
Dietary Vegetable Powders Modulate Immune Homeostasis and Intestinal Microbiota in Mice


Abstract
:1. Introduction
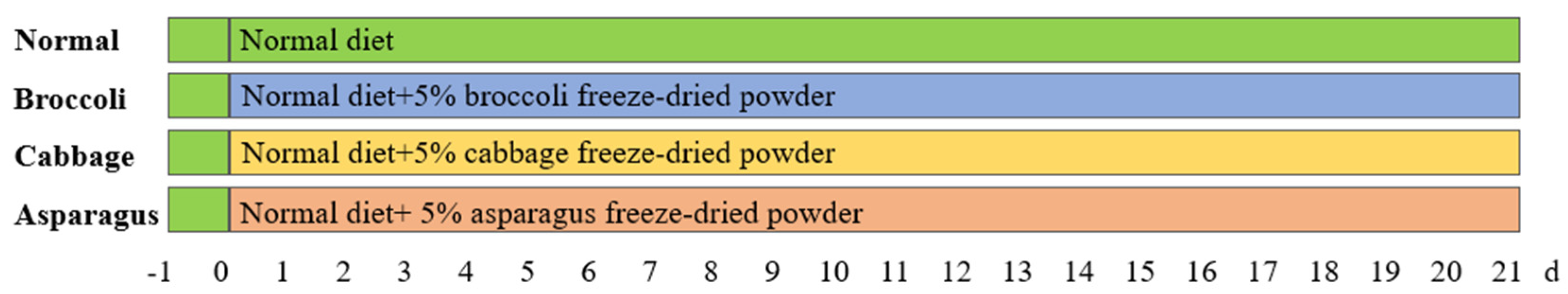
2. Materials and Methods
2.1. Qualification of Average Daily Body Weight Gain (ADG) of Mice
2.2. Determination of Immunoglobulin Concentration
2.3. Determination of Intestinal Cytokine Concentration in Mice
2.4. Quantification of Short-Chain Fatty Acids (SCFAs)
2.5. Determination of Intestinal Flora in Mice
2.6. Statistics Analysis
3. Results
3.1. Calculation of the Average Daily Gain of Mice in Each Group
3.2. Analysis of Immunoglobulin Concentration in Mice
3.3. Analysis of Cytokine Concentration in Jejunum and Ileum of Mice
3.4. Analysis of Short-Chain Fatty Acid Content in Cecum of Mice
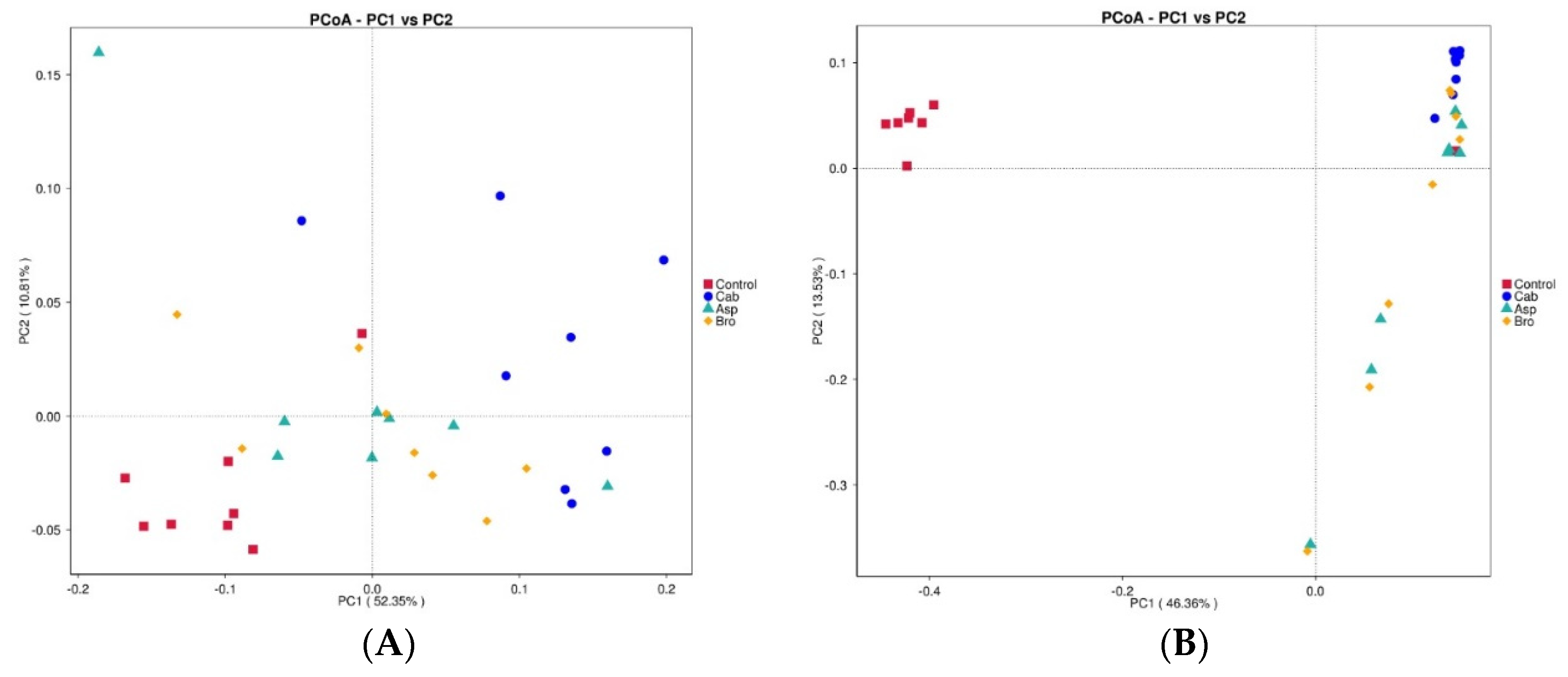
3.5. Analysis of High-Throughput 16S rRNA Gene Sequencing Results
4. Discussion
4.1. Dietary Intake of Vegetable Powder Increased Immunoglobulin Concentration in Mice
4.2. Dietary Intake of Vegetable Powder Changed the Concentration of Cytokines in the Jejunum and Ileum of Mice
4.3. Dietary Intake of Vegetable Powder Changed the Composition and Abundance of Intestinal Flora in Mice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sender, R.; Fuchs, S.; Milo, R. Are We Really Vastly Outnumbered? Revisiting the Ratio of Bacterial to Host Cells in Humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.; Ye, J.; Zhao, B.; Sun, J.; Cao, P.; Yang, Y. The Role of Intestinal Microbiota in Colorectal Cancer. Front. Pharmacol. 2021, 12, 674807. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Ling, Z.; Li, L. The Intestinal Microbiota and Colorectal Cancer. Front. Immunol. 2020, 11, 3100. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Xu, S.; Fang, J.; Jiang, H. The Protective Effect of Polyphenols for Colorectal Cancer. Front. Immunol. 2020, 11, 1407. [Google Scholar] [CrossRef]
- Huang, P.; Liu, Y. A Reasonable Diet Promotes Balance of Intestinal Microbiota: Prevention of Precolorectal Cancer. BioMed Res. Int. 2019, 2019, 3405278. [Google Scholar] [CrossRef]
- Hubbard, T.D.; Murray, I.A.; Nichols, R.; Cassel, K.; Podolsky, M.; Kuzu, G.; Tian, Y.; Smith, P.; Kennett, M.J.; Patterson, A.; et al. Dietary broccoli impacts microbial community structure and attenuates chemically induced colitis in mice in an Ah receptor dependent manner. J. Funct. Foods 2017, 37, 685–698. [Google Scholar] [CrossRef]
- Garcia-Ibañez, P.; Roses, C.; Agudelo, A.; Milagro, F.; Barceló, A.; Viadel, B.; Nieto, J.; Moreno, D.; Carvajal, M. The Influence of Red Cabbage Extract Nanoencapsulated with Brassica Plasma Membrane Vesicles on the Gut Microbiome of Obese Volunteers. Foods 2021, 10, 1038. [Google Scholar] [CrossRef]
- Power, K.A.; Lu, J.T.; Monk, J.M.; Lepp, D.; Wu, W.; Zhang, C.; Liu, R.; Tsao, R.; Robinson, L.E.; Wood, G.A.; et al. Purified rutin and rutin-rich asparagus attenuates disease severity and tissue damage following dextran sodium sulfate-induced colitis. Mol. Nutr. Food Res. 2016, 60, 2396–2412. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, C.Y.; Pace-Templeton, J.G.; Millard, C.B.; Wannemacher, R.W.; Hewetson, J.F. Validation of ELISA for the determination of anti-ricin immunoglobulin G concentration in mouse sera. Biologicals 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Matsuda, T.; Hirano, T. Establishment of the ELISA for Murine-Soluble GP130, a Signal Transducer for the IL-6 Family Cytokine, and Its Detection in the Ascitic Fluids of Tumor-Bearing Mice. Biochem. Biophys. Res. Commun. 1994, 202, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Hu, S.; Ni, K.; Chang, G.; Sun, X.; Yu, L. Characterisation of Fecal Soap Fatty Acids, Calcium Contents, Bacterial Community and Short-Chain Fatty Acids in Sprague Dawley Rats Fed with Different sn-2 Palmitic Triacylglycerols Diets. PLoS ONE 2016, 11, e0164894. [Google Scholar] [CrossRef]
- Han, Y.; Song, M.; Gu, M.; Ren, D.; Zhu, X.; Cao, X.; Li, F.; Wang, W.; Cai, X.; Yuan, B.; et al. Dietary Intake of Whole Strawberry Inhibited Colonic Inflammation in Dextran-Sulfate-Sodium-Treated Mice via Restoring Immune Homeostasis and Alleviating Gut Microbiota Dysbiosis. J. Agric. Food Chem. 2019, 67, 9168–9177. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Misra, B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114. [Google Scholar] [CrossRef]
- Francisco, M.; Tortosa, M.; Ballesta, M.M.; Velasco, P.; Garcia-Viguera, C.; Moreno, D. Nutritional and phytochemical value ofBrassicacrops from the agri-food perspective. Ann. Appl. Biol. 2016, 170, 273–285. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Fan, S.-T.; Huang, D.-F.; Yu, Q.; Liu, X.-Z.; Li, C.; Wang, S.; Xiong, T.; Nie, S.-P.; Xie, M.-Y. Effect of Lactobacillus plantarum NCU116 Fermentation on Asparagus officinalis Polysaccharide: Characterization, Antioxidative, and Immunoregulatory Activities. J. Agric. Food Chem. 2018, 66, 10703–10711. [Google Scholar] [CrossRef]
- Megha, K.; Mohanan, P. Role of immunoglobulin and antibodies in disease management. Int. J. Biol. Macromol. 2021, 169, 28–38. [Google Scholar] [CrossRef]
- Liu, N.; Deng, X.; Liang, C.; Cai, H. Effect of Broccoli Residues Fermented with Probiotics on the Growth Performance and Health Status of Broilers Challenged with Clostridium perfringens. Braz. J. Poult. Sci. 2018, 20, 625–632. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, Y.; Tokunaga, Y.; Takagaki, K.; Tsusaki, S.; Tachibana, H.; Yamada, K. Effect of Dietary Cabbage Fermentation Extract and Young Barley Leaf Powder on Immune Function of Sprague-Dawley Rats. J. Nutr. Sci. Vitaminol. 2001, 47, 253–257. [Google Scholar] [CrossRef] [Green Version]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Gutzeit, C.; Magri, G.; Cerutti, A. Intestinal IgA production and its role in host-microbe interaction. Immunol. Rev. 2014, 260, 76–85. [Google Scholar] [CrossRef]
- Vieira, A.T.; Teixeira, M.M.; Martins, F.D.S. The Role of Probiotics and Prebiotics in Inducing Gut Immunity. Front. Immunol. 2013, 4, 445. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Saulnier, D.; Pot, B.; Versalovic, J. How can probiotics and prebiotics impact mucosal immunity? Gut Microbes 2010, 1, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Xing, J.; Zheng, S.; Bo, R.; Luo, L.; Huang, Y.; Niu, Y.; Li, Z.; Wang, D.; Hu, Y.; et al. Ganoderma lucidum polysaccharides encapsulated in liposome as an adjuvant to promote Th1-bias immune response. Carbohydr. Polym. 2016, 142, 141–148. [Google Scholar] [CrossRef]
- Mantovani, A.; Dinarello, C.A.; Molgora, M.; Garlanda, C. Interleukin-1 and Related Cytokines in the Regulation of Inflammation and Immunity. Immunity 2019, 50, 778–795. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.M.; Shin, D.W.; Lim, B.O. Chlorogenic Acid Improves Symptoms of Inflammatory Bowel Disease in Interleukin-10 Knockout Mice. J. Med. Food 2020, 23, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Yoo, H.J.; Byun, H.-J.; Kim, B.-R.; Lee, K.H.; Park, S.-Y.; Rho, S.B. DAPk1 inhibits NF-κB activation through TNF-α and INF-γ-induced apoptosis. Cell. Signal. 2012, 24, 1471–1477. [Google Scholar] [CrossRef]
- Gupta, K.K.; Khan, M.A.; Singh, S.K. Constitutive Inflammatory Cytokine Storm: A Major Threat to Human Health. J. Interf. Cytokine Res. 2020, 40, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Cai, B.; Wang, J.; Sheng, Z.; Yang, H.; Wang, D.; Chen, J.; Ning, Q. Mulberry leaf-derived polysaccharide modulates the immune response and gut microbiota composition in immunosuppressed mice. J. Funct. Foods 2021, 83, 104545. [Google Scholar] [CrossRef]
- Ding, Y.; Yan, Y.; Chen, D.; Ran, L.; Mi, J.; Lu, L.; Jing, B.; Li, X.; Zeng, X.; Cao, Y. Modulating effects of polysaccharides from the fruits of Lycium barbarumon the immune response and gut microbiota in cyclophosphamide-treated mice. Food Funct. 2019, 10, 3671–3683. [Google Scholar] [CrossRef]
- Hua, M.; Liu, Z.; Sha, J.; Li, S.; Dong, L.; Sun, Y. Effects of ginseng soluble dietary fiber on serum antioxidant status, immune factor levels and cecal health in healthy rats. Food Chem. 2021, 365, 130641. [Google Scholar] [CrossRef]
- Luo, J.; Lin, X.; Bordiga, M.; Brennan, C.; Xu, B. Manipulating effects of fruits and vegetables on gut microbiota—A critical review. Int. J. Food Sci. Technol. 2021, 56, 2055–2067. [Google Scholar] [CrossRef]
- Li, F.; Hullar, M.A.J.; Schwarz, Y.; Lampe, J.W. Human Gut Bacterial Communities Are Altered by Addition of Cruciferous Vegetables to a Controlled Fruit- and Vegetable-Free Diet. J. Nutr. 2009, 139, 1685–1691. [Google Scholar] [CrossRef]
- Zagato, E.; Pozzi, C.; Bertocchi, A.; Schioppa, T.; Saccheri, F.; Guglietta, S.; Fosso, B.; Melocchi, L.; Nizzoli, G.; Troisi, J.; et al. Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat. Microbiol. 2020, 5, 511–524. [Google Scholar] [CrossRef]
- Ma, Y.; Jiang, S.; Zeng, M. In vitro simulated digestion and fermentation characteristics of polysaccharide from oyster (Crassostrea gigas), and its effects on the gut microbiota. Food Res. Int. 2021, 149, 110646. [Google Scholar] [CrossRef] [PubMed]
- Yachida, S.; Mizutani, S.; Shiroma, H.; Shiba, S.; Nakajima, T.; Sakamoto, T.; Watanabe, H.; Masuda, K.; Nishimoto, Y.; Kubo, M.; et al. Metagenomic and metabolomic analyses reveal distinct stage-specific phenotypes of the gut microbiota in colorectal cancer. Nat. Med. 2019, 25, 968–976. [Google Scholar] [CrossRef]
- Leylabadlo, H.E.; Ghotaslou, R.; Feizabadi, M.M.; Farajnia, S.; Moaddab, S.Y.; Ganbarov, K.; Khodadadi, E.; Tanomand, A.; Sheykhsaran, E.; Yousefi, B.; et al. The critical role of Faecalibacterium prausnitzii in human health: An overview. Microb. Pathog. 2020, 149, 104344. [Google Scholar] [CrossRef] [PubMed]
- Rui, Y.; Wan, P.; Chen, G.; Xie, M.; Sun, Y.; Zeng, X.; Liu, Z. Simulated digestion and fermentation in vitro by human gut microbiota of intra- and extra-cellular polysaccharides from Aspergillus cristatus. LWT 2019, 116, 108508. [Google Scholar] [CrossRef]
- Chen, D.; Ding, Y.; Chen, G.; Sun, Y.; Zeng, X.; Ye, H. Components identification and nutritional value exploration of tea (Camellia sinensis L.) flower extract: Evidence for functional food. Food Res. Int. 2020, 132, 109100. [Google Scholar] [CrossRef]
- Xia, Y.; Chen, Y.; Wang, G.; Yang, Y.; Song, X.; Xiong, Z.; Zhang, H.; Lai, P.; Wang, S.; Ai, L. Lactobacillus plantarum AR113 alleviates DSS-induced colitis by regulating the TLR4/MyD88/NF-κB pathway and gut microbiota composition. J. Funct. Foods 2020, 67, 103854. [Google Scholar] [CrossRef]
- Ren, Q.; Yang, B.; Zhang, H.; Ross, R.; Stanton, C.; Chen, H.; Chen, W. c9, t11, c15-CLNA and t9, t11, c15-CLNA from Lactobacillus plantarum ZS2058 Ameliorate Dextran Sodium Sulfate-Induced Colitis in Mice. J. Agric. Food Chem. 2020, 68, 3758–3769. [Google Scholar] [CrossRef]
- Mu, Z.; Yang, Y.; Xia, Y.; Wang, F.; Sun, Y.; Yang, Y.; Ai, L. Probiotic yeast BR14 ameliorates DSS-induced colitis by restoring the gut barrier and adjusting the intestinal microbiota. Food Funct. 2021, 12, 8386–8398. [Google Scholar] [CrossRef]
- Arivarasu, N.; Priyamvada, S.; Mahmood, R. Oral administration of caffeic acid ameliorates the effect of cisplatin on brush border membrane enzymes and antioxidant system in rat intestine. Exp. Toxicol. Pathol. 2013, 65, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Simpson, H.L.; Campbell, B.J. Review article: Dietary fibre-microbiota interactions. Aliment. Pharmacol. Ther. 2015, 42, 158–179. [Google Scholar] [CrossRef] [Green Version]
- Schwiertz, A.; Lehmann, U.; Jacobasch, G.; Blaut, M. Influence of resistant starch on the SCFA production and cell counts of butyrate-producing Eubacterium spp. in the human intestine. J. Appl. Microbiol. 2002, 93, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Tan, M.; Chen, X. Punicic acid ameliorates obesity and liver steatosis by regulating gut microbiota composition in mice. Food Funct. 2021, 12, 7897–7908. [Google Scholar] [CrossRef]
- Zhao, Q.; Hou, D.; Fu, Y.; Xue, Y.; Guan, X.; Shen, Q. Adzuki Bean Alleviates Obesity and Insulin Resistance Induced by a High-Fat Diet and Modulates Gut Microbiota in Mice. Nutrient 2021, 13, 3240. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ADG (g/day) |
|---|---|
| Control | 0.855 ± 0.036 |
| Asparagus | 0.816 ± 0.039 |
| Broccoli | 0.788 ± 0.025 |
| Cabbage | 0.737 ± 0.043 |
| Group | Control | Asparagus | Broccoli | Cabbage |
|---|---|---|---|---|
| IgA, ng/mL | 80.12 ± 0.82 c | 88.05 ± 1.21 b | 93.21± 1.05 a | 84.86 ± 0.93 b |
| IgG, ng/mL | 813.45 ± 9.83 | 838.03 ± 12.02 | 823.11 ± 9.84 | 827.20 ± 11.82 |
| IgM, ng/mL | 8.56 ± 0.15 b | 9.34 ± 0.14 a | 9.26 ± 0.08 a | 8.87 ± 0.13 ab |
| Group | Control | Asparagus | Broccoli | Cabbage |
|---|---|---|---|---|
| Jejunum, pg/mL | ||||
| IL-1 | 69.02 ± 1.17 b | 80.27 ± 1.47 a | 61.07 ± 1.61 c | 68.84 ± 1.23 b |
| IL-10 | 1676.67 ± 36.71 a | 1326.68 ± 17.17 c | 1494.44 ± 23.72 b | 1589.64 ± 23.35 ab |
| TNF-α | 728.13 ± 14.92 b | 950.23 ± 18.45 a | 729.37 ± 17.83 b | 918.64 ± 8.94 a |
| IFN-γ | 762.96 ± 23.15 c | 889.94 ± 11.79 b | 1131.92 ± 18.49 a | 1071.57 ± 10.54 a |
| Ileum, pg/mL | ||||
| IL-1 | 65.39 ± 2.71 b | 70.62 ± 1.47 b | 68.37 ± 1.69 b | 100.39 ± 1.41 a |
| IL-10 | 1629.44 ± 44.49 a | 1338.77 ± 19.50 b | 1546.33 ± 20.21 a | 1228.75 ± 24.14 b |
| TNF-α | 713.17 ± 13.56 c | 762.04 ± 14.36 c | 928.09 ± 16.00 a | 829.00 ± 13.79 b |
| IFN-γ | 735.64 ± 6.64 b | 1066.44 ± 19.37 a | 792.01 ± 19.42 b | 765.54 ± 16.22 b |
| Group | Control | Asparagus | Broccoli | Cabbage |
|---|---|---|---|---|
| Caecum, μmol/g | ||||
| Acetic acid | 6.03 ± 0.36 ab | 4.67 ± 0.23 b | 4.87 ± 0.32 b | 6.40 ± 0.47 a |
| Propionic acid | 1.70 ± 0.20 b | 2.00 ± 0.14 b | 2.02 ± 0.18 b | 3.59 ± 0.33 a |
| Butyric acid | 1.48 ± 0.22 b | 1.95 ± 0.18 ab | 2.16 ± 0.38 ab | 2.82 ± 0.38 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, Y.; Yu, H.; Zhang, L.; Ruan, Z. Dietary Vegetable Powders Modulate Immune Homeostasis and Intestinal Microbiota in Mice. Foods 2022, 11, 27. https://doi.org/10.3390/foods11010027
Zou Y, Yu H, Zhang L, Ruan Z. Dietary Vegetable Powders Modulate Immune Homeostasis and Intestinal Microbiota in Mice. Foods. 2022; 11(1):27. https://doi.org/10.3390/foods11010027
Chicago/Turabian StyleZou, Yixin, Haifei Yu, Li Zhang, and Zheng Ruan. 2022. "Dietary Vegetable Powders Modulate Immune Homeostasis and Intestinal Microbiota in Mice" Foods 11, no. 1: 27. https://doi.org/10.3390/foods11010027
APA StyleZou, Y., Yu, H., Zhang, L., & Ruan, Z. (2022). Dietary Vegetable Powders Modulate Immune Homeostasis and Intestinal Microbiota in Mice. Foods, 11(1), 27. https://doi.org/10.3390/foods11010027