7 | ASP206 (1.92 Å), ASN210 (1.83, 1.86 Å), SER563 (2.00 Å) |
| 16 | Daidzein | −8.0 | HIS164 (3.09 Å), THR190 (2.04 Å) | −6.8 | LEU155 (2.69 Å), THR294 (1.87 Å) | −5.9 | ASP623 (1.68 Å), TYR619 (2.13 Å), ASP761 (1.71 Å) | −6.8 | ALA406 (2.13Å), ASN515 (1.96Å) | −5.9 | PHE361 (1.69 Å), TYR365 (2.10 Å), ASN364 (1.81 Å), LEU482 (2.07 Å) | −7.9 | GLN198 (2.58 Å), GLU564 (1.91 Å) |
| 17 | Chrysin | −7.8 | GLU166 (2.09 Å), THR190 (1.91 Å) | −6.9 | ARG159 (1.79, 2.19, 2.94 Å), THR294 (1.84 Å) | −6.2 | LYS621 (1.99 Å), ASP623 (1.91, 2.17 Å) | −7.1 | ASN178 (1.96, 2.12 Å), ASP533 (2.07 Å) | −5.6 | SER363 (2.04 Å), TYR365 (1.74 Å), ASN464 (2.03, 2.71 Å) | −8.5 | ASN210 (2.03, 2.07 Å), SER563 (2.02 Å), GLU564 (3.03 Å) |
| 18 | Liquiritigenin | −7.8 | ASP187 (1.95 Å), THR190 (1.96 Å) | −7.0 | GLY156 (1.99 Å), ASP295 (1.84 Å) | −6.2 | LYS621 (2.10 Å), CYS622 (2.95 Å), ASP623 (2.07 Å), ASP760 (2.35 Å) | −7.0 | ASN178 (2.08 Å), LEU416 (2.31, 2.54 Å), ASP533 (2.26 Å) | −5.9 | GLU420 (1.77 Å), TYR463 (1.98 Å), GLY510 (1.85 Å) | −8.4 | ASP188 (1.92 Å), GLN192 (1.79 Å), SER545 (2.04 Å) |
| 19 | Luteolin | −7.8 | TYR54 (2.76 Å), HIS164 (2.17 Å), THR190 (2.06 Å), GLN192 (2.42 Å) | −7.1 | LYS150 (2.43 Å), GLU160 (2.29 Å), TYR266 (2.17 Å), THR294 (1.90, 2.04 Å) | −6.9 | TYR619 (2.00, 2.01 Å), ASP761 (2.06 Å), GLU811 (2.33 Å) | −6.9 | ASN78 (2.30 Å), LEU416 (2.09 Å), ASN556 (2.07Å) | −4.9 | PHE361 (1.77, 1.96 Å), TYR365 (2.45 Å), TYR463 (2.16 Å) | −8.9 | ASP206 (1.88, 1.98 Å) ASN210 (1.82, 1.86 Å) SER563 (2.02 Å) |
| 20 | Vestitone | −7.8 | HIS163 (2.04 Å), GLU166 (2.11 Å), ASP187 (2.54 Å) | −7.5 | ARG159 (1.94 Å), TYR261 (2.15, 2.44 Å) | −5.9 | ASP623 (1.99 Å), ASP760 (2.13 Å) | −7.5 | ASP533 (1.68 Å) | −4.7 | TYR365 (1.87 Å), TYR463 (1.95 Å) | −7.6 | LYS94 (1.74 Å), ALA396 (2.11 Å), TRP566 (2.28 Å) |
| 21 | Caffeic acid phenethyl ester (CAPE) | −7.8 | LEU141 (2.11, 2.12 Å), GLY143 (2.44 Å), SER144 (2.15 Å), HIS163 (2.09 Å), GLU166 (2.35 Å) | −7.5 | TYR266 (1.97 Å) ASP295 (1.78, 2.13 Å) | −6.5 | TYR619 (2.13, 2.21 Å), ASP760 (1.96 Å), CYS813 (2.72 Å), SER814 (2.03 Å) | −6.5 | THR415 (1.83, 2.09 Å) | −4.0 | GLU498 (1.97, 2.04 Å) | −8.2 | ASN210 (1.95, 2.47 Å) ALA396 (1.95, 1.97 Å), TRP564 (2.29 Å) |
| 22 | Formononetin | −7.7 | PHE140 (2.66 Å), CYS145 (2.38 Å), GLU166 (1.97 Å) | −7.2 | THR294 (1.77 Å) | −6.2 | LYS621 (1.81 Å), LYS798 (1.94 Å) | −6.8 | ASN515 (1.94 Å) | −5.3 | PHE361 (1.69 Å), TYR365 (2.11 Å), ASN464 (1.81 Å) | −7.6 | TYR196 (2.22 Å), GLU208 (1.70 Å), ASN210 (2.11 Å) |
| 23 | Pinocembrin | −7.7 | GLU166 (2.12 Å), THR190 (1.76 Å) | −7.0 | ARG159 (1.82, 2.10, 2.86 Å) | −6.0 | TRP617 (3.08 Å), ASP761 (1.99 Å), ALA762 (1.81 Å), SER814 (2.35 Å) | −7.3 | ASN178 (1.97, 2.15 Å), ASP533 (2.02 Å) | −5.1 | SER363 (2.02 Å), TYR365 (1.79 Å), ASN464 (2.11, 2.71 Å) | −8.4 | ASN210 (2.02, 2.14 Å) SER563 (2.11 Å), GLU564 (3.03 Å) |
| 24 | Isoliquiritigenin | −7.6 | HIS164 (2.36 Å), ASP187 (1.98 Å), THR190 (2.07 Å) | −6.8 | ALA239 (2.33 Å), ASP295 (1.77 Å) | −6.0 | TYR617 (1.21 Å), ASP761 (2.18 Å), GLU811 (1.96 Å), SER814 (1.93 Å) | −7.1 | ALA406 (2.09 Å), ARG408 (2.37, 3.08 Å), LEU416 (2.14, 2.23 Å), ASP533 (2.09 Å) | −4.3 | PHE361 (1.95 Å), TYR365 (1.90 Å) | −8.1 | ASN210 (1.96, 2.07 Å), SER563 (2.15 Å) |
| 25 | Violanone | −7.6 | CYS44 (1.91 Å) | −7.3 | LEU155 (2.04 Å), THR294 (2.02 Å) | −6.1 | ASP164 (1.89 Å), LYS621 (2.31, 2.13 Å), TYR619 (2.30 Å), LYS798 (2.07 Å) | −7.3 | ARG408 (2.12 Å), ASN515 (2.09 Å) | −4.9 | SER363 (1.88, 1.96, 2.32 Å), ALA366 (2.13 Å), TYR463 (1.96 Å) | −8.1 | LYS94 (2.08 Å), GLN98 (2.50 Å) ASN210 (2.20, 2.49 Å) ALA396 (1.90 Å), TRP566 (2.24 Å) |
| 26 | Drupanin | −7.5 | HIS164 (1.96 Å), THR190 (1.83 Å), GLN192 (2.06 Å) | −7.0 | ASP157 (1.87 Å), ASN260 (1.94 Å) | −6.3 | ASP760 (1.85 Å) | −6.1 | ASN178 (1.90 Å), TYR179 (1.72 Å), SER485 (2.08 Å), ASN515 (1.91 Å) | −4.0 | GLU498 (1.68 Å), SER508 (2.01 Å) | −7.8 | ASN210 (2.10 Å), GLU564 (1.70 Å) |
| 27 | Galangin | −7.5 | HIS164 (2.21 Å), ASP187 (1.92 Å) | −6.8 | ARG159 (1.85 Å), THR294 (1.73 Å) | −5.8 | TRP617 (2.40 Å), ASP761 (2.33 Å), TRP800 (2.15 Å), SER814 (3.04 Å) | −6.6 | LEU416 (1.91 Å), ASN556 (1.85 Å), ARG559 (2.71 Å) | −5.3 | SER363 (2.06 Å), TYR365 (1.74 Å), ASN464 (2.01 Å) | −8.7 | GLU208 (1.88 Å), ASN210 (2.04, 2.16, 2.86 Å), SER563 (1.99 Å), GLU564 (2.98 Å) |
| 28 | Pinobaskin | −7.5 | HIS164 (2.23 Å), ASP187 (1.90 Å) | −6.7 | ARG159 (1.83 Å), THR294 (1.73 Å) | −5.6 | TRP617 (2.39 Å), ASP761 (2.30 Å), TRP800 (2.17 Å), SER814 (3.04 Å) | −6.3 | LEU416 (1.91 Å), ASN556 (1.86 Å), ARG559 (2.70 Å) | −5.0 | SER363 (2.04 Å), TYR365 (1.76 Å), ASN464 (2.01 Å) | −8.6 | GLU208 (1.86 Å), ASN210 (2.04, 2.18, 2.85 Å), SER563 (1.99 Å), GLU564 (2.98 Å) |
| 29 | Pinostrobin | −7.4 | SER144 (2.92 Å), HIS163 (1.98 Å) | −6.9 | ARG159 (1.80 Å) | −6.1 | ASP761 (2.23 Å) | −6.7 | LEU416 (2.05, 2.30 Å), ARG559 (1.93 Å) | −5.1 | TYR365 (2.20 Å), ASN464 (2.23, 2.27 Å) | −8.6 | ASN210 (1.80, 1.98 Å) |
| 30 | Biochanin A | −7.4 | THR190 (1.78 Å), GLN192 (3.06 Å) | −7.5 | ARG159 (1.93 Å), TYR261 (2.13 Å) ALA239 (2.92 Å) | −6.3 | TYR619 (3.05 Å), LYS621 (2.06 Å), ASP760 (1.82 Å), SER795 (2.79 Å) | −7.0 | ASN178 (2.14 Å), ASN515 (1.96 Å) | −5.1 | PHE361 (1.75 Å), TYR365 (2.11 Å), ASN464 (1.78 Å) | −7.9 | TYR196 (2.22 Å), GLU208 (1.71, 2.01 Å), ASN210 (2.18 Å) |
| 31 | Pratensein | −7.4 | LEU141 (2.17 Å), GLY143 (2.16 Å), ASP187 (2.85 Å) | −7.6 | LEU155 (2.09 Å), THR294 (1.90 Å) | −6.9 | LYS621 (2.13 Å), SER795 (2,11, 2.69 Å) | −7.2 | ASN178 (2.16 Å), ASN515 (1.96 Å) | −4.9 | GLU498 (1.83 Å), GLN507 (2.38 Å) | −7.8 | GLN98 (2.72 Å), ASN210 (2.17 Å), GLU564 (1.85 Å) |
| 32 | 2,2-Dimethyl chromene-6-propenoic acid | −7.3 | THR190 (1.76 Å) | −7.3 | ASP157 (2.07 Å) | −6.4 | TRP617 (2.79 Å), ASP761 (1.88 Å) | −6.4 | ASN178 (2.24 Å), THR531 (2.40 Å), ASN515 (2.03, 2.22 Å), ARG559 (2.89 Å) | −5.6 | TYR467 (1.84 Å) | −7.2 | GLU564 (1.95 Å) |
| 33 | Neovestitol | −7.2 | GLU166 (2.03 Å) | −7.1 | ARG159 (2.01 Å), TYR261 (2.11 Å) | −5.9 | LYS798 (2.13 Å) | −6.8 | ALA416 (2.16 Å), ARG408 (2.27 Å) | −5.5 | SER508 (2.09 Å) | −7.4 | ASN210 (1.80, 2.04 Å) |
| 34 | Rutin | −7.2 | ASP187 (1.90 Å), HIS164 (2.25 Å), GLU166 (2.05 Å), GLN192 (2.86 Å), THR190 (2.19, 1.92 Å) | −5.2 | GLU160 (1.95, 1.92 Å), SER163 (2.11 Å) GLU196 (2.12 Å), ARG176 (2.84 Å), MET201 (2.13 Å) | −4.5 | ASP760 (2.23 Å) | −6.7 | THR415 (1.84 Å), THR551 (2.51, 3.01 Å) | −3.3 | SER363 (2.07 Å), TYR365 (1.80 Å), TYR463 (2.27 Å), ASN464 (1.98, 2.63, 3.05 Å) | −9.3 | TYR196 (2.60 Å), ASP206 (2.01, 2.02 Å), GLU208 (2.20, 2.24 Å), ASN210 (2.33, 2.49 Å), TRP566 (2.32 Å) |
| 35 | Medicarpin | −6.5 | GLY143 (2.18 Å), SER144 (2.62 Å), CYS145 (1.83 Å), HIS163 (1.99 Å), | −7.0 | TYR266 (1.76 Å), ASP295 (1.82 Å) | −6.5 | ASP761 (2.23 Å), HIS810 (2.17 Å), SER814 (2.02 Å) | −7.3 | ARG408 (2.10 Å), THR415 (1.90 Å), LEU 416 (2.05 Å), ARG559 (1.89, 2.63 Å) | −5.6 | ARG417 (2.14 Å), GLN507 (2.33 Å), SER508 (2.08 Å), GLY510 (1.97 Å) | −7.7 | LYS94 (2.55 Å) |
| 36 | Ferulic acid | −5.9 | CYS44 (2.48 Å), TYR54 (2.20, 2.44 Å), GLU166 (2.25 Å), ASP187 (2.01 Å) | −5.3 | ASP157 (1.88 Å) ASN260 (2.13 Å) | −4.9 | TRP617 (2.77 Å), ASP761 (1.83 Å) | −6.1 | LYS201 (2.11, 2.79 Å) SER484 (1.87 Å), ASN515 (1.94 Å) | −5.0 | GLU420 (1.62 Å), GLY510 (2.12 Å) | −6.1 | ASN210 (1.76, 1.83 Å), ALA396 (1.88 Å) |
| 37 | m-Coumaric acid | −5.8 | CYS44 (1.74 Å), TYR54 (2.01 Å), HIS164 (2.02 Å), ASP187 (1.82 Å) | −5.0 | ARG159 (2.01 Å), ASN260 (1.75 Å), TYR261 (1.85 Å) | −5.5 | ASP760 (1.95 Å), ASP761 (1.99 Å) | −6.8 | LYS201 (2.11 Å), ASP482 (1.96 Å), SER485 (1.77 Å), ASN515 (1.92 Å) | −5.0 | ASP481 (1.78 Å), GLU485 (1.83 Å) | −6.0 | ASN210 (1.78 Å), ALA396 (1.86 Å), TRP566 (2.23 Å) |
| 38 | p-Coumaric acid | −5.8 | CYS44 (1.73 Å), TYR54 (2.04 Å), HIS164 (2.03 Å), ASP187 (1.83 Å) | −5.2 | ASP157 (1.99 Å), ASN260 (1.89 Å) | −4.9 | ASP761 (1.81 Å), TRP800 (2.45 Å) | −5.3 | ASN176 (2.88 Å), SER485 (2.06 Å) | −4.3 | TYR467 (2.06 Å), SER508 (1.78Å), TYR519 (1.78 Å) | −5.5 | ASN210 (2.05 Å), ALA396 (1.82 Å), TRP566 (2.22 Å) |
| 39 | Caffeic acid | −5.7 | MET49 (1.95 Å), TYR54 (2.15 Å), GLU166 (2.05, 2.19 Å) | −4.9 | ASN260 (1.79 Å), TYR266 (2.30 Å), THR294 (1.78, 1.90 Å) | −5.2 | TRP617 (3.09 Å), ASP761 (1.73, 1.89 Å), HIS810 (2.07 Å), GLU811 (1.84 Å) | −6.5 | LYS201 (2.11 Å), ASP482 (1.95 Å), SER485 (1.79 Å), ASN515 (1.81 Å) | −4.4 | ARG417 (2.38 Å), GLU420 (1.72 Å), GLY510 (1.93, 2.04 Å) | −6.3 | ASN210 (1.96, 1.99 Å), ALA396 (1.82 Å), TRP566 (2.24 Å) |
| 40 | Trans-Cinnamic acid | −5.5 | CYS44 (1.74 Å), TYR54 (2.01 Å), ASP187 (1.82 Å) | −5.0 | ARG159 (1.78, 2.10 Å), TYR261 (1.97 Å) | −5.1 | TRP617 (2.73 Å), ASP761 (1.81 Å), TRP800 (2.42 Å) | −5.0 | ASN176 (2.57 Å), SER485 (1.82 Å) | −4.7 | TYR467 (1.87 Å), SER508 (1.81 Å) | −5.5 | ASN210 (1.94 Å) |
| Compound Name | MW a | Log P b | TPSA c | nON d | nOHNH e | RBs f | BBB+ g | Caco2+ h | HIA+ i | AMES Toxicity | Carcinogenicity |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Retusapurpurin A | 522.6 | 6.2 | 98.4 | 7 | 2 | 5 | 0.41 | 74.84 | 98.97 | Non toxic | Non carcinogenic |
| Baccharin | 318.46 | 3.16 | 77.75 | 3 | 3 | 7 | 0.87 | 57.33 | 97.10 | Non toxic | Non carcinogenic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yosri, N.; Abd El-Wahed, A.A.; Ghonaim, R.; Khattab, O.M.; Sabry, A.; Ibrahim, M.A.A.; Moustafa, M.F.; Guo, Z.; Zou, X.; Algethami, A.F.M.; et al. Anti-Viral and Immunomodulatory Properties of Propolis: Chemical Diversity, Pharmacological Properties, Preclinical and Clinical Applications, and In Silico Potential against SARS-CoV-2. Foods 2021, 10, 1776. https://doi.org/10.3390/foods10081776
Yosri N, Abd El-Wahed AA, Ghonaim R, Khattab OM, Sabry A, Ibrahim MAA, Moustafa MF, Guo Z, Zou X, Algethami AFM, et al. Anti-Viral and Immunomodulatory Properties of Propolis: Chemical Diversity, Pharmacological Properties, Preclinical and Clinical Applications, and In Silico Potential against SARS-CoV-2. Foods. 2021; 10(8):1776. https://doi.org/10.3390/foods10081776
Chicago/Turabian StyleYosri, Nermeen, Aida A. Abd El-Wahed, Reem Ghonaim, Omar M. Khattab, Aya Sabry, Mahmoud A. A. Ibrahim, Mahmoud F. Moustafa, Zhiming Guo, Xiaobo Zou, Ahmed F. M. Algethami, and et al. 2021. "Anti-Viral and Immunomodulatory Properties of Propolis: Chemical Diversity, Pharmacological Properties, Preclinical and Clinical Applications, and In Silico Potential against SARS-CoV-2" Foods 10, no. 8: 1776. https://doi.org/10.3390/foods10081776
APA StyleYosri, N., Abd El-Wahed, A. A., Ghonaim, R., Khattab, O. M., Sabry, A., Ibrahim, M. A. A., Moustafa, M. F., Guo, Z., Zou, X., Algethami, A. F. M., Masry, S. H. D., AlAjmi, M. F., Afifi, H. S., Khalifa, S. A. M., & El-Seedi, H. R. (2021). Anti-Viral and Immunomodulatory Properties of Propolis: Chemical Diversity, Pharmacological Properties, Preclinical and Clinical Applications, and In Silico Potential against SARS-CoV-2. Foods, 10(8), 1776. https://doi.org/10.3390/foods10081776