
Effects of Different Ionic Polysaccharides in Cooked Lean Pork Batters on Intestinal Health in Mice


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals and Diets
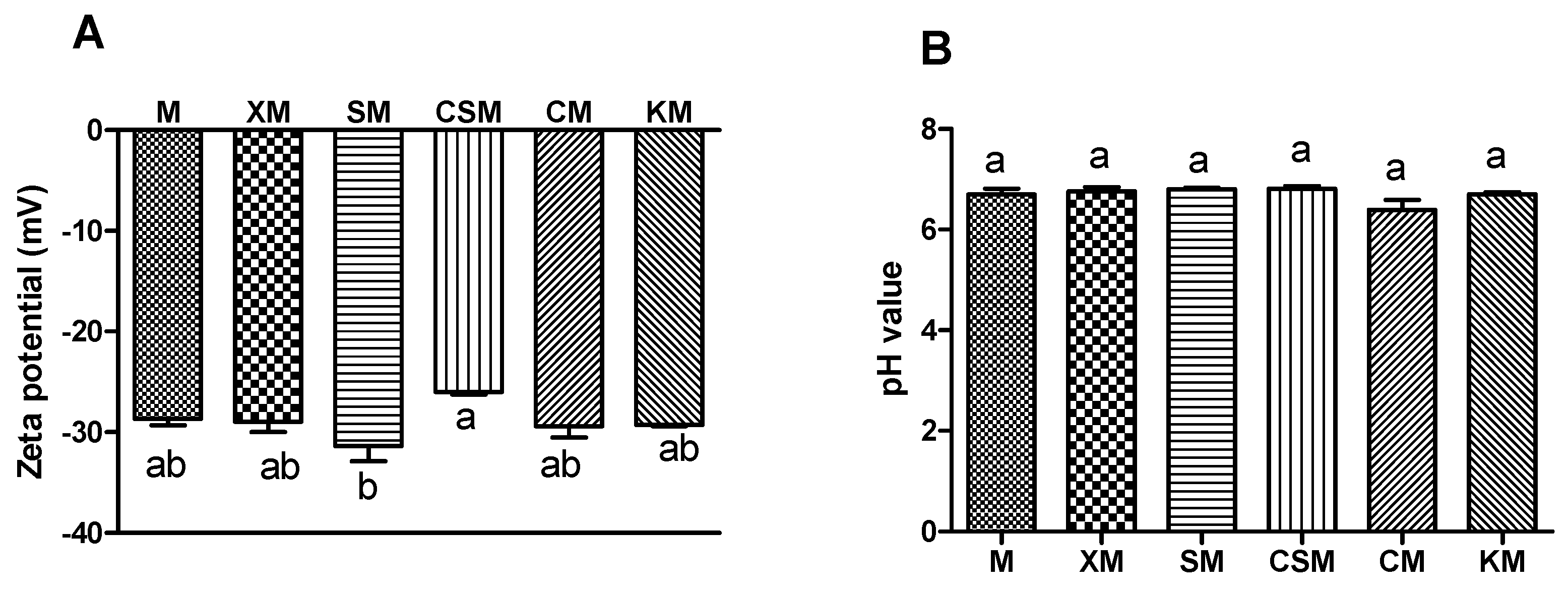
2.3. Zeta Potential and pH Value of the Small Intestinal Digesta
2.4. Free Amino Acid Profile of Small Intestinal Digesta
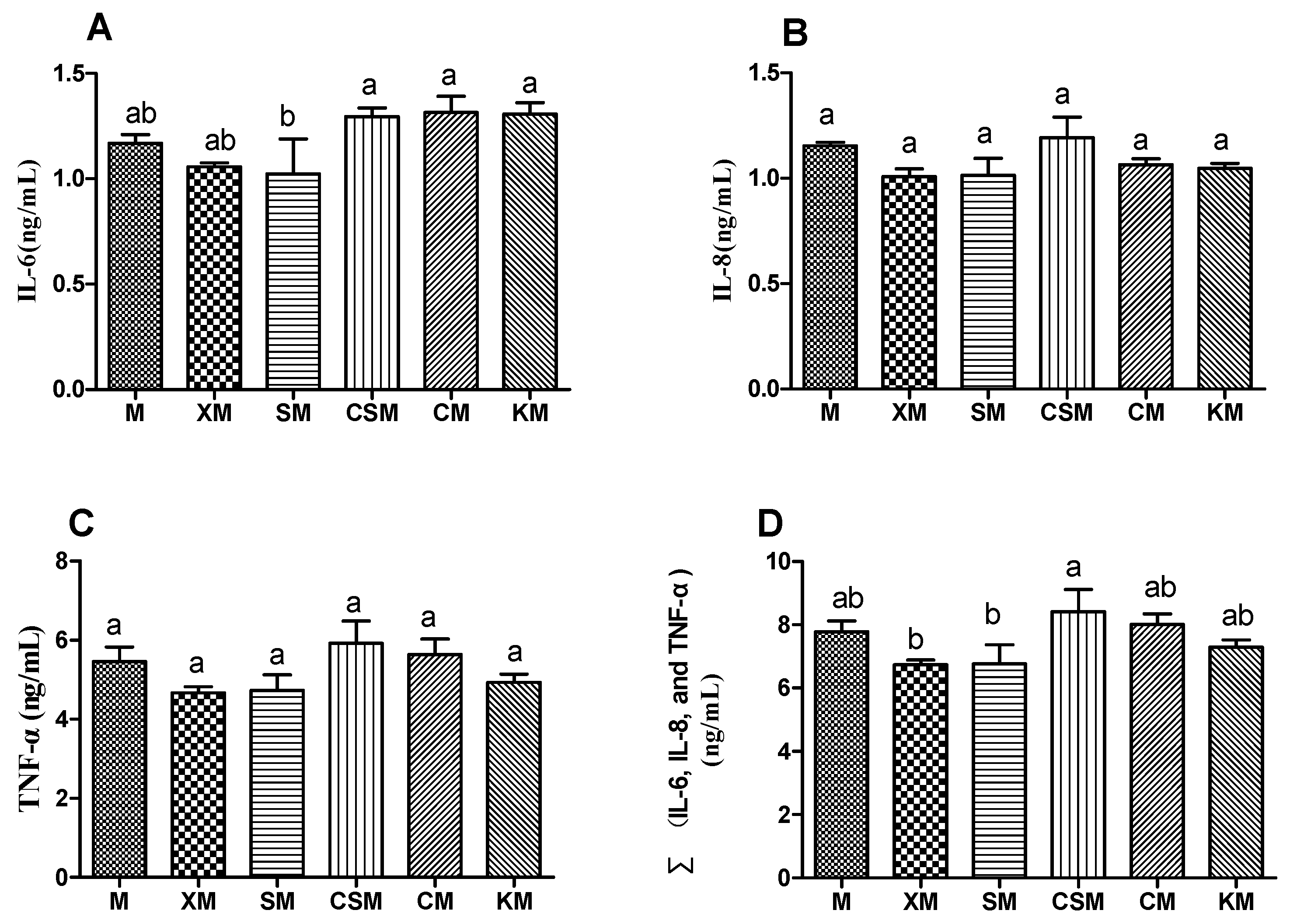
2.5. Biochemical and Morphologic Analysis of the Colon
2.6. Deoxyribonucleic Acid Isolation, Polymerase Chain Reaction Amplification and Illumina MiSeq Sequencing
2.7. Short-Chain Fatty Acids (SCFAs) of the Faeces
2.8. Statistical Analysis
3. Results and Discussion
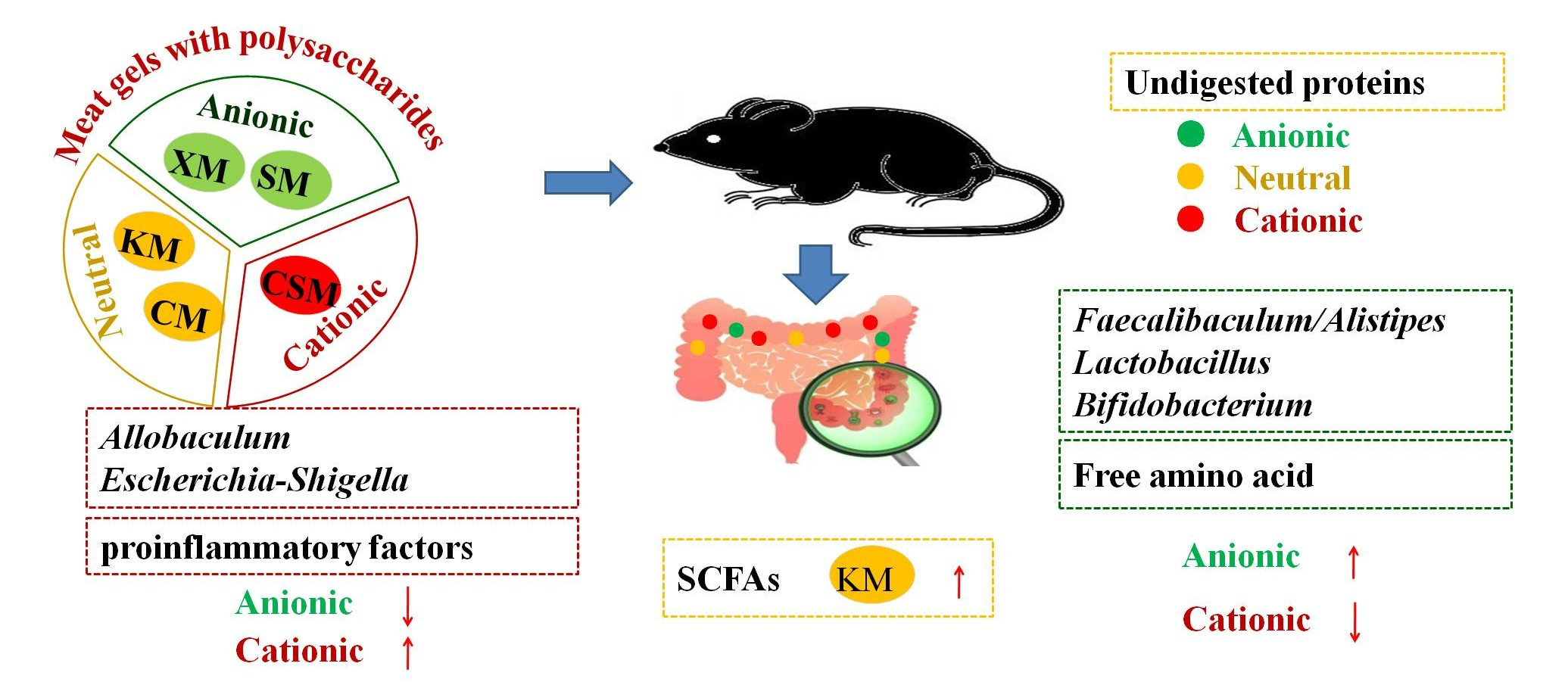
3.1. Zeta Potential
3.2. Free Amino Acids
3.3. Colon Cytokines and Morphology
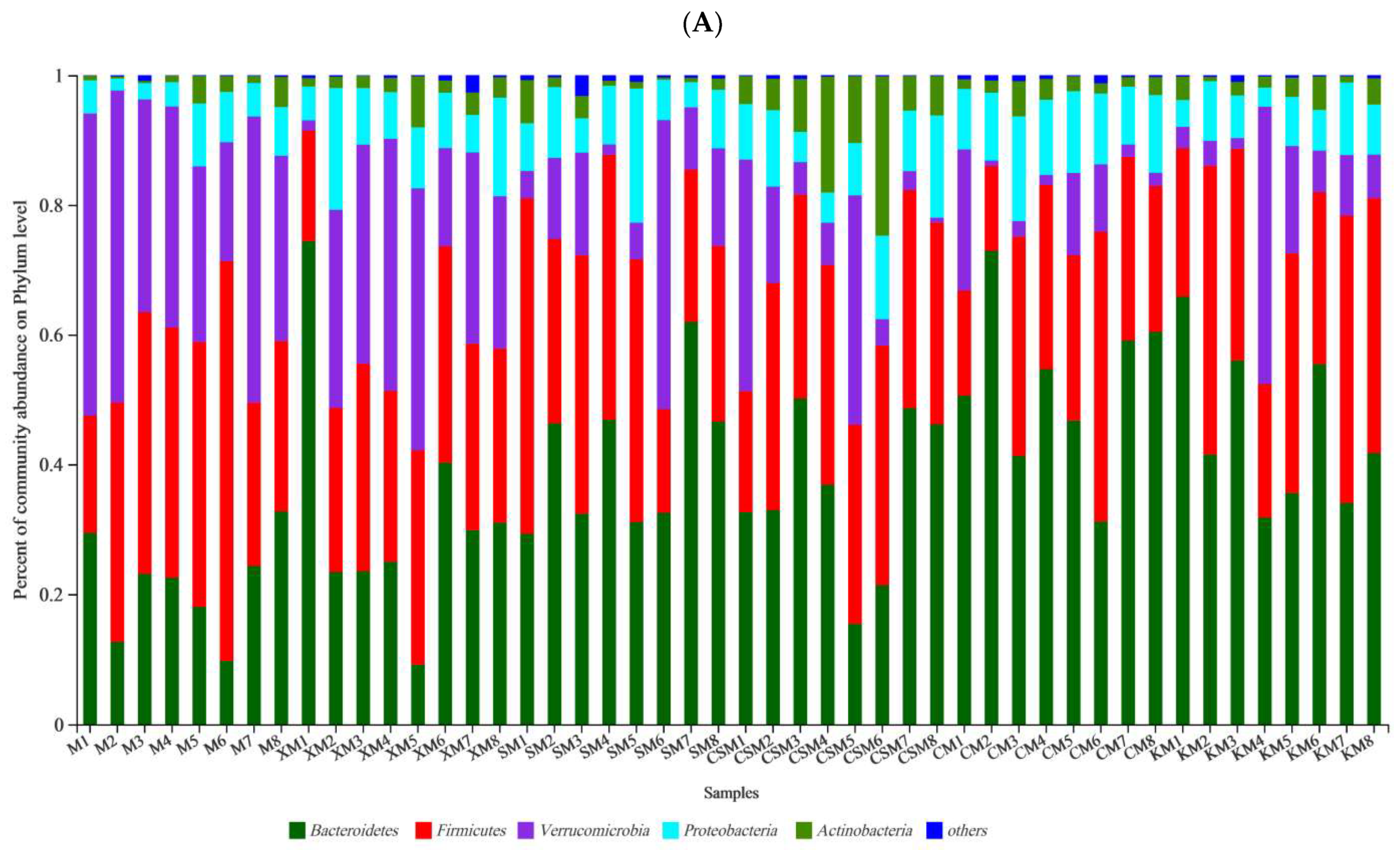
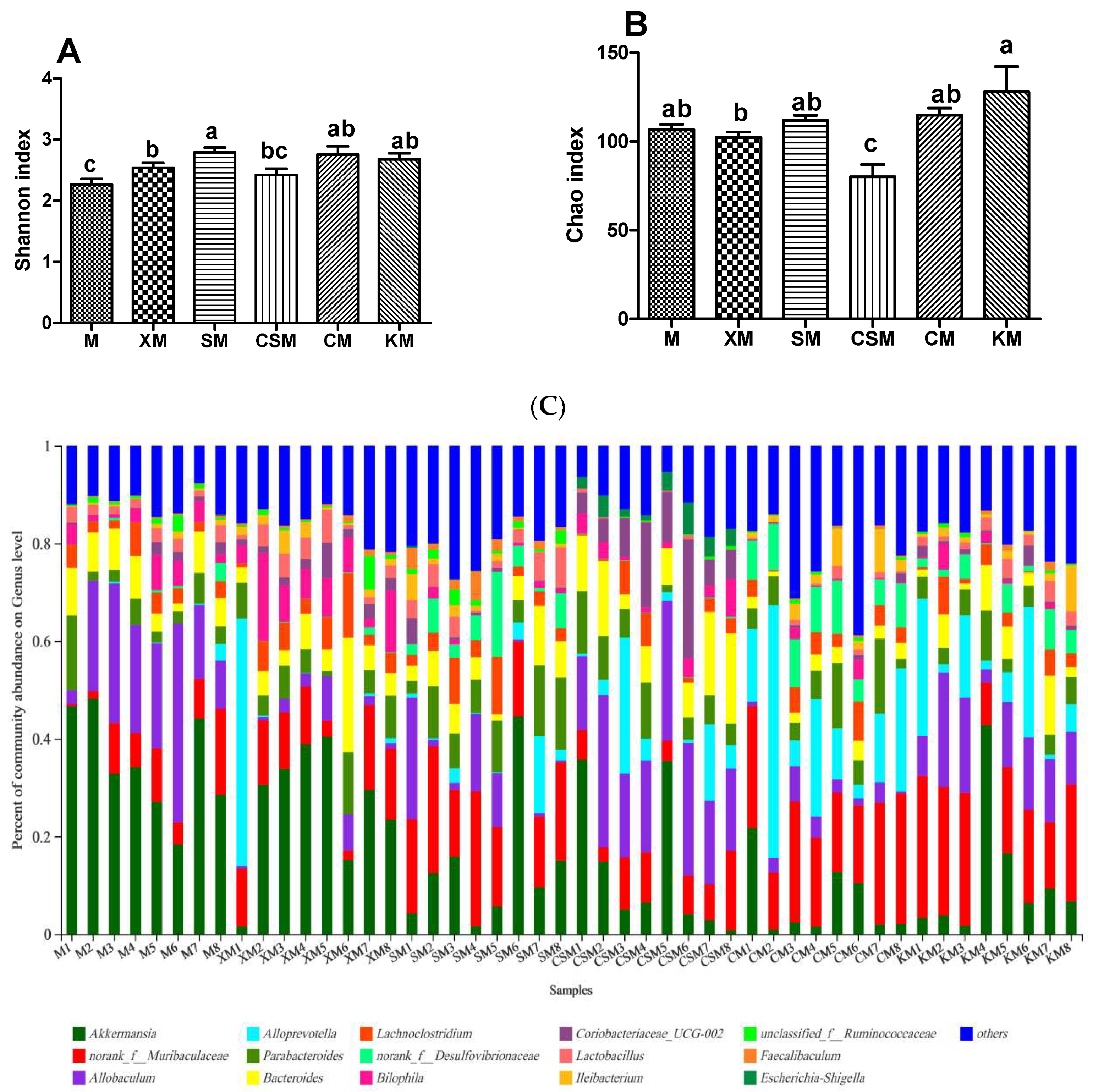
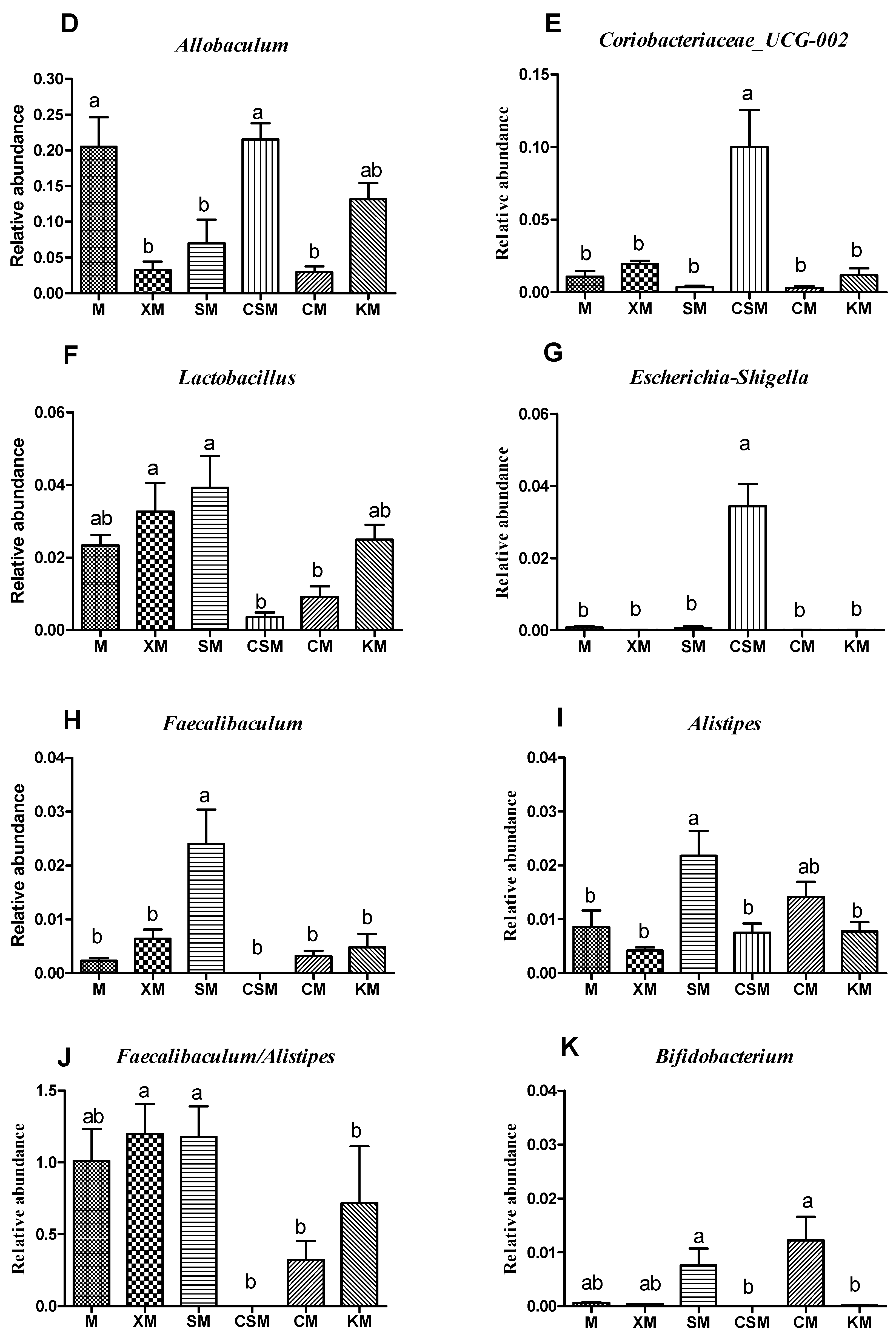
3.4. Microbial Diversity and Composition of the Gut Microbiota
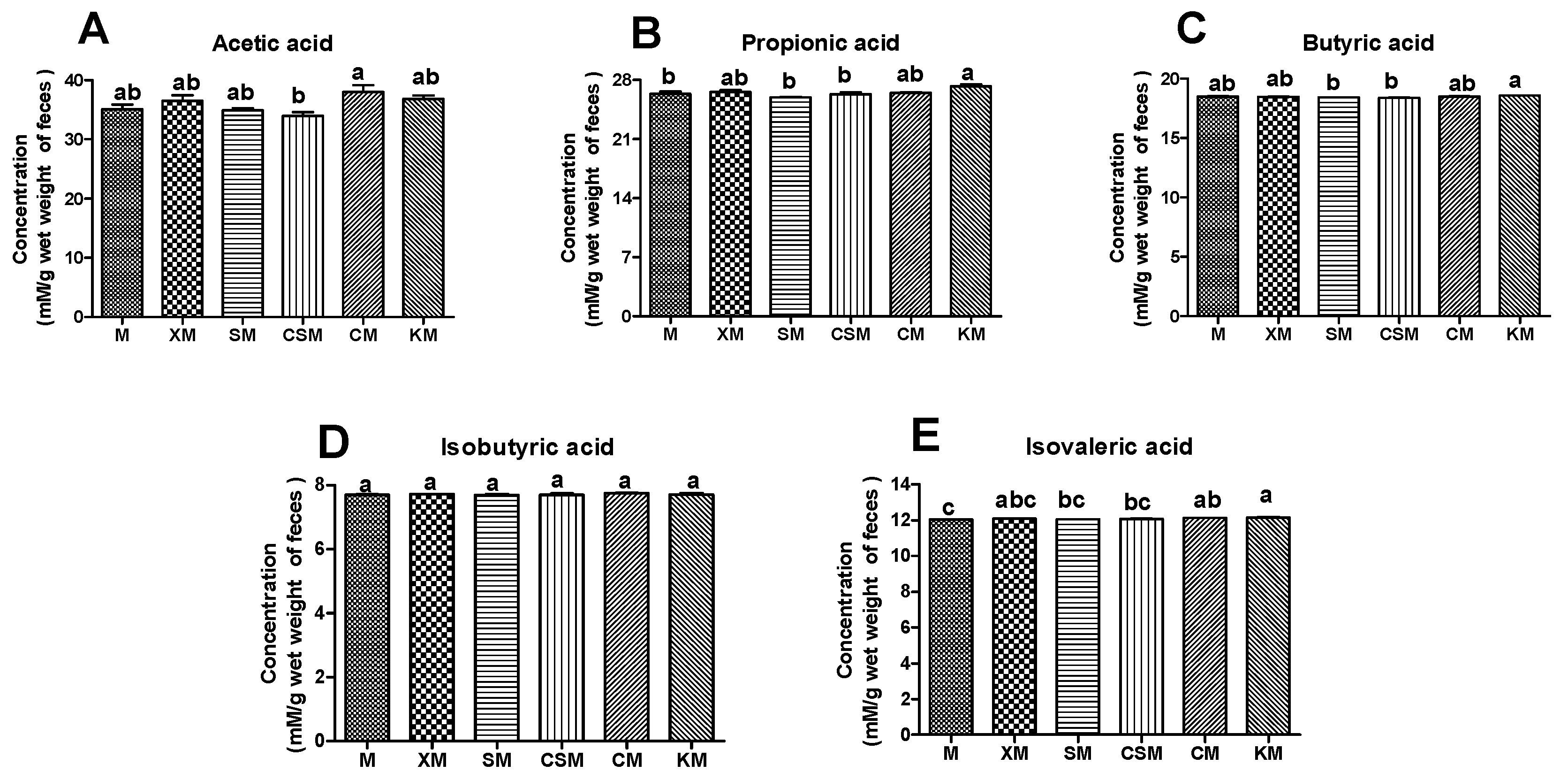
3.5. SCFAs of Mice Faeces
3.6. Growth Performance in Mice
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Toldrá, F.; Rubio, M.A.; Navarro, J.L.; Cabrerizo, L. Quality aspects of pork meat and its nutritional impact. Qual. Fresh Process. Foods 2004, 542, 25–31. [Google Scholar] [CrossRef]
- Zhang, M.; Zou, X.; Zhao, D.; Zhao, F.; Li, C. Front cover: Pork meat proteins alter gut microbiota and lipid metabolism genes in the colon of adaptive immune-deficient mice. Mol. Nutr. Food Res. 2020, 64, 2070021. [Google Scholar] [CrossRef]
- Zeng, W.; He, D.; Xing, Y.; Liu, J.; Xing, X. Internal connections between dietary intake and gut microbiota homeostasis in disease progression of ulcerative colitis: A review. Food Sci. Hum. Well 2021, 10, 119–130. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10 −/− mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zock, P.L.; Blom, W.A.M.; Nettleton, J.A.; Hornstra, G. Progressing insights into the role of dietary fats in the prevention of cardiovascular disease. Curr. Cardiol Rep. 2016, 18, 111. [Google Scholar] [CrossRef] [Green Version]
- Amaral, D.S.; Cardelle-Cobas, A.; do Nascimento, B.M.S.; Monteiro, M.J.; Madruga, M.S.; Pintado, M.M.E. Development of a low fat fresh pork sausage based on chitosan with health claims: Impact on the quality, functionality and shelf-life. Food Funct. 2015, 6, 2768–2778. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, C.; Chen, X.; Li, P.; Ma, F.; Lu, Q. Contribution of three ionic types of polysaccharides to the thermal gelling properties of chicken breast myosin. J. Agric. Food Chem. 2014, 62, 2655–2662. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Gut microbiota fermentation of marine polysaccharides and its effects on intestinal ecology: An overview. Carbohydr. Polym. 2018, 179, 173–185. [Google Scholar] [CrossRef]
- Hu, J.L.; Nie, S.P.; Min, F.F.; Xie, M.Y. Polysaccharide from seeds of plantago asiatical. increases short-chain fatty acid production and fecal moisture along with lowering pH in mouse colon. J. Agric. Food Chem. 2012, 60, 11525–11532. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, Z.; Zhu, C.; Mou, H.; Kong, Q. Nondigestible carbohydrates, butyrate, and butyrate-producing bacteria. Crit. Rev. Food Sci. 2019, 59, S130–S152. [Google Scholar] [CrossRef]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thøgersen, R.; Castro-Mejía, J.L.; Sundekilde, U.K.; Hansen, L.H.; Hansen, A.K.; Nielsen, D.S.; Bertram, H.C. Ingestion of an inulin-enriched pork sausage product positively modulates the gut microbiome and metabolome of healthy rats. Mol. Nutr. Food Res. 2018, 62, 1800608. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Pan, Q.; Ma, F.; Li, P.J.; Xu, B.C.; Chen, C.G. Amelioration of growth performance, lipid accumulation and intestinal health in mice by a cooked mixture of lean meat and resistant starch. Mol. Nutr. Food Res. 2019, 63, 1801364. [Google Scholar] [CrossRef]
- Lu, W.; Nishinari, K.; Matsukawa, S.; Fang, Y. The future trends of food hydrocolloids. Food Hydrocolloid. 2020, 103, 105713. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, W.; Jin, W.; Shah, B.R.; Li, Y.; Li, B. Influence of anionic alginate and cationic chitosan on physicochemical stability and carotenoids bioaccessibility of soy protein isolate-stabilized emulsions. Food Res. Int. 2015, 77, 419–425. [Google Scholar] [CrossRef]
- Qin, D.; Yang, X.; Gao, S.; Yao, J.; McClements, D.J. Influence of hydrocolloids (dietary fibers) on lipid digestion of protein-stabilized emulsions: Comparison of neutral, anionic, and cationic polysaccharides. J. Food Sci. 2016, 81, C1636–C1645. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Wang, X.X.; Yu, X.; Wang, Y.; Li, P.J.; Xu, B.C.; Cai, K.Z.; Chen, C.G. Gastrointestinal digestion and cecal fermentation of a mixed gel of lean pork meat and resistant starch in mice. Food Funct. 2020, 11, 6834–6842. [Google Scholar] [CrossRef]
- Liu, J.; Chai, J.; Yuan, Y.; Zhang, T.; Saini, R.K.; Yang, M.; Shang, X. Dextran sulfate facilitates egg white protein to form transparent hydrogel at neutral pH: Structural, functional, and degradation properties. Food Hydrocolloid. 2022, 122, 107094. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhang, R.; Zou, L.; McClements, D.J. Protein encapsulation in alginate hydrogel beads: Effect of pH on microgel stability, protein retention and protein release. Food Hydrocolloid. 2016, 58, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.M.; Finlay, B.B. Gut Microbiota in Health and Disease. Physiol Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jahan-Mihan, J.A.; Luhovyy, B.L.; El Khoury, D.; Anderson, G.H. Dietary proteins as determinants of metabolic and physiologic functions of the gastrointestinal tract. Nutrients 2011, 3, 574–603. [Google Scholar] [CrossRef] [Green Version]
- Wu, G. Functional amino acids in growth, reproduction, and health. Adv. Nutr. 2010, 1, 31–37. [Google Scholar] [CrossRef]
- Marc Rhoads, J.; Wu, G. Glutamine, arginine, and leucine signaling in the intestine. Amino. Acids 2009, 37, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Schaart, M.W.; Schierbeek, H.; van der Schoor, S.R.D.; Stoll, B.; Burrin, D.; Reeds, P.J.; Goudoever, J.B.V. Threonine utilization is high in the intestine of piglets. J. Nutr. 2005, 135, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.; Wu, Y.; Wang, W.; Wu, Y.; Zhang, Y.; Wang, S. Structural modification and digestibility change of β-lactoglobulin modified by methylglyoxal with the simulated reheating of dairy products. Food Chem. 2019, 288, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2011, 56, 184–196. [Google Scholar] [CrossRef]
- Lindström, C.; Holst, O.; Hellstrand, P.; Öste, R.; Andersson, K.E. Evaluation of commercial microbial hydrocolloids concerning their effects on plasma lipids and caecal formation of SCFA in mice. Food Hydrocolloid. 2012, 28, 367–372. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Hu, Y.; An, N.; Feng, Z.; Liu, J.; Mou, J.; Hu, T.; Guan, H.; Zhang, D.F.; Mao, Y.J. Alginate oligosaccharide alleviates monocrotaline-induced pulmonary hypertension via anti-oxidant and anti-inflammation pathways in rats. Int. Heart J. 2020, 61, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Aparicio, I.; Mengíbar, M.; Heras, A. Effect of chito-oligosaccharides over human faecal microbiota during fermentation in batch cultures. Carbohydr. Polym. 2016, 137, 617–624. [Google Scholar] [CrossRef]
- Shen, F.; Zheng, R.D.; Sun, X.Q.; Ding, W.J.; Wang, X.Y.; Fan, J.G. Gut microbiota dysbiosis in patients with non-alcoholic fatty liver disease. Hepatob Pancreat Dis. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review article: Insights into colonic protein fermentation, its modulation and potential health implications. Aliment. Pharm Ther. 2015, 43, 181–196. [Google Scholar] [CrossRef] [Green Version]
- Madsen, K.; Cornish, A.; Soper, P.; McKaigney, C.; Jijon, H.; Yachimec, C.; Doyle, J.; Jewell, L.; Simone, C.D. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology 2001, 121, 580–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Yazaki, T.; Takahashi, H.; Kuda, T.; Kimura, B. Diet-induced changes in alginate- and laminaran-fermenting bacterial levels in the caecal contents of rats. J. Funct. Foods. 2013, 5, 389–394. [Google Scholar] [CrossRef]
- Kuda, T.; Yano, T.; Matsuda, N.; Nishizawa, M. Inhibitory effects of laminaran and low molecular alginate against the putrefactive compounds produced by intestinal microflora in vitro and in rats. Food Chem. 2005, 91, 745–749. [Google Scholar] [CrossRef]
- Zagato, E.; Pozzi, C.; Bertocchi, A.; Schioppa, T.; Saccheri, F.; Guglietta, S.; Fosso, B.; Melocchi, L.; Nizzoli, G.; Troisi, J.; et al. Endogenous murine microbiota member Faecalibaculum rodentium and its human homologue protect from intestinal tumour growth. Nat. Microbiol. 2020, 5, 511–524. [Google Scholar] [CrossRef] [PubMed]
- Li, D.P.; Cui, M.; Tan, F.; Liu, X.Y.; Yao, P. High Red Meat Intake Exacerbates Dextran Sulfate-Induced Colitis by Altering Gut Microbiota in Mice. Front. Nutr. 2021, 8, 646819. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut. Microbes. 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, M.; Shang, W.; Ma, Q.; Strappe, P.; Zhou, Z. Abundance of probiotics and butyrate-production microbiome manages constipation via short-chain fatty acids production and hormones secretion. Mol. Nutr. Food Res. 2019, 63, 1801187. [Google Scholar] [CrossRef]
- Vinolo, M.A.R.; Rodrigues, H.G.; Hatanaka, E.; Sato, F.T.; Sampaio, S.C.; Curi, R. Suppressive effect of short-chain fatty acids on production of proinflammatory mediators by neutrophils. J. Nutr. Biochem. 2011, 22, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. AM J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Geypens, B.; Claus, D.; Evenepoel, P.; Hiele, M.; Maes, B.; Peeters, M.; Rutgeerts, P.; Ghoos, Y. Influence of dietary protein supplements on the formation of bacterial metabolites in the colon. Gut 1997, 41, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Henao-Mejia, J.; Elinav, E.; Jin, H.; Mehal, L.; Strowig, W.Z.; Thaiss, T.; Kau, C.A.; Eisenbarth, A.L.; Jurczak, S.C.; Camporez, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elinav, E.; Strowig, T.; Kau, A.L.; Henao-Mejia, J.; Thaiss, C.A.; Booth, C.J.; Peaper, D.R.; Bertin, J.; Eisenbarth, S.C.; Gordon, J.I.; et al. NLRP6 Inflammasome Regulates Colonic Microbial Ecology and Risk for Colitis. Cell 2011, 145, 745–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Diets (g kg−1) | |||||
|---|---|---|---|---|---|---|
| M | XM | SM | CSM | CM | KM | |
| Meat powder (Corresponding minced meat) | 200 (800) | 200 (800) | 200 (800) | 200 (800) | 200 (800) | 200 (800) |
| Cellulose | 50 | 45.2 | 45.2 | 45.2 | 45.2 | 45.2 |
| Xanthan gum | 0 | 4.8 | 0 | 0 | 0 | 0 |
| Sodium alginate | 0 | 0 | 4.8 | 0 | 0 | 0 |
| Chitosan | 0 | 0 | 0 | 4.8 | 0 | 0 |
| Curdlan | 0 | 0 | 0 | 0 | 4.8 | 0 |
| Konjac gum | 0 | 0 | 0 | 0 | 0 | 4.8 |
| Sodium chloride | 8 | 8 | 8 | 8 | 8 | 8 |
| Corn starch | 397 | 397 | 397 | 397 | 397 | 397 |
| Dextrinized corn starch | 132 | 132 | 132 | 132 | 132 | 132 |
| Sucrose | 100 | 100 | 100 | 100 | 100 | 100 |
| Soybean oil | 70 | 70 | 70 | 70 | 70 | 70 |
| l-Cystine | 3 | 3 | 3 | 3 | 3 | 3 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| Mineral mix (AIN-93G-MX) | 32.4 | 32.4 | 32.4 | 32.4 | 32.4 | 32.4 |
| Vitamin mix (AIN-93G-VX) | 10 | 10 | 10 | 10 | 10 | 10 |
| Tert-butylhydroquinone | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 |
| Total | 1004.914 | 1004.914 | 1004.914 | 1004.914 | 1004.914 | 1004.914 |
| Amino Acids | M | XM | SM | CSM | CM | KM |
|---|---|---|---|---|---|---|
| Asparagine | 97.39 ± 22.92 ab | 112.39 ± 8.40 a | 0.00 | 0.00 | 0.00 | 0.00 |
| Threonine | 115.08 ± 26.15 a | 135.83 ± 7.28 a | 74.63 ± 2.71 b | 52.69 ± 0.68 b | 64.17 ± 9.67 b | 64.63 ± 9.17 b |
| Serine | 112.61 ± 21.40 a | 124.45 ± 5.00 a | 134.32 ± 7.38 a | 99.09 ± 1.45 a | 122.51 ± 16.78 a | 124.72 ± 13.75 a |
| Glutamine | 214.06 ± 49.49 a | 255.79 ± 15.25 a | 264.53 ± 12.42 a | 225.19 ± 9.52 a | 230.21 ± 26.05 a | 232.84 ± 17.94 a |
| Glycine | 64.65 ± 6.58 ab | 70.44 ± 0.81 ab | 76.07 ± 5.28 a | 55.53 ± 1.06 b | 70.90 ± 8.73 ab | 72.32 ± 11.27 ab |
| Alanine | 155.70 ± 28.58 ab | 179.92 ± 5.46 a | 192.47 ± 10.27 a | 134.31 ± 2.15 b | 172.01 ± 21.38 ab | 173.33 ± 11.76 ab |
| Cysteine | 21.83 ± 1.76 c | 27.25 ± 0.98 ab | 29.62 ± 0.65 a | 22.02 ± 0.09 c | 24.24 ± 1.99 bc | 25.78 ± 1.24 b |
| Valine | 126.04 ± 32.22 a | 151.39 ± 7.14 a | 160.68 ± 10.14 a | 125.15 ± 3.11 a | 140.66 ± 19.34 a | 144.20 ± 3.77 a |
| Methionine | 78.45 ± 15.93 bc | 101.69 ± 3.64 a | 96.87 ± 4.64 ab | 71.57 ± 2.21 c | 90.19 ± 11.26 abc | 95.13 ± 1.04 ab |
| Isoleucine | 120.74 ± 33.14 a | 141.16 ± 7.97 a | 150.40 ± 9.39 a | 119.04 ± 1.92 a | 131.09 ± 18.55 a | 136.35 ± 5.50 a |
| Leucine | 262.10 ± 62.02 ab | 323.13 ± 11.81 ab | 335.13 ± 18.63 a | 241.61 ± 8.42 b | 300.82 ± 41.84 ab | 306.55 ± 6.35 ab |
| Tyrosine | 172.74 ± 32.25 bc | 219.99 ± 7.18 a | 222.29 ± 4.95 a | 153.67 ± 9.36 c | 215.20 ± 20.93 ab | 217.78 ± 11.37 ab |
| Phenylalanine | 199.42 ± 37.78 bc | 248.31 ± 5.69 ab | 259.00 ± 17.80 a | 177.77 ± 13.61 c | 236.32 ± 23.88 ab | 239.20 ± 10.37 ab |
| Lysine | 210.09 ± 30.41 ab | 244.48 ± 2.57 a | 258.40 ± 11.00 a | 181.52 ± 3.77 b | 242.37 ± 28.60 a | 244.02 ± 14.57 a |
| Histidine | 50.63 ± 6.57 ab | 58.46 ± 1.16 a | 62.02 ± 3.19 a | 45.12 ± 0.54 b | 58.18 ± 5.92 a | 58.16 ± 5.82 a |
| Arginine | 241.04 ± 43.05 bc | 285.51 ± 5.87 ab | 305.92 ± 10.81 a | 187.32 ± 2.30 c | 286.51 ± 34.49 ab | 292.61 ± 9.73 ab |
| Proline | 66.08 ± 12.39 a | 74.50 ± 5.18 a | 76.14 ± 4.72 a | 58.19 ± 2.17 a | 65.70 ± 11.36 a | 67.12 ± 4.99 a |
| Total amino acid | 2308.64 ± 454.91 ab | 2754.68 ± 86.63a | 2733.72 ± 74.80a | 1949.78 ± 49.29 b | 2451.09 ± 299.30 ab | 2531.04 ± 103.13 ab |
| Diet Groups | Body Weight Gain (g) | Food Intake (g/week) | Liver Weight (g) | Small Intestine Weight (g) | Colon Weight (g) |
|---|---|---|---|---|---|
| M | 2.97 ± 2.09 b | 18.88 ± 2.56 a | 0.96 ± 0.12 a | 0.84 ± 0.21 a | 0.23 ± 0.14 a |
| XM | 5.05 ± 2.04 a | 22.03 ± 1.33 a | 1.02 ± 0.15 a | 0.82 ± 0.13 a | 0.26 ± 0.14 a |
| SM | 3.34 ± 1.00 ab | 20.61 ± 1.49 a | 1.00 ± 0.10 a | 0.75 ± 0.16 a | 0.17 ± 0.10 a |
| CSM | 2.70 ± 1.61 b | 22.02 ± 1.57 a | 0.97 ± 0.10 a | 0.77 ± 0.27 a | 0.19 ± 0.04 a |
| CM | 3.52 ± 1.26 ab | 22.41 ± 1.99 a | 1.02 ± 0.11 a | 0.90 ± 0.09 a | 0.16 ± 0.03 a |
| KM | 3.36 ± 1.59 ab | 20.58 ± 2.33 a | 1.03 ± 0.14 a | 0.89 ± 0.12 a | 0.18 ± 0.04 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Zou, L.-F.; Xiong, J.-H.; Pan, J.-Z.; Li, P.-J.; Chen, C.-G. Effects of Different Ionic Polysaccharides in Cooked Lean Pork Batters on Intestinal Health in Mice. Foods 2022, 11, 1372. https://doi.org/10.3390/foods11101372
Yu X, Zou L-F, Xiong J-H, Pan J-Z, Li P-J, Chen C-G. Effects of Different Ionic Polysaccharides in Cooked Lean Pork Batters on Intestinal Health in Mice. Foods. 2022; 11(10):1372. https://doi.org/10.3390/foods11101372
Chicago/Turabian StyleYu, Xia, Li-Fang Zou, Jia-Hao Xiong, Jing-Zhi Pan, Pei-Jun Li, and Cong-Gui Chen. 2022. "Effects of Different Ionic Polysaccharides in Cooked Lean Pork Batters on Intestinal Health in Mice" Foods 11, no. 10: 1372. https://doi.org/10.3390/foods11101372
APA StyleYu, X., Zou, L. -F., Xiong, J. -H., Pan, J. -Z., Li, P. -J., & Chen, C. -G. (2022). Effects of Different Ionic Polysaccharides in Cooked Lean Pork Batters on Intestinal Health in Mice. Foods, 11(10), 1372. https://doi.org/10.3390/foods11101372