
Effective Connectivity in the Human Brain for Sour Taste, Retronasal Smell, and Combined Flavour

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Stimulus Solutions and Delivery
2.3. Experimental Procedures
2.3.1. Training Outside the MRI Unit
2.3.2. Sensory Evaluation of Taste, Smell, and Flavour before fMRI Scanning
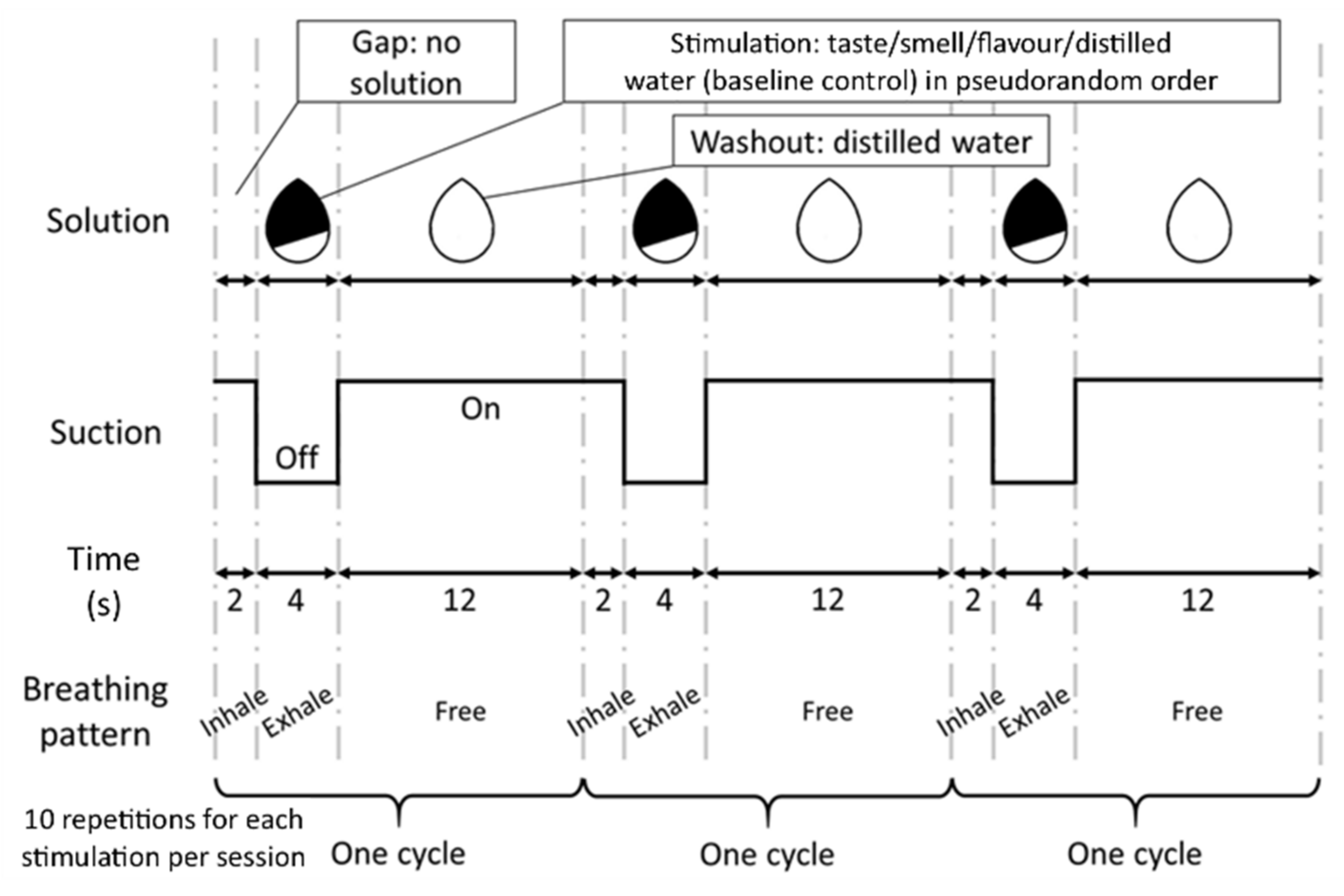
2.3.3. fMRI Study Design
2.3.4. fMRI Data Acquisition
2.4. Data Analysis
2.4.1. Sensory Evaluation
2.4.2. fMRI
fMRI Pre-Processing
Brain Mapping by Whole-Brain Analysis: Conventional General Linear Model Analysis
Effective Connectivity Analysis Using Dynamic Causal Modelling
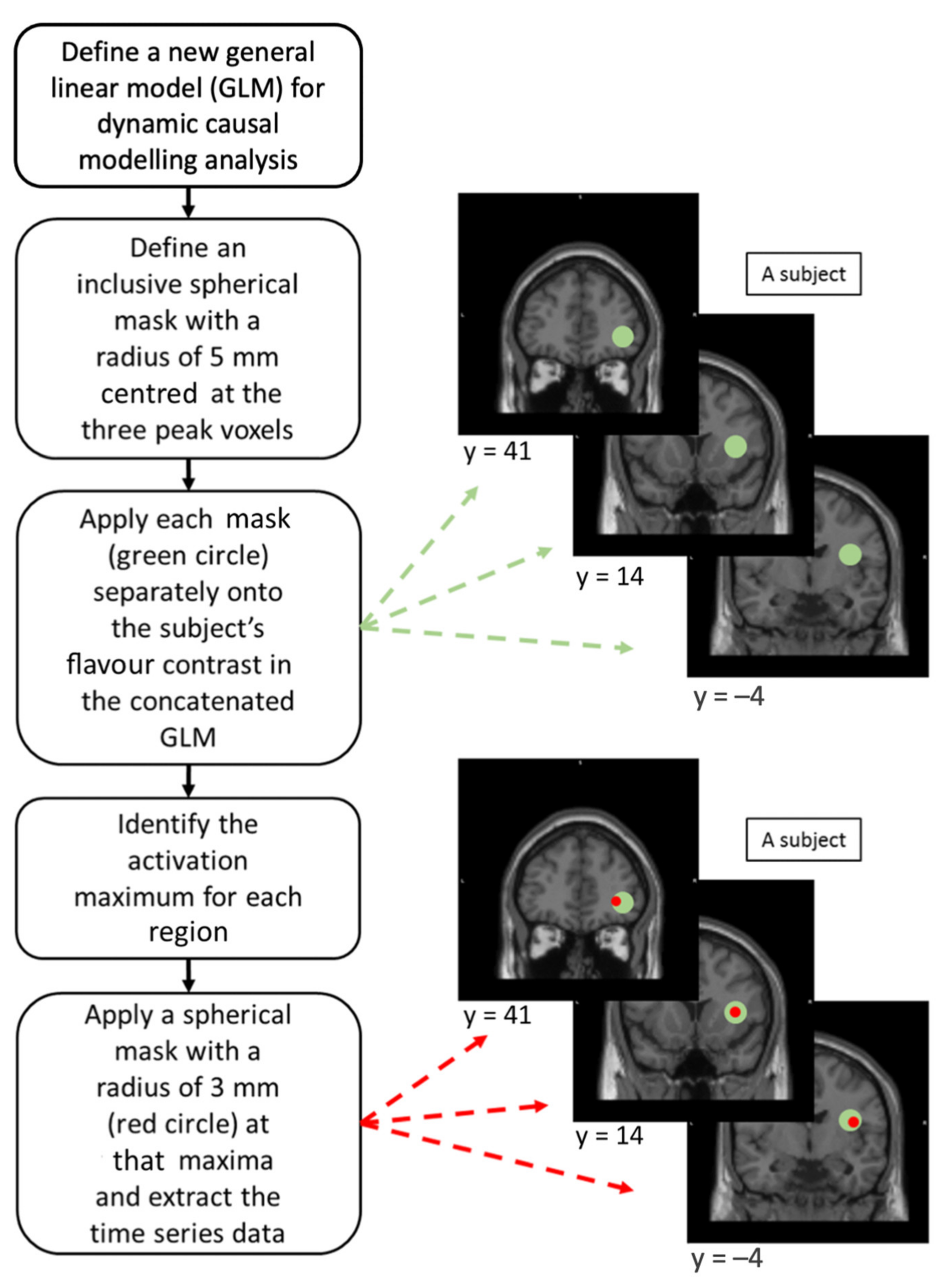
Definition and Extraction of Volume of Interest
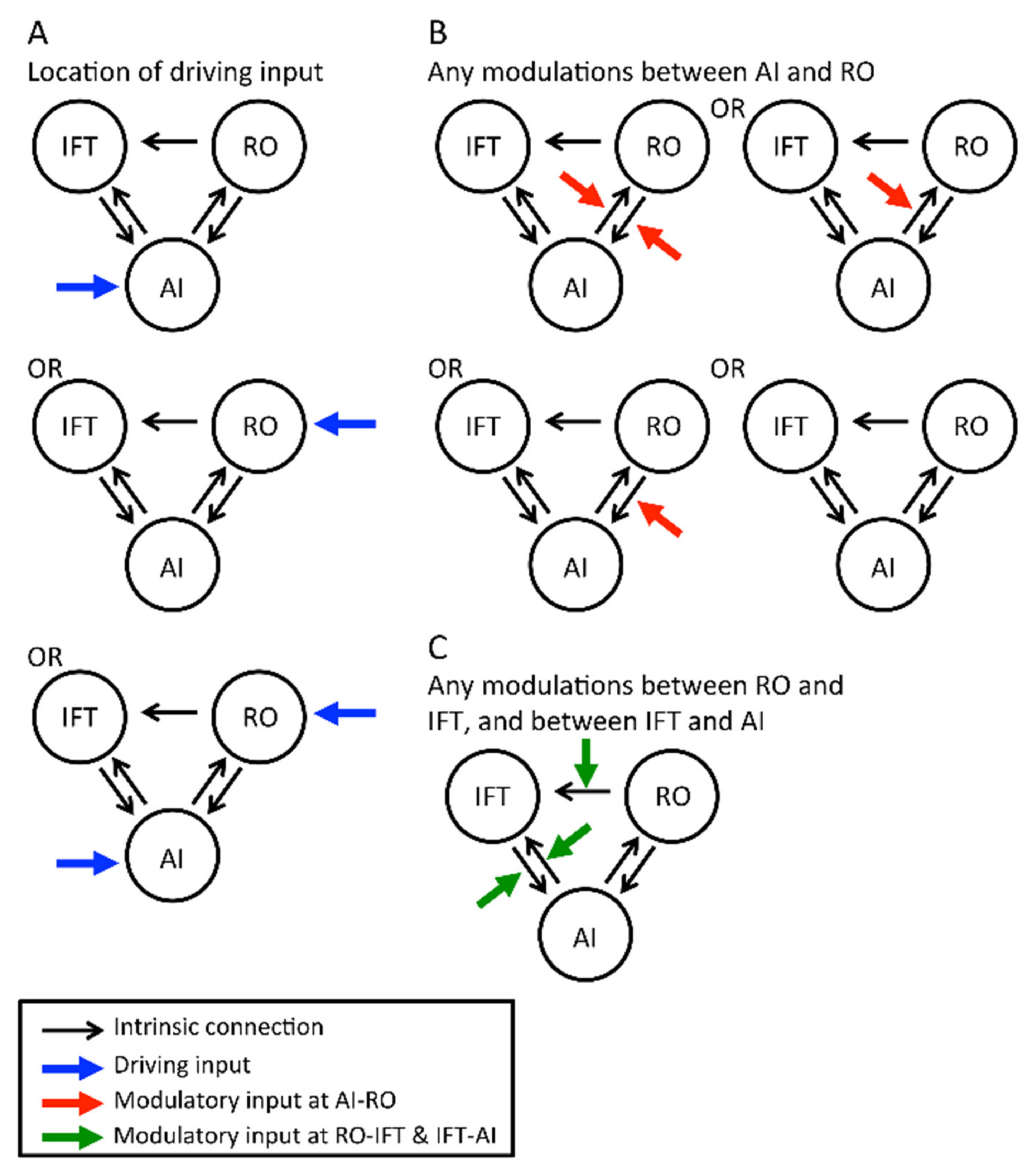
Definition of Intrinsic Connections
Definition of Driving Inputs
Definition of Modulatory Inputs
Inference on Model Structure Using Bayesian Model Selection (BMS)
3. Results
3.1. Sensory Evaluation of Taste, Smell, and Flavour before fMRI
3.2. fMRI Results
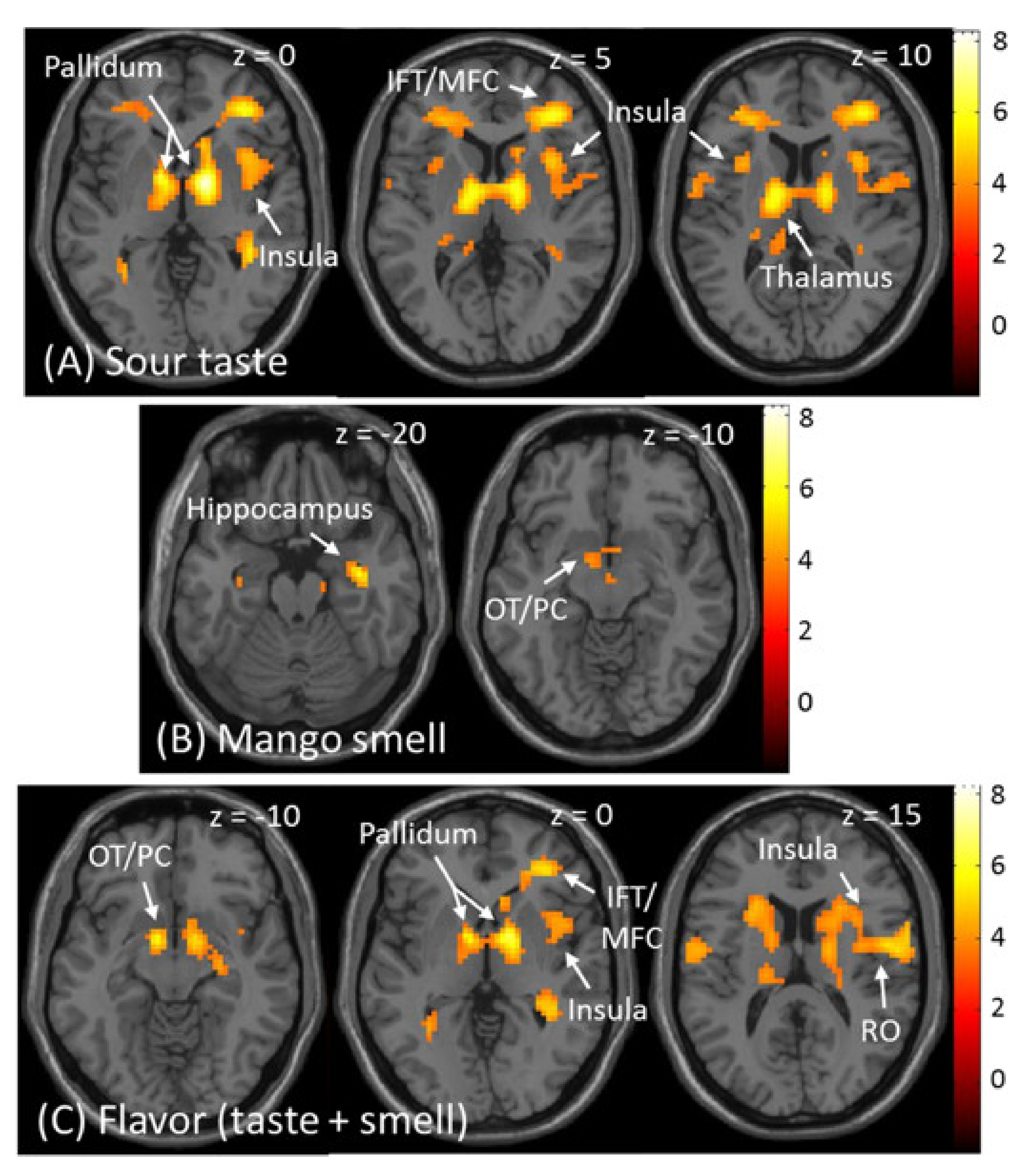
3.2.1. Brain Response to Taste, Smell, and Flavour
3.2.2. Effective Connectivity from DCM Analysis
4. Discussion
4.1. Sensory Evaluation of Taste, Smell, and Flavour
4.2. Brain Activation Maps for Taste, Smell, and Flavour
4.3. Sour Taste
4.4. Mango Smell
4.5. Combined Flavour of Sour Taste and Mango Smell
4.6. Effective Connectivity in the Flavour-Processing Network
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Additional Information
References
- Huerta, C.I.; Sarkar, P.R.; Duong, T.Q.; Laird, A.R.; Fox, P.T. Neural bases of food perception: Coordinate-based meta-analyses of neuroimaging studies in multiple modalities. Obesity 2014, 22, 1439–1446. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Goto, T.K.; Leung, W.K. Basic taste processing recruits bilateral anteroventral and middle dorsal insulae: An activation likelihood estimation meta-analysis of fMRI studies. Brain Behav. 2017, 7, e00655. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Goto, T.K.; Leung, W.K. Affective value, intensity and quality of liquid tastants/food discernment in the human brain: An activation likelihood estimation meta-analysis. Neuroimage 2018, 169, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Veldhuizen, M.G.; Albrecht, J.; Zelano, C.; Boesveldt, S.; Breslin, P.; Lundström, J.N. Identification of human gustatory cortex by activation likelihood estimation. Hum. Brain Mapp. 2011, 32, 2256–2266. [Google Scholar] [CrossRef] [Green Version]
- Perdigón, G.; Fuller, R.; Raya, R. Lactic acid bacteria and their effect on the immune system. Curr. Issues Intest. Microbiol. 2001, 2, 27–42. [Google Scholar]
- Blando, F.; Gerardi, C.; Nicoletti, I. Sour cherry (Prunus cerasus L.) anthocyanins as ingredients for functional foods. Biomed. Res. Int. 2004, 2004, 253–258. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Nair, M.G.; Strasburg, G.M.; Chang, Y.-C.; Booren, A.M.; Gray, J.I.; DeWitt, D.L. Antioxidant and antiinflammatory activities of anthocyanins and their aglycon, cyanidin, from tart cherries. J. Nat. Prod. 1999, 62, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Jezzini, A.; Mazzucato, L.; La Camera, G.; Fontanini, A. Processing of hedonic and chemosensory features of taste in medial prefrontal and insular networks. J. Neurosci. 2013, 33, 18966–18978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaskan, P.M.; Dean, A.M.; Nicholas, M.A.; Mitz, A.R.; Murray, E.A. Gustatory responses in macaque monkeys revealed with fMRI: Comments on taste, taste preference, and internal state. Neuroimage 2019, 184, 932–942. [Google Scholar] [CrossRef] [PubMed]
- Castriota-Scanderbeg, A.; Hagberg, G.E.; Cerasa, A.; Committeri, G.; Galati, G.; Patria, F.; Pitzalis, S.; Caltagirone, C.; Frackowiak, R. The appreciation of wine by sommeliers: A functional magnetic resonance study of sensory integration. Neuroimage 2005, 25, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.; Quiñones, I.; Veldhuizen, M.; Alava, J.-I.; Small, D.; Carreiras, M. What can the brain teach us about winemaking? An fMRI study of alcohol level preferences. PLoS ONE 2015, 10, e0119220. [Google Scholar] [CrossRef]
- Pazart, L.; Comte, A.; Magnin, E.; Millot, J.-L.; Moulin, T. An fMRI study on the influence of sommeliers’ expertise on the integration of flavor. Front. Behav. Neurosci. 2014, 8, 358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, D.M.; Jones-Gotman, M.; Zatorre, R.J.; Petrides, M.; Evans, A.C. A role for the right anterior temporal lobe in taste quality recognition. J. Neurosci. 1997, 17, 5136–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoenfeld, M.; Neuer, G.; Tempelmann, C.; Schüssler, K.; Noesselt, T.; Hopf, J.-M.; Heinze, H.-J. Functional magnetic resonance tomography correlates of taste perception in the human primary taste cortex. Neuroscience 2004, 127, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Haase, L.; Cerf-Ducastel, B.; Murphy, C. Cortical activation in response to pure taste stimuli during the physiological states of hunger and satiety. Neuroimage 2009, 44, 1008–1021. [Google Scholar] [CrossRef] [Green Version]
- Crouzet, S.M.; Busch, N.A.; Ohla, K. Taste quality decoding parallels taste sensations. Curr. Biol. 2015, 25, 890–896. [Google Scholar] [CrossRef] [Green Version]
- Avery, J.A.; Liu, A.G.; Ingeholm, J.E.; Riddell, C.D.; Gotts, S.J.; Martin, A. Taste quality representation in the human brain. J. Neurosci. 2020, 40, 1042–1052. [Google Scholar] [CrossRef]
- Porcu, E.; Benz, K.M.; Ball, F.; Tempelmann, C.; Hanke, M.; Noesselt, T. Macroscopic information-based taste representations in insular cortex are shaped by stimulus concentration. Proc. Natl. Acad. Sci. USA 2020, 117, 7409–7417. [Google Scholar] [CrossRef]
- Murphy, K.; Garavan, H. An empirical investigation into the number of subjects required for an event-related fMRI study. Neuroimage 2004, 22, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Thirion, B.; Pinel, P.; Mériaux, S.; Roche, A.; Dehaene, S.; Poline, J.-B. Analysis of a large fMRI cohort: Statistical and methodological issues for group analyses. Neuroimage 2007, 35, 105–120. [Google Scholar] [CrossRef] [Green Version]
- Zandbelt, B.B.; Gladwin, T.E.; Raemaekers, M.; van Buuren, M.; Neggers, S.F.; Kahn, R.S.; Ramsey, N.F.; Vink, M. Within-subject variation in BOLD-fMRI signal changes across repeated measurements: Quantification and implications for sample size. Neuroimage 2008, 42, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Cerf-Ducastel, B.; Murphy, C. fMRI activation in response to odorants orally delivered in aqueous solutions. Chem. Senses 2001, 26, 625–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozin, P. “Taste-smell confusions” and the duality of the olfactory sense. Atten. Percept. Psychophys. 1982, 31, 397–401. [Google Scholar] [CrossRef] [Green Version]
- Small, D.M.; Gerber, J.C.; Mak, Y.E.; Hummel, T. Differential neural responses evoked by orthonasal versus retronasal odorant perception in humans. Neuron 2005, 47, 593–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottfried, J.A. Central mechanisms of odour object perception. Nat. Rev. Neurosci. 2010, 11, 628–641. [Google Scholar] [CrossRef] [PubMed]
- Seubert, J.; Freiherr, J.; Djordjevic, J.; Lundström, J.N. Statistical localization of human olfactory cortex. Neuroimage 2013, 66, 333–342. [Google Scholar] [CrossRef]
- Sezille, C.; Messaoudi, B.; Bertrand, A.; Joussain, P.; Thevenet, M.; Bensafi, M. A portable experimental apparatus for human olfactory fMRI experiments. J. Neurosci. Methods 2013, 218, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Mohebbi, M.; Seo, H.-S.; Hummel, T. Sensitivity to sweetness correlates to elevated reward brain responses to sweet and high-fat food odors in young healthy volunteers. Neuroimage 2020, 208, 116413. [Google Scholar] [CrossRef]
- Small, D.M.; Jones-Gotman, M.; Zatorre, R.J.; Petrides, M.; Evans, A.C. Flavor processing: More than the sum of its parts. Neuroreport 1997, 8, 3913–3917. [Google Scholar] [CrossRef] [PubMed]
- Gotow, N.; Omata, T.; Uchida, M.; Matsuzaki, N.; Takata, S.; Hagiwara, I.; Kobayakawa, T. Multi-sip time–intensity evaluation of retronasal aroma after swallowing oolong tea beverage. Foods 2018, 7, 177. [Google Scholar] [CrossRef] [Green Version]
- De Araujo, I.E.; Rolls, E.T.; Kringelbach, M.L.; McGlone, F.; Phillips, N. Taste-olfactory convergence, and the representation of the pleasantness of flavour, in the human brain. Eur. J. Neurosci. 2003, 18, 2059–2068. [Google Scholar] [CrossRef] [Green Version]
- McCabe, C.; Rolls, E.T. Umami: A delicious flavor formed by convergence of taste and olfactory pathways in the human brain. Eur. J. Neurosci. 2007, 25, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Seubert, J.; Ohla, K.; Yokomukai, Y.; Kellermann, T.; Lundström, J.N. Superadditive opercular activation to food flavor is mediated by enhanced temporal and limbic coupling. Hum. Brain Mapp. 2015, 36, 1662–1676. [Google Scholar] [CrossRef]
- Shepherd, G.M. Neuroenology: How the brain creates the taste of wine. Flavour 2015, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Small, D.M.; Voss, J.; Mak, Y.E.; Simmons, K.B.; Parrish, T.; Gitelman, D. Experience-dependent neural integration of taste and smell in the human brain. J. Neurophysiol. 2004, 92, 1892–1903. [Google Scholar] [CrossRef]
- Friston, K.J.; Harrison, L.; Penny, W. Dynamic causal modelling. Neuroimage 2003, 19, 1273–1302. [Google Scholar] [CrossRef]
- Nakamura, Y.; Tokumori, K.; Tanabe, H.C.; Yoshiura, T.; Kobayashi, K.; Nakamura, Y.; Honda, H.; Yoshiura, K.; Goto, T.K. Localization of the primary taste cortex by contrasting passive and attentive conditions. Exp. Brain Res. 2013, 227, 185–197. [Google Scholar] [CrossRef]
- Yeung, A.W.K.; Tanabe, H.C.; Suen, J.L.K.; Goto, T.K. Taste intensity modulates effective connectivity from the insular cortex to the thalamus in humans. Neuroimage 2016, 135, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Iannilli, E.; Singh, P.B.; Schuster, B.; Gerber, J.; Hummel, T. Taste laterality studied by means of umami and salt stimuli: An fMRI study. Neuroimage 2012, 60, 426–435. [Google Scholar] [CrossRef]
- Veldhuizen, M.G.; Douglas, D.; Aschenbrenner, K.; Gitelman, D.R.; Small, D.M. The anterior insular cortex represents breaches of taste identity expectation. J. Neurosci. 2011, 31, 14735–14744. [Google Scholar] [CrossRef] [Green Version]
- Dalton, P.; Doolittle, N.; Nagata, H.; Breslin, P. The merging of the senses: Integration of subthreshold taste and smell. Nat. Neurosci. 2000, 3, 431–432. [Google Scholar] [CrossRef] [PubMed]
- Malundo, T.; Shewfelt, R.; Ware, G.; Baldwin, E. Sugars and acids influence flavor properties of mango (Mangifera indica). J. Am. Soc. Hortic. Sci. 2001, 126, 115–121. [Google Scholar] [CrossRef] [Green Version]
- Saha, P.; SIngh, J.P.; Sourav, S.; Humayun, A.; Ramalingam, C. Optimization of citric acid and malic acid to enhance flavour and shelf life of mango juice. J. Chem. Pharm. Res. 2020, 5, 90–95. [Google Scholar]
- Goto, T.K.; Yeung, A.W.K.; Suen, J.L.K.; Fong, B.S.K.; Ninomiya, Y. High resolution time–intensity recording with synchronized solution delivery system for the human dynamic taste perception. J. Neurosci. Methods 2015, 245, 147–155. [Google Scholar] [CrossRef]
- Goto, T.K.; Yeung, A.W.K.; Tanabe, H.C.; Ito, Y.; Jung, H.-S.; Ninomiya, Y. Enhancement of combined umami and salty taste by glutathione in the human tongue and brain. Chem. Senses 2016, 41, 623–630. [Google Scholar] [CrossRef] [Green Version]
- Pernet, C.R. Misconceptions in the use of the General Linear Model applied to functional MRI: A tutorial for junior neuro-imagers. Front. Neurosci. 2014, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Rolls, E.T. The representation of umami taste in the taste cortex. J. Nutr. 2000, 130, 960S–965S. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. Functional and effective connectivity in neuroimaging: A synthesis. Hum. Brain Mapp. 1994, 2, 56–78. [Google Scholar] [CrossRef]
- Maldjian, J.A.; Laurienti, P.J.; Kraft, R.A.; Burdette, J.H. An automated method for neuroanatomic and cytoarchitectonic atlas-based interrogation of fMRI data sets. Neuroimage 2003, 19, 1233–1239. [Google Scholar] [CrossRef]
- Kahnt, T.; Chang, L.J.; Park, S.Q.; Heinzle, J.; Haynes, J.-D. Connectivity-based parcellation of the human orbitofrontal cortex. J. Neurosci. 2012, 32, 6240–6250. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, H.C.; Sakai, T.; Morito, Y.; Kochiyama, T.; Sadato, N. Neural correlates and effective connectivity of subjective colors during the Benham’s top illusion: A functional MRI study. Cereb. Cortex 2011, 21, 124–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustine, J.R. Circuitry and functional aspects of the insular lobe in primates including humans. Brain Res. Rev. 1996, 22, 229–244. [Google Scholar] [CrossRef]
- Catani, M.; Dell’Acqua, F.; Vergani, F.; Malik, F.; Hodge, H.; Roy, P.; Valabregue, R.; De Schotten, M.T. Short frontal lobe connections of the human brain. Cortex 2012, 48, 273–291. [Google Scholar] [CrossRef] [PubMed]
- Frank, R.A.; Byram, J. Taste–smell interactions are tastant and odorant dependent. Chem. Senses 1988, 13, 445–455. [Google Scholar] [CrossRef]
- Green, B.G.; Nachtigal, D.; Hammond, S.; Lim, J. Enhancement of retronasal odors by taste. Chem. Senses 2012, 37, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, R.J.; Prescott, J.; Boakes, R.A. Confusing tastes and smells: How odours can influence the perception of sweet and sour tastes. Chem. Senses 1999, 24, 627–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephan, K.E.; Penny, W.D.; Moran, R.J.; den Ouden, H.E.; Daunizeau, J.; Friston, K.J. Ten simple rules for dynamic causal modeling. Neuroimage 2010, 49, 3099–3109. [Google Scholar] [CrossRef] [PubMed]
- Besser, G.; Liu, D.T.; Renner, B.; Hummel, T.; Mueller, C.A. Reversible obstruction of the olfactory cleft: Impact on olfactory perception and nasal patency. Int. Forum Allergy Rhinol. 2020, 10, 713–718. [Google Scholar] [CrossRef] [PubMed]
- Auvray, M.; Spence, C. The multisensory perception of flavor. Conscious. Cogn. 2008, 17, 1016–1031. [Google Scholar] [CrossRef]
- Prescott, J. Multimodal chemosensory interactions and perception of flavor. In The Neural Bases of Multisensory Processes; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2012. [Google Scholar]
- Okamoto, M.; Dan, I. Extrinsic information influences taste and flavor perception: A review from psychological and neuroimaging perspectives. Semin. Cell Dev. Biol. 2013, 24, 247–255. [Google Scholar] [CrossRef]
- Royet, J.-P.; Delon-Martin, C.; Plailly, J. Odor mental imagery in non-experts in odors: A paradox? Front. Hum. Neurosci. 2013, 7, 87. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, S.; Kubota, F.; Nisijima, K.; Washiya, S.; Kato, S. Cerebral activation focusing on strong tasting food: A functional magnetic resonance imaging study. Neuroreport 2005, 16, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Bensafi, M.; Croy, I.; Phillips, N.; Rouby, C.; Sezille, C.; Gerber, J.; Small, D.M.; Hummel, T. The effect of verbal context on olfactory neural responses. Hum. Brain Mapp. 2014, 35, 810–818. [Google Scholar] [CrossRef]
- Royet, J.-P.; Plailly, J.; Saive, A.-L.; Veyrac, A.; Delon-Martin, C. The impact of expertise in olfaction. Front. Psychol. 2013, 4, 928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolls, E.T. Functions of the anterior insula in taste, autonomic, and related functions. Brain Cogn. 2016, 110, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, M.D.; Cunningham, W.A. Type I and Type II error concerns in fMRI research: Re-balancing the scale. Soc. Cogn. Affect. Neurosci. 2009, 4, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Kami, Y.N.; Goto, T.K.; Tokumori, K.; Yoshiura, T.; Kobayashi, K.; Nakamura, Y.; Honda, H.; Ninomiya, Y.; Yoshiura, K. The development of a novel automated taste stimulus delivery system for fMRI studies on the human cortical segregation of taste. J. Neurosci. Methods 2008, 172, 48–53. [Google Scholar] [CrossRef]
- Kobayakawa, T.; Ogawa, H.; Kaneda, H.; Ayabe-Kanamura, S.; Saito, S. Spatio-temporal analysis of cortical activity evoked by gustatory stimulation in humans. Chem. Senses 1999, 24, 201–209. [Google Scholar] [CrossRef] [Green Version]
- Kobayakawa, T.; Saito, S.; Gotow, N.; Ogawa, H. Representation of salty taste stimulus concentrations in the primary gustatory area in humans. Chemosens. Percept. 2008, 1, 227–234. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Goto, T.K.; Tokumori, K.; Yoshiura, T.; Kobayashi, K.; Nakamura, Y.; Honda, H.; Ninomiya, Y.; Yoshiura, K. Localization of brain activation by umami taste in humans. Brain Res. 2011, 1406, 18–29. [Google Scholar] [CrossRef]
- Small, D.M.; Zald, D.H.; Jones-Gotman, M.; Zatorre, R.J.; Pardo, J.V.; Frey, S.; Petrides, M. Human cortical gustatory areas: A review of functional neuroimaging data. Neuroreport 1999, 10, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Grabenhorst, F.; Rolls, E.T. Selective attention to affective value alters how the brain processes taste stimuli. Eur. J. Neurosci. 2008, 27, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Veldhuizen, M.G.; Bender, G.; Constable, R.T.; Small, D.M. Trying to detect taste in a tasteless solution: Modulation of early gustatory cortex by attention to taste. Chem. Senses 2007, 32, 569–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelano, C.; Bensafi, M.; Porter, J.; Mainland, J.; Johnson, B.; Bremner, E.; Telles, C.; Khan, R.; Sobel, N. Attentional modulation in human primary olfactory cortex. Nat. Neurosci. 2005, 8, 114–120. [Google Scholar] [CrossRef]
- Kelley, A.E.; Berridge, K.C. The neuroscience of natural rewards: Relevance to addictive drugs. J. Neurosci. 2002, 22, 3306–3311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kringelbach, M.L.; Berridge, K.C. Pleasures of the Brain; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Saper, C.B.; Chou, T.C.; Elmquist, J.K. The need to feed: Homeostatic and hedonic control of eating. Neuron 2002, 36, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Fellows, L.; Small, D.; Dagher, A. Food and drug cues activate similar brain regions: A meta-analysis of functional MRI studies. Physiol. Behav. 2012, 106, 317–324. [Google Scholar] [CrossRef]
- Carmichael, S.T.; Clugnet, M.C.; Price, J.L. Central olfactory connections in the macaque monkey. J. Comp. Neurol. 1994, 346, 403–434. [Google Scholar] [CrossRef]
- Sobel, N.; Prabhakaran, V.; Zhao, Z.; Desmond, J.E.; Glover, G.H.; Sullivan, E.V.; Gabrieli, J.D. Time course of odorant-induced activation in the human primary olfactory cortex. J. Neurophysiol. 2000, 83, 537–551. [Google Scholar] [CrossRef]
- Moser, M.B.; Moser, E.I. Functional differentiation in the hippocampus. Hippocampus 1998, 8, 608–619. [Google Scholar] [CrossRef]
- Tulving, E.; Markowitsch, H.J. Episodic and declarative memory: Role of the hippocampus. Hippocampus 1998, 8, 198–204. [Google Scholar] [CrossRef]
- Zeidman, P.; Lutti, A.; Maguire, E.A. Investigating the functions of subregions within anterior hippocampus. Cortex 2015, 73, 240–256. [Google Scholar] [CrossRef] [Green Version]
- Sinding, C.; Thibault, H.; Hummel, T.; Thomas-Danguin, T. Odor-induced saltiness enhancement: Insights into the brain chronometry of flavor perception. Neuroscience 2021, 452, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Onuma, T.; Maruyama, H.; Sakai, N. Enhancement of saltiness perception by monosodium glutamate taste and soy sauce odor: A near-infrared spectroscopy study. Chem. Senses 2018, 43, 151–167. [Google Scholar] [CrossRef] [Green Version]
- Gotow, N.; Kobayakawa, T. Trial measurement of brain activity underlying olfactory–gustatory synchrony perception using event-related potentials from five female participants. J. Neurosci. Res. 2019, 97, 253–266. [Google Scholar] [CrossRef] [PubMed]
- Fondberg, R.; Lundström, J.N.; Blöchl, M.; Olsson, M.J.; Seubert, J. Multisensory flavor perception: The relationship between congruency, pleasantness, and odor referral to the mouth. Appetite 2018, 125, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Small, D.M. Flavor is in the brain. Physiol. Behav. 2012, 107, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Flynn, F.G. Anatomy of the insula functional and clinical correlates. Aphasiology 1999, 13, 55–78. [Google Scholar] [CrossRef]
- Bertino, M.; Beauchamp, G.K.; Engelman, K. Naltrexone, an opioid blocker, alters taste perception and nutrient intake in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1991, 261, R59–R63. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
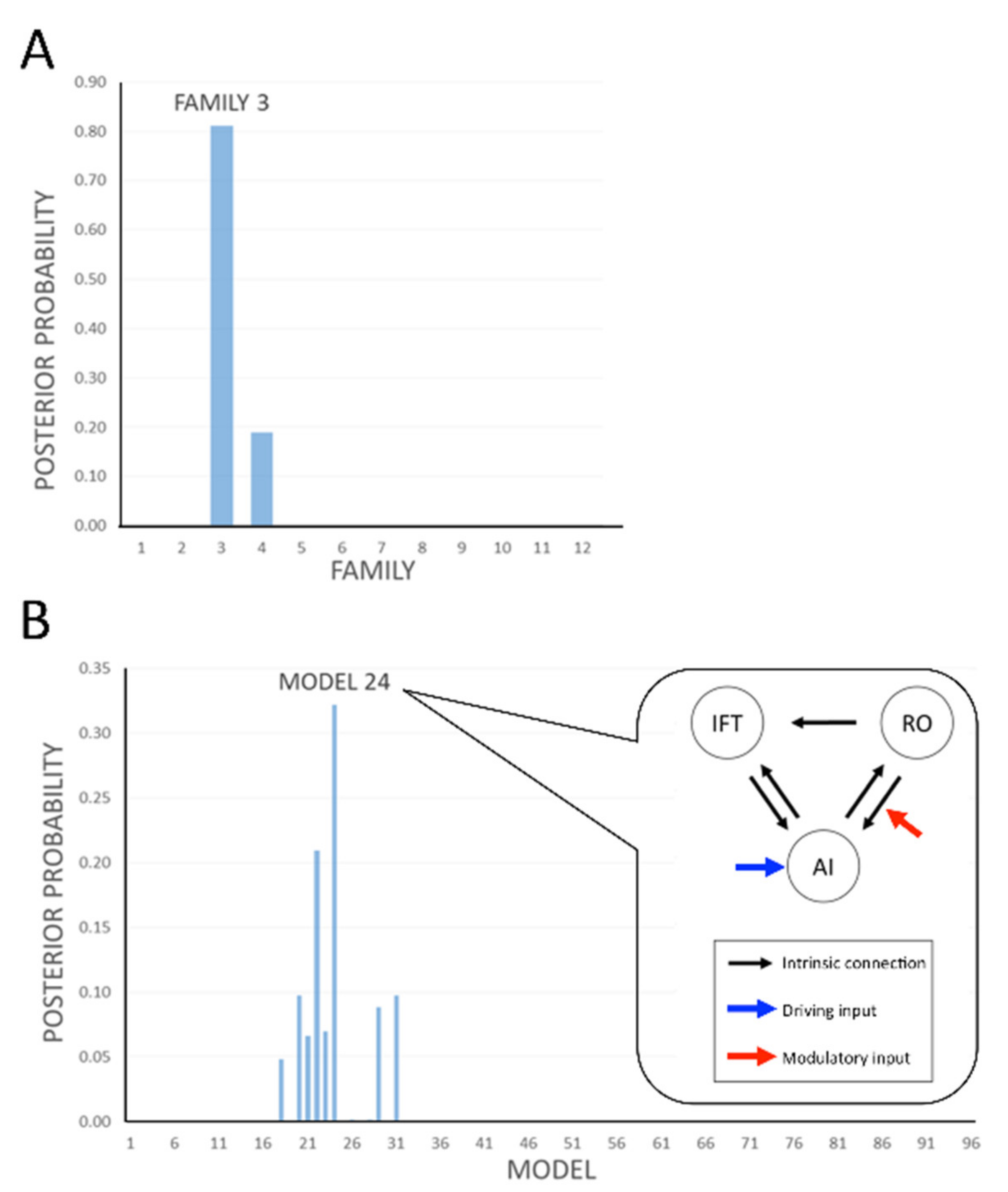{kind=link}
| Modulatory Input (s) c | ||||
|---|---|---|---|---|
| Location of Driving Input b | both AI 🡪 RO and RO 🡪 AI | AI 🡪 RO | RO 🡪 AI | None |
| AI | 1 (M1–8) | 2 (M9–16) | 3 (M17–24) | 4 (M25–32) |
| RO | 5 (M33–40) | 6 (M41–48) | 7 (M49–56) | 8 (M57–64) |
| Both AI and RO | 9 (M65–72) | 10 (M73–80) | 11 (M81–88) | 12 (M89–96) |
| Location | Cluster Size (Voxel) | MNI Coordinate (x,y,z) | t | p-Value a |
|---|---|---|---|---|
| Sour taste | ||||
| R Precentral gyrus | 4463 | 48,−13,34 | 8.23 | <0.001 |
| R Postcentral gyrus | - | 54,−7,25 | 7.60 | <0.001 |
| R Pallidum | - | 15,−1,1 | 7.52 | <0.001 |
| L Precentral gyrus | - | −48,−13,37 | 7.03 | <0.001 |
| L Pallidum | - | −12,2,1 | 6.26 | 0.003 |
| R Inferior frontal triangularis/Middle frontal gyrus | - | 36,41,4 | 6.00 | 0.006 |
| L Thalamus | - | −12,−7,10 | 5.88 | 0.007 |
| L Middle cingulate cortex | - | −6,−25,28 | 5.64 | 0.013 |
| R Superior medial frontal gyrus/Anterior cingulate cortex | - | 15,14,34 | 5.59 | 0.015 |
| R Middle cingulate cortex | - | 9,−22,31 | 5.57 | 0.016 |
| L Anterior cingulate cortex | - | −9,26,22 | 5.42 | 0.023 |
| R Supramarginal gyrus | - | 30,−40,31 | 5.23 | 0.037 |
| R Middle temporal gyrus | - | 39,−40,1 | 5.17 | 0.043 |
| L Superior medial frontal gyrus | - | −9,11,37 | 5.13 | 0.047 |
| L Anterior insula | - | −30,14,13 | 4.97 | <0.001 b |
| R Anterior insula | - | 36,14,7 | 4.70 | <0.001 b |
| R Anterior insula | - | 39,11,1 | 4.47 | <0.001 b |
| R Middle insula | - | 42,8,−2 | 4.45 | <0.001 b |
| R Cerebellum | 565 | 12,−58,−23 | 6.70 | 0.001 |
| Vermis | - | 0,−43,−20 | 6.12 | 0.004 |
| R Vermis | - | 3,−52,−26 | 6.10 | 0.004 |
| L Cerebellum | - | −15,−58,−23 | 5.89 | 0.007 |
| Vermis | - | 0,−67,−17 | 5.18 | 0.042 |
| Mango smell | ||||
| R Hippocampus | 55 | 36,−10,−20 | 5.59 | 0.014 |
| R Hippocampus | - | 39,−16,−17 | 5.04 | <0.001 b |
| L Olfactory tubercle/Piriform | 109 | −9,−4,−11 | 4.57 | <0.001 b |
| R Pallidum | - | 12,5,−2 | 4.36 | <0.001 b |
| R Pallidum | - | 12,−1,−2 | 4.16 | <0.001 b |
| Flavour (sour taste plus mango smell) | ||||
| R Precentral gyrus | 2331 | 48,−10,34 | 7.05 | <0.001 |
| R Precentral gyrus | - | 60,2,28 | 6.42 | 0.002 |
| R Pallidum | - | 12,2,1 | 6.29 | 0.002 |
| L Pallidum | - | −12,2,−2 | 5.90 | 0.006 |
| L Olfactory tubercle/Piriform | - | −9,−1,−8 | 5.42 | 0.021 |
| R Superior medial frontal gyrus | - | 6,2,64 | 5.12 | 0.042 |
| R Anterior insula | - | 33,14,13 | 4.95 | <0.001 b |
| R Rolandic operculum | - | 39,−4,19 | 4.70 | <0.001 b |
| R Middle insula | - | 39,−1,4 | 3.92 | <0.001 b |
| R Cerebellum | 425 | 12,−58,−23 | 6.82 | 0.001 |
| L Cerebellum | - | −12,−61,−23 | 6.34 | 0.002 |
| L Postcentral gyrus | 310 | −48,−16,31 | 6.26 | 0.003 |
| L Postcentral gyrus | - | −57,−13,28 | 6.06 | 0.004 |
| L Middle cingulate gyrus | 190 | −3,−25,28 | 5.55 | 0.015 |
| R Inferior frontal triangularis/Middle frontal gyrus | 127 | 33,41,4 | 5.28 | 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suen, J.L.K.; Yeung, A.W.K.; Wu, E.X.; Leung, W.K.; Tanabe, H.C.; Goto, T.K. Effective Connectivity in the Human Brain for Sour Taste, Retronasal Smell, and Combined Flavour. Foods 2021, 10, 2034. https://doi.org/10.3390/foods10092034
Suen JLK, Yeung AWK, Wu EX, Leung WK, Tanabe HC, Goto TK. Effective Connectivity in the Human Brain for Sour Taste, Retronasal Smell, and Combined Flavour. Foods. 2021; 10(9):2034. https://doi.org/10.3390/foods10092034
Chicago/Turabian StyleSuen, Justin Long Kiu, Andy Wai Kan Yeung, Ed X. Wu, Wai Keung Leung, Hiroki C. Tanabe, and Tazuko K. Goto. 2021. "Effective Connectivity in the Human Brain for Sour Taste, Retronasal Smell, and Combined Flavour" Foods 10, no. 9: 2034. https://doi.org/10.3390/foods10092034
APA StyleSuen, J. L. K., Yeung, A. W. K., Wu, E. X., Leung, W. K., Tanabe, H. C., & Goto, T. K. (2021). Effective Connectivity in the Human Brain for Sour Taste, Retronasal Smell, and Combined Flavour. Foods, 10(9), 2034. https://doi.org/10.3390/foods10092034