
Role of Yeasts on the Sensory Component of Wines


 ,
,  , and
, and
Abstract
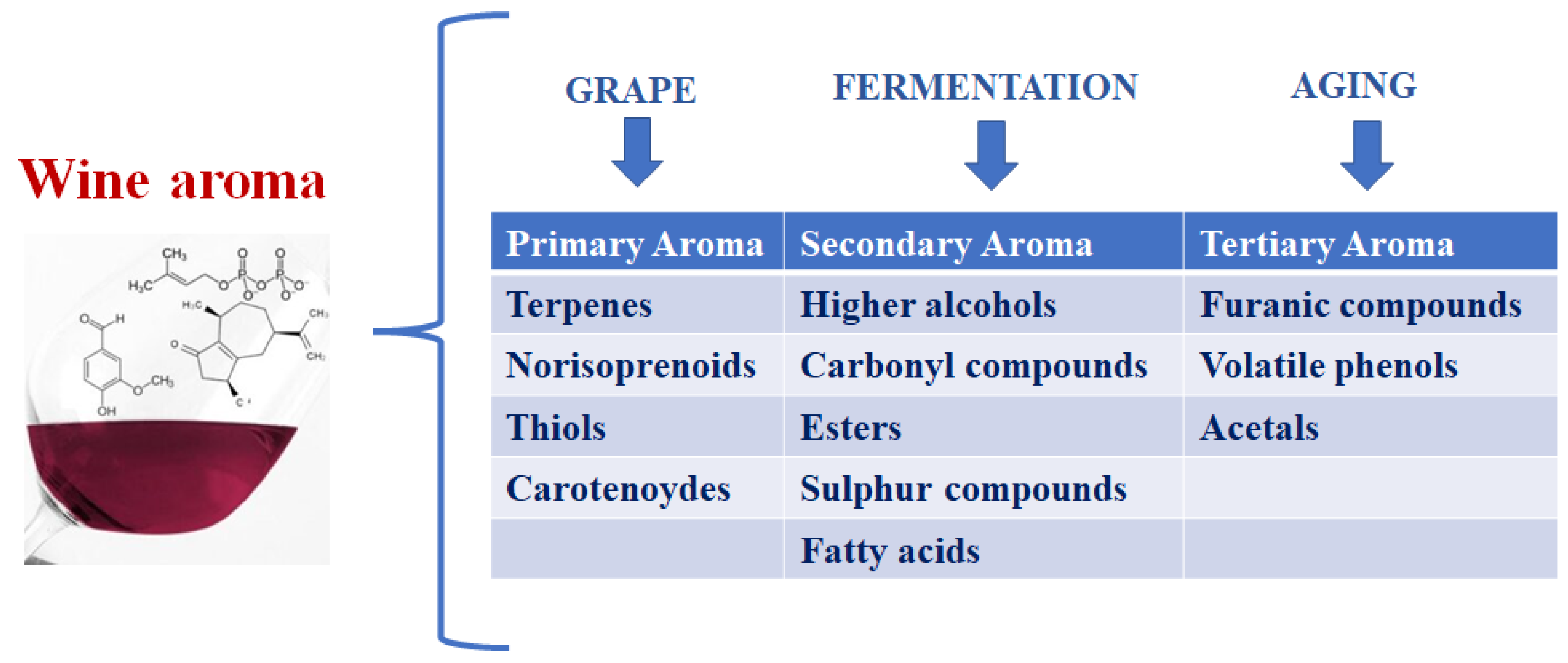
:1. Introduction
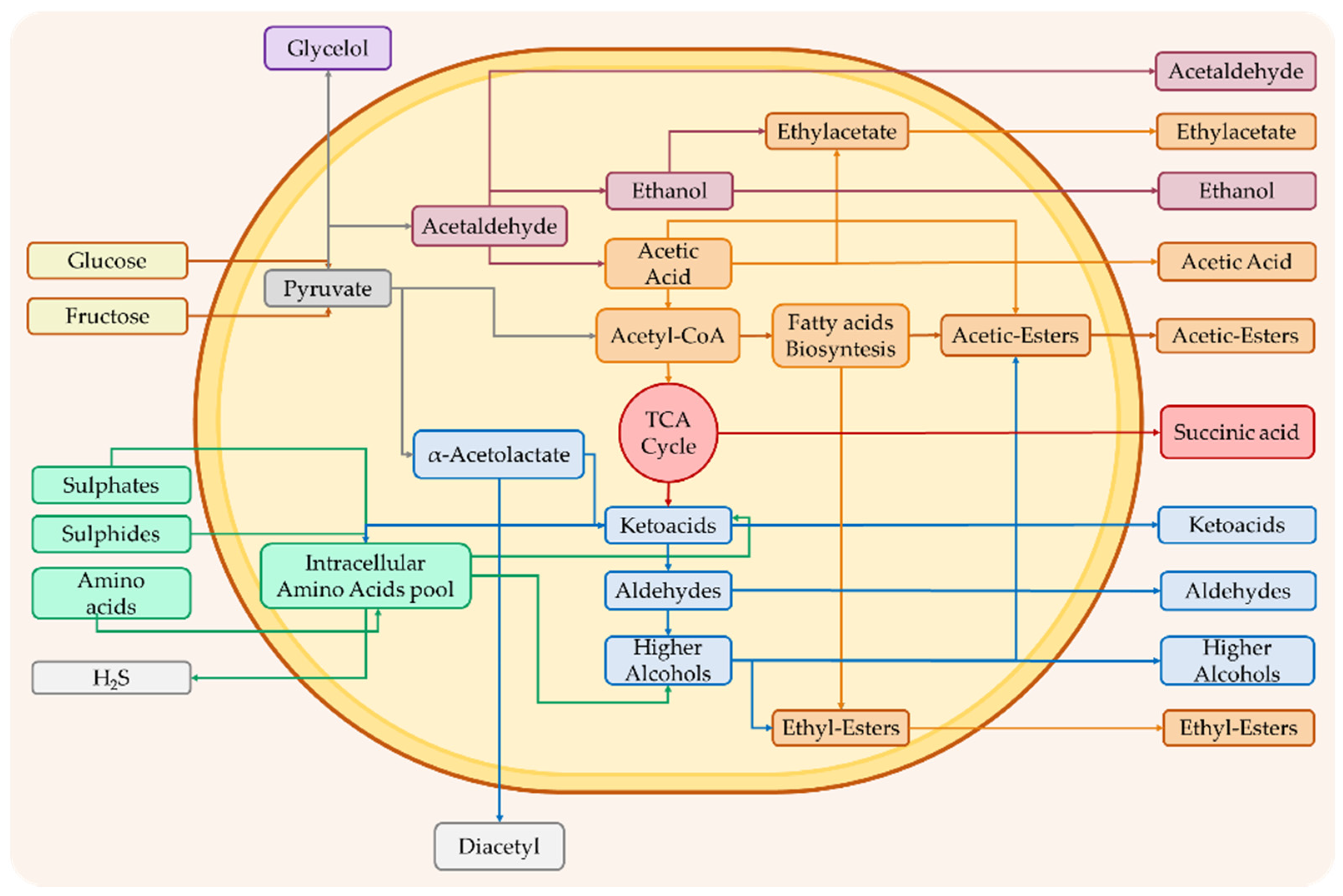
1.1. Compounds from Primary Metabolism
1.2. Compounds from Secondary Metabolism
1.2.1. Esters and Alcohols
1.2.2. Aldehydes
2. Production of Aromatic Compounds in Relation to the Main Compositional and Process Variables
2.1. Effect of the Carbon Source on Fermentation Processes and Aroma Compounds
2.2. Effect of the Fermentation Temperature on Fermentation Processes and Aroma Compounds
2.3. Effect of Non-Thermal Technologies on the Production of Wine Aroma Compounds
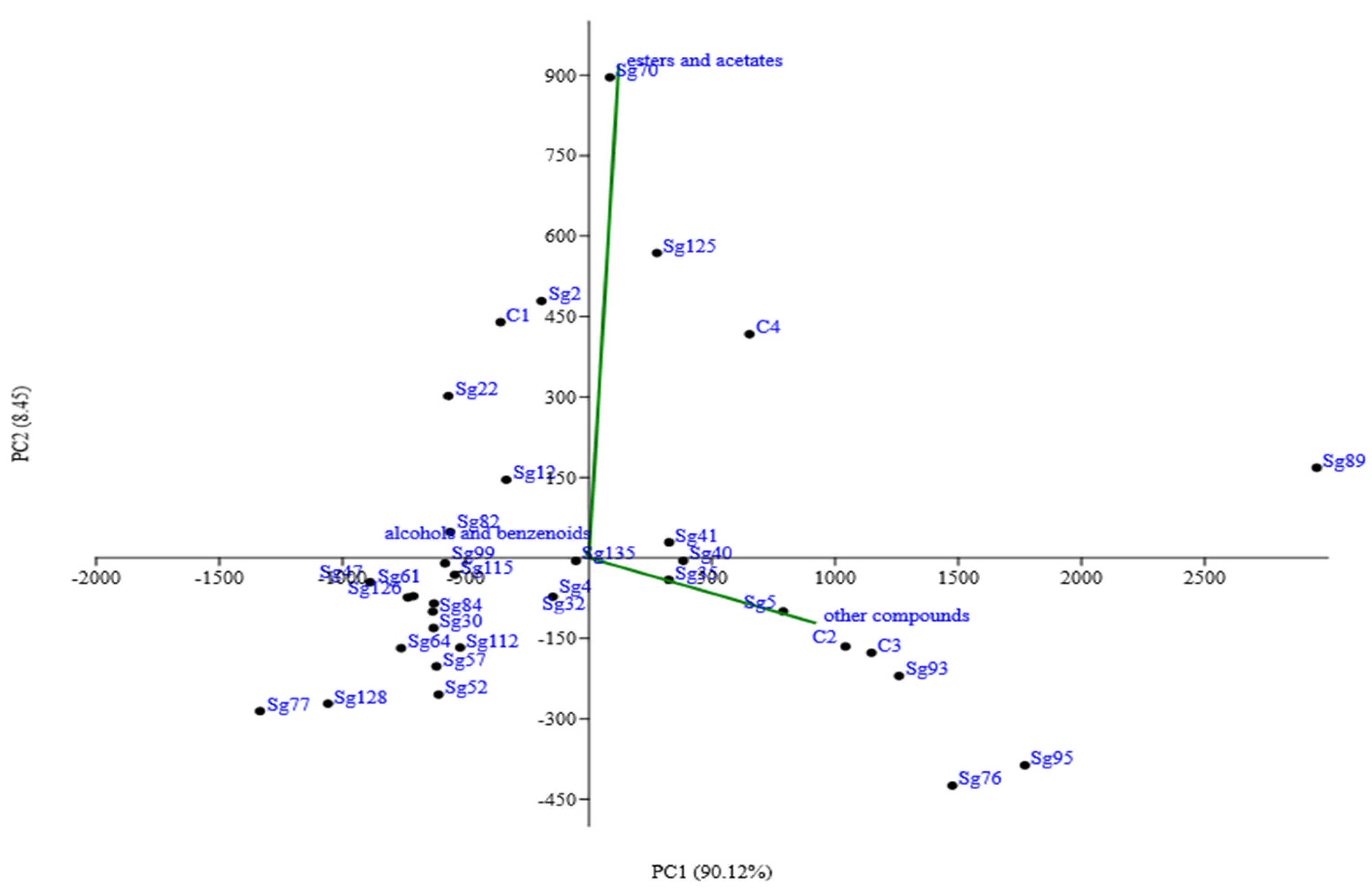
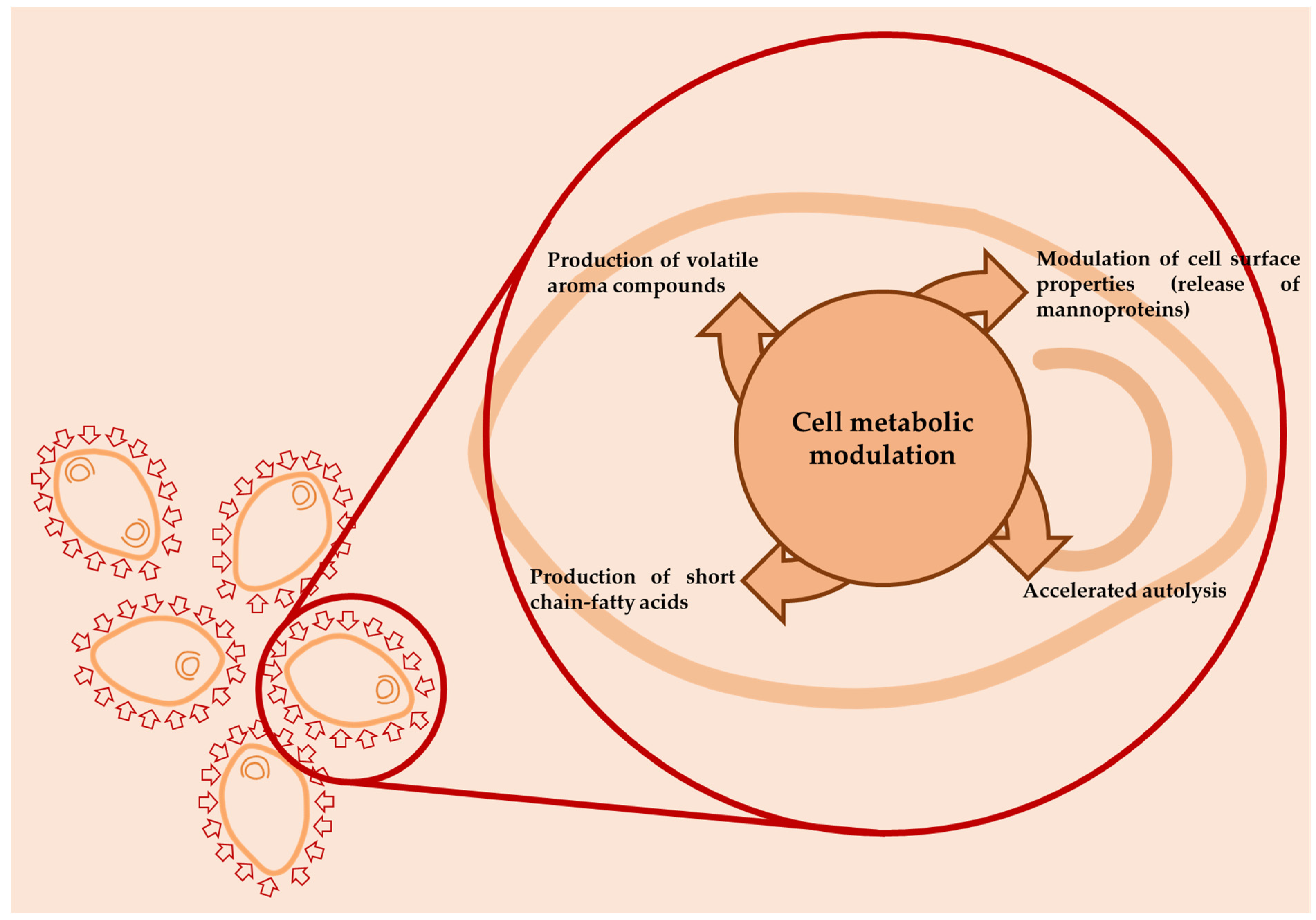
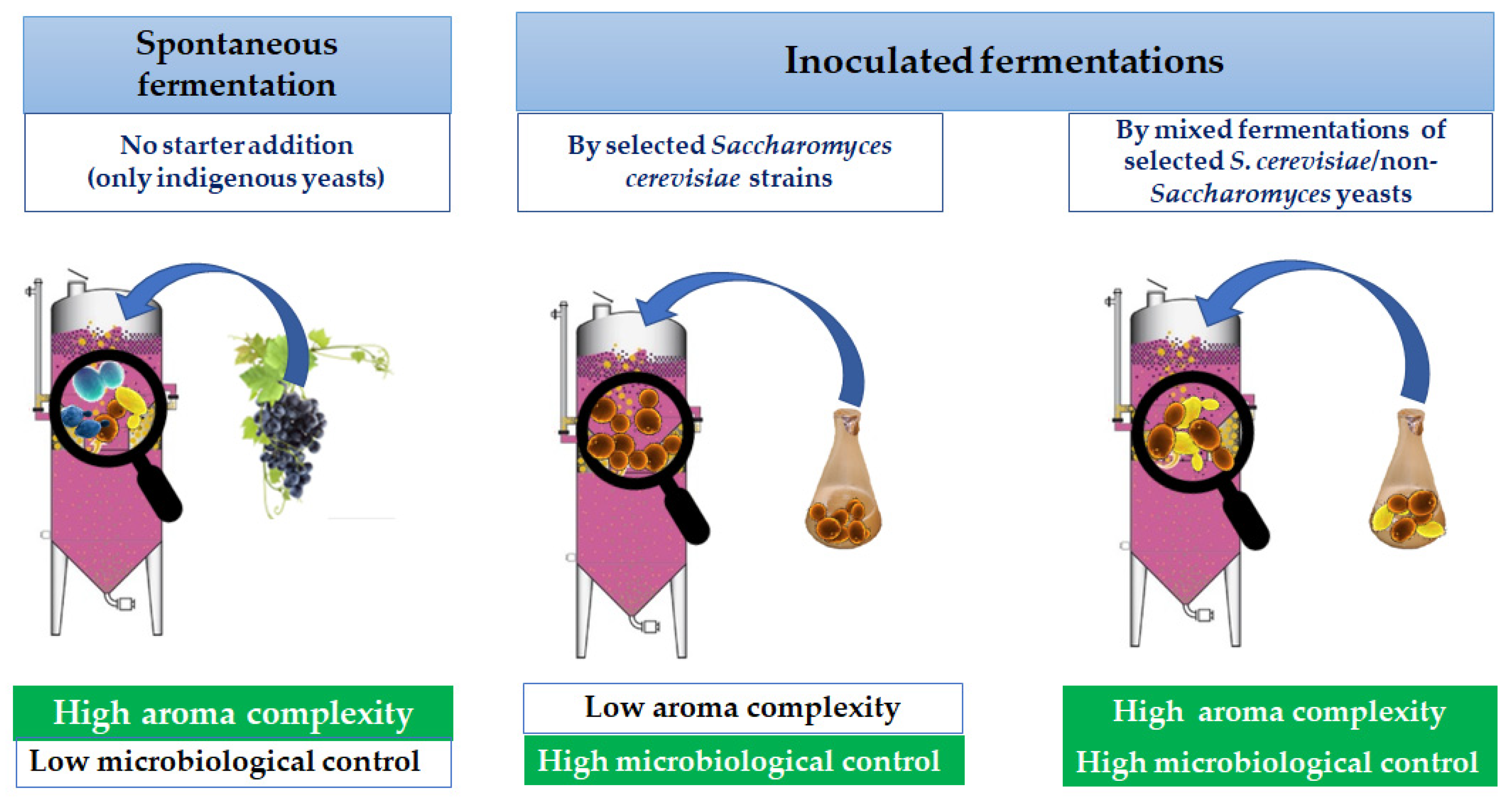
3. Aromatic Compounds and Sensory Profiles of Wines in Relation to the Interactions between Saccharomyces and Non-Saccharomyces Yeasts

{kind=link}
{kind=link}
{kind=link}
{kind=link}
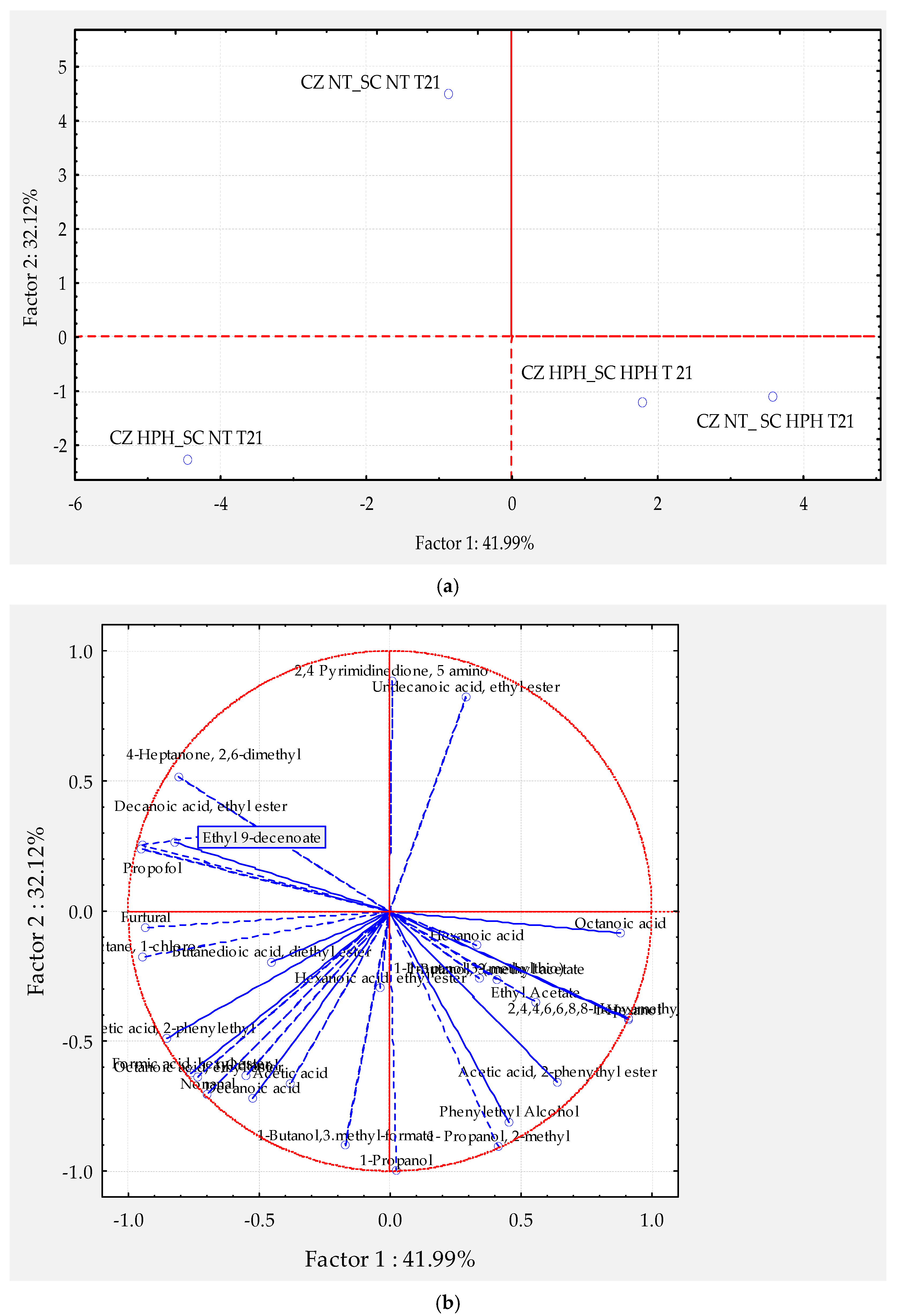{kind=link}
{kind=link}
| Yeast | Characteristics |
|---|---|
| Torulaspora delbrueckii | Low production of acetic acid [83] Increase in glycerol content [84,86] High production of diacetyl, ethyl lactate, and ethyl acetate [87] |
| Lachancea thermotolerans | Production of lactic acid during its fermentative metabolism [97] Increase in the anthocyanins color intensity [93] Reduction in volatile acidity [98] Low production of acetic acid [99,100] Moderate ethanol resistance [99,102] |
| Metschnikowia pulcherrima | High β-glucosidase activity [77] Release of varietal thiols [93] Improvement in some esters [91,103] |
| Schizosaccharomyces pombe | Deacidification of musts metabolizing malic acid [91] High release of polysaccharides [93,107,108] High production of acetic acid [108] |
| Pichia kluyveri | Release of flavour precursors from grape juice [103] Increase in terpene concentration [93,103] |
| Starmerella bacillaris | High production of glycerol [111] Fructophilic character [113] High ethanol tolerance [114] |
| Hanseniasporaspp. (H. guilliermondii, H. uvarum, H. vinae) | Production of β-glucosidase, β-xylosidase, protease [93] High production of 2-phenylethyl acetate [74] Low production of higher alcohols [93,103] |
| Wickerhamomyces anomalus | High production of β-glucosidases, β-D-xylosidases, monoterpenes [112] High production of fruity acetate esters [115] High production of ethyl acetate [116] |
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Perestrelo, R.; Silva, C.; Câmara, J.S. Madeira wine volatile profile. A platform to establish madeira wine aroma descriptors. Molecules 2019, 24, 3028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestres, M.; Busto, O.; Guasch, J. Analysis of organic sulfur compounds in wine aroma. J. Chromatogr. 2000, 881, 569–581. [Google Scholar] [CrossRef]
- López, R.; Aznar, M.; Cacho, J.; Ferreira, V. Determination of minor and trace volatile compounds in wine by solid-phase extraction and gas chromatography with mass spectrometric detection. J. Chromatogr. 2002, 966, 167–177. [Google Scholar] [CrossRef]
- Román, S.M.-S.; Rubio-Bretón, P.; Pérez-Álvarez, E.P.; Garde-Cerdán, T. Advancement in analytical techniques for the extraction of grape and wine volatile compounds. Food Res. Int. 2020, 137, 109712. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Du, B.; Li, J. Aroma compounds in wine. In Grape and Wine Biotechnology; IntechOpen: Rijeka, Croatia, 2016; pp. 273–283. [Google Scholar]
- Chen, L.; Capone, D.L.; Jeffery, D.W. Analysis of potent odour-active volatile thiols in foods and beverages with a focus on wine. Molecules 2019, 24, 2472. [Google Scholar] [CrossRef] [Green Version]
- Perestrelo, R.; Silva, C.; Gonçalves, C.; Castillo, M.; Câmara, J.S. An approach of the madeira wine chemistry. Beverages 2020, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Capece, A.; Romano, P. Yeasts and their metabolic impact on wine flavour. In Yeasts in the Production of Wine; Springer: New York, NY, USA, 2019; pp. 43–80. [Google Scholar] [CrossRef]
- Carpena, M.; Pereira, A.G.; Prieto, M.A.; Simal-Gandara, J. Wine aging technology: Fundamental role of wood barrels. Foods 2020, 9, 1160. [Google Scholar] [CrossRef]
- Molina, A.M.; Swiegers, J.H.; Varela, C.; Pretorius, I.S.; Agosin, E. Influence of wine fermentation temperature on the synthesis of yeast-derived volatile aroma compounds. Appl. Microbiol. Biotechnol. 2007, 77, 675–687. [Google Scholar] [CrossRef]
- Slaghenaufi, D.; Indorato, C.; Troiano, E.; Luzzini, G.; Felis, G.E.; Ugliano, M. Fate of grape-derived terpenoids in model systems containing active yeast cells. J. Agric. Food Chem. 2020, 68, 13294–13301. [Google Scholar] [CrossRef]
- Eldarov, M.; Kishkovskaia, S.; Tanaschuk, T.; Mardanov, A. Genomics and biochemistry of Saccharomyces cerevisiae wine yeast strains. Biochemistry 2016, 81, 1650–1668. [Google Scholar] [CrossRef]
- Bauer, F.; Pretorius, I.S. Yeast stress response and fermentation efficiency: How to survive the making of wine. S. Afr. J. Enol. Vitic. 2000, 21, 27–51. [Google Scholar] [CrossRef]
- Matallana, E.; Aranda, A. Biotechnological impact of stress response on wine yeast. Lett. Appl. Microbiol. 2017, 64, 103–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mina, M.; Tsaltas, D. Contribution of yeast in wine aroma and flavour. Yeast Ind. Appl. 2017, 5, 117–134. [Google Scholar] [CrossRef] [Green Version]
- Robinson, A.L.; Boss, P.K.; Solomon, P.S.; Trengove, R.D.; Heymann, H.; Ebeler, S.E. Origins of grape and wine aroma. Part 1. Chemical components and viticultural impacts. Am. J. Enol. Vitic. 2014, 65, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Ruiz, J.; Esteban-Fernández, A.; Navascués, E.; Marquina, D.; Santos, A.; Moreno-Arribas, M.V. Microbial contribution to wine aroma and its intended use for wine quality improvement. Molecules 2017, 22, 189. [Google Scholar] [CrossRef] [Green Version]
- Hirst, M.B.; Richter, C.L. Review of aroma formation through metabolic pathways of Saccharomyces cerevisiae in beverage fermentations. Am. J. Enol. Vitic. 2016, 67, 361–370. [Google Scholar] [CrossRef] [Green Version]
- Benito, S. Combined Use of Lachancea thermotolerans and Schizosaccharomyces pombe in Winemaking: A Review. Microorganisms 2020, 8, 655. [Google Scholar] [CrossRef]
- Carpena, M.; Fraga-Corral, M.; Otero, P.; Nogueira, R.A.; Garcia-Oliveira, P.; Prieto, M.A.; Simal-Gandara, J. Secondary aroma: Influence of wine microorganisms in their aroma profile. Foods 2021, 10, 51. [Google Scholar] [CrossRef]
- Ugliano, M.; Henschke, P.A. Yeasts and Wine Flavour. In Wine Chemistry and Biochemistry; Moreno-Arribas, M.V., Polo, M.C., Eds.; Springer: New York, NY, USA, 2009; pp. 313–392. [Google Scholar]
- Hazelwood, L.A.; Daran, J.-M.; Van Maris, A.J.; Pronk, J.T.; Dickinson, J.R. The Ehrlich pathway for fusel alcohol production: A century of research on Saccharomyces cerevisiae metabolism. Appl. Environ. Microbiol. 2008, 74, 2259–2266. [Google Scholar] [CrossRef] [Green Version]
- Genovese, A.; Lamorte, S.A.; Gambuti, A.; Moio, L. Aroma of Aglianico and Uva di Troia grapes by aromatic series. Food Res. Int. 2013, 53, 15–23. [Google Scholar] [CrossRef]
- Romano, P.; Pietrafesa, R.; Romaniello, R.; Zambuto, M.; Calabretti, A.; Capece, A. Impact of yeast starter formulations on the production of volatile compounds during wine fermentation. Yeast 2015, 32, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Masneuf-Pomarède, I.; Bely, M.; Marullo, P.; Lonvaud-Funel, A.; Dubourdieu, D. Reassessment of phenotypic traits for Saccharomyces bayanus var. uvarum wine yeast strains. Int. J. Food Microbiol. 2010, 139, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Lopes, C.A.; Rodríguez, M.E.; Sangorrín, M.; Querol, A.; Caballero, A.C. Patagonian wines: The selection of an indigenous yeast starter. J. Ind. Microbiol. Biotechnol. 2007, 34, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Carrau, F.; Gaggero, C.; Aguilar, P.S. Yeast diversity and native vigor for flavor phenotypes. Trends Biotechnol. 2015, 33, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Romaniello, R.; Pietrafesa, R.; Romano, P. Indigenous Saccharomyces cerevisiae yeasts as a source of biodiversity for the selection of starters for specific fermentations. BIO Web Conf. 2014, 3, 02003. [Google Scholar] [CrossRef] [Green Version]
- Romano, P.; Capece, A.; Serafino, V.; Romaniello, R.; Poeta, C. Biodiversity of wild strains of Saccharomyces cerevisiae as tool to complement and optimize wine quality. World J. Microbiol. Biotechnol. 2008, 24, 1797–1802. [Google Scholar] [CrossRef]
- Lambrechts, M.; Pretorius, I. Yeast and its importance to wine aroma-a review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Ehrlich, F. Über die Bedingungen der Fuselölbildung und über ihren Zusammenhang mit dem Eiweissaufbau der Hefe. Ber. Dtsch. Chem. Ges. 1907, 40, 1027–1047. [Google Scholar] [CrossRef] [Green Version]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [CrossRef]
- Ugliano, M.; Henschke, P.A.; Herderich, M.J.; Pretorius, I.S. Nitrogen management is critical for wine flavour and style. Wine Ind. J. 2007, 22, 24–30. [Google Scholar]
- Vilanova, M.; Pretorius, I.S.; Henschke, P.A. Influence of diammonium phosphate addition to fermentation on wine biologicals. In Processing and Impact on Active Components in Food; Elsevier: Amsterdam, The Netherlands, 2015; pp. 483–491. [Google Scholar]
- Carrau, F.M.; Medina, K.; Farina, L.; Boido, E.; Henschke, P.A.; Dellacassa, E. Production of fermentation aroma compounds by Saccharomyces cerevisiae wine yeasts: Effects of yeast assimilable nitrogen on two model strains. FEMS Yeast Res. 2008, 8, 1196–1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swigers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorious, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Odor & Flavor Detection Thresholds in Water (In Parts per Billion). Available online: http://www.leffingwell.com/odorthre.htm (accessed on 13 June 2022).
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial modulation of aromatic esters in wine: Current knowledge and future prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Saerens, S.; Pretorius, I.S. Novel yeast strains as tools for adjusting the flavor of fermented beverages to market specifications. Biotechnol. Flavor Prod. 2016, 3, 62–132. [Google Scholar] [CrossRef]
- Styger, G.; Prior, B.; Bauer, F.F. Wine flavor and aroma. J. Ind. Microbiol. Biotechnol. 2011, 38, 1145. [Google Scholar] [CrossRef] [PubMed]
- Saerens, S.M.; Delvaux, F.R.; Verstrepen, K.J.; Thevelein, J.M. Production and biological function of volatile esters in Saccharomyces cerevisiae. Microb. Biotechnol. 2010, 3, 165–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.Q.; Pilone, G.J. An overview of formation and roles of acetaldehyde in winemaking with emphasis on microbiological implications. Int. J. Food Sci. Technol. 2000, 35, 49–61. [Google Scholar] [CrossRef]
- Kłosowski, G.; Mikulski, D.; Rolbiecka, A.; Czupryński, B. Changes in the Concentration of Carbonyl Compounds during the Alcoholic Fermentation Process Carried out with Saccharomyces cerevisiae Yeast. Pol. J. Microbiol. 2017, 66, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Romano, P.; Suzzi, G.; Turbanti, L.; Polsinelli, M. Acetaldehyde production in Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 1994, 118, 213–218. [Google Scholar] [CrossRef]
- Pereira, A.G.; Jesus, S.-G.; Maria, F.; Paula, G.-O.; Maria, C.; Cecilia, J.-L.; Catarina, L.-L.; Lillian, B.; CFR Ferreira, I.; Angel, P. Chapter Management of Wine Aroma Compounds: Principal Basis and Future Perspectives; IntechOpen: Vienna, Austria, 2020. [Google Scholar]
- Roldán, A.M.; Sánchez-García, F.; Pérez-Rodríguez, L.; Palacios, V.M. Influence of different vinification techniques on volatile compounds and the aromatic profile of Palomino fino wines. Foods 2021, 10, 453. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Siesto, G.; Capece, A.; Pietrafesa, R.; Lanciotti, R.; Patrignani, F.; Granchi, L.; Galli, V.; Bevilacqua, A.; Campaniello, D. Validation of a Standard Protocol to Assess the Fermentative and Chemical Properties of Saccharomyces cerevisiae Wine Strains. Front. Microbiol. 2022, 13, 830277. [Google Scholar] [CrossRef] [PubMed]
- Patrignani, F.; Montanari, C.; Serrazanetti, D.I.; Braschi, G.; Vernocchi, P.; Tabanelli, G.; Parpinello, G.P.; Versari, A.; Gardini, F.; Lanciotti, R. Characterisation of yeast microbiota, chemical and sensory properties of organic and biodynamic Sangiovese red wines. Ann. Microbiol. 2017, 67, 99–109. [Google Scholar] [CrossRef]
- Braschi, G.; Ricci, A.; Grazia, L.; Versari, A.; Patrignani, F.; Lanciotti, R. Mannoprotein content and volatile molecule profiles of trebbiano wines obtained by Saccharomyces cerevisiae and Saccharomyces bayanus strains. Fermentation 2019, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Costa, G.P.; Nicolli, K.P.; Welke, J.E.; Manfroi, V.; Zini, C.A. Volatile profile of sparkling wines produced with the addition of mannoproteins or lees before second fermentation performed with free and immobilized yeasts. J. Braz. Chem. Soc. 2018, 29, 1866–1875. [Google Scholar] [CrossRef]
- Zhang, W.; Du, G.; Zhou, J.; Chen, J. Regulation of sensing, transportation, and catabolism of nitrogen sources in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2018, 82, e00040-17. [Google Scholar] [CrossRef] [Green Version]
- Torrea, D.; Varela, C.; Ugliano, M.; Ancin-Azpilicueta, C.; Francis, I.L.; Henschke, P.A. Comparison of inorganic and organic nitrogen supplementation of grape juice–Effect on volatile composition and aroma profile of a Chardonnay wine fermented with Saccharomyces cerevisiae yeast. Food Chem. 2011, 127, 1072–1083. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. The Yeasts: A Taxonomic Study; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Barbosa, C.; Falco, V.; Mendes-Faia, A.; Mendes-Ferreira, A. Nitrogen addition influences formation of aroma compounds, volatile acidity and ethanol in nitrogen deficient media fermented by Saccharomyces cerevisiae wine strains. J. Biosci. Bioeng. 2009, 108, 99–104. [Google Scholar] [CrossRef]
- Molina, A.M.; Guadalupe, V.; Varela, C.; Swiegers, J.H.; Pretorius, I.S.; Agosin, E. Differential synthesis of fermentative aroma compounds of two related commercial wine yeast strains. Food Chem. 2009, 117, 189–195. [Google Scholar] [CrossRef]
- Parpinello, G.P.; Ricci, A.; Folegatti, B.; Patrignani, F.; Lanciotti, R.; Versari, A. Unraveling the potential of cryotolerant Saccharomyces eubayanus in Chardonnay white wine production. LWT 2020, 134, 110183. [Google Scholar] [CrossRef]
- Magalhães, F.; Krogerus, K.; Castillo, S.; Ortiz-Julien, A.; Dequin, S.; Gibson, B. Exploring the potential of Saccharomyces eubayanus as a parent for new interspecies hybrid strains in winemaking. FEMS Yeast Res. 2017, 17, foy052. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.V.M.; van Wyk, S. Emerging Non-Thermal Technologies as Alternative to SO2 for the Production of Wine. Foods 2021, 10, 2175. [Google Scholar] [CrossRef] [PubMed]
- Santamera, A.; Escott, C.; Loira, I.; del Fresno, J.M.; González, C.; Morata, A. Pulsed Light: Challenges of a Non-Thermal Sanitation Technology in the Winemaking Industry. Beverages 2020, 6, 45. [Google Scholar] [CrossRef]
- Serrazanetti, D.I.; Patrignani, F.; Russo, A.; Vannini, L.; Siroli, L.; Gardini, F.; Lanciotti, R. Cell membrane fatty acid changes and desaturase expression of Saccharomyces bayanus exposed to high pressure homogenization in relation to the supplementation of exogenous unsaturated fatty acids. Front. Microbiol. 2015, 6, 1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patrignani, F.; Ndagijimana, M.; Vernocchi, P.; Gianotti, A.; Riponi, C.; Gardini, F.; Lanciotti, R. High-Pressure Homogenization to Modify Yeast Performance for Sparkling Wine Production According to Traditional Methods. Am. J. Enol. Vitic. 2013, 64, 258–267. [Google Scholar] [CrossRef]
- Lanciotti, R.; Patrignani, F.; Iucci, L.; Saracino, P.; Guerzoni, M.E. Potential of high pressure homogenization in the control and enhancement of proteolytic and fermentative activities of some Lactobacillus species. Food Chem. 2007, 102, 542–550. [Google Scholar] [CrossRef]
- Gottardi, D.; Siroli, L.; Braschi, G.; Rossi, S.; Serrazanetti, D.I.; Patrignani, F.; Lanciotti, R. Sublethal HPH treatment, a sustainable tool that induces autolytic-like process on the early gene expression of Saccharomyces cerevisiae. Food Res. Int. 2022; in press. [Google Scholar]
- Braschi, G.; D’alessandro, M.; Gottardi, D.; Siroli, L.; Patrignani, F.; Lanciotti, R. Effects of sub lethal high pressure homogenization treatment on adhesion mechanisms and stress response genes in Lactobacillus acidophilus 08. Front. Microbiol. 2021, 12, 1202. [Google Scholar] [CrossRef]
- Torresi, S.; Frangipane, M.T.; Anelli, G. Biotechnologies in sparkling wine production. Interesting approaches for quality improvement: A review. Food Chem. 2011, 129, 1232–1241. [Google Scholar] [CrossRef]
- Fornairon-Bonnefond, C.; Camarasa, C.; Moutounet, M.; Salmon, J.-M. New trends on yeast autolysis and wine ageing on lees: A bibliographic review. OENO One 2002, 36, 49–69. [Google Scholar] [CrossRef] [Green Version]
- Patrignani, F.; Vannini, L.; Kamdem, S.L.S.; Lanciotti, R.; Guerzoni, M.E. Potentialities of high-pressure homogenization to inactivate Zygosaccharomyces bailii in fruit juices. J. Food Sci. 2010, 75, M116–M120. [Google Scholar] [CrossRef]
- Gallardo-Chacon, J.J.; Vichi, S.; Lopez-Tamames, E.; Buxaderas, S. Changes in the sorption of diverse volatiles by Saccharomyces cerevisiae lees during sparkling wine aging. J. Agric. Food Chem. 2010, 58, 12426–12430. [Google Scholar] [CrossRef] [PubMed]
- Pradelles, R.; Chassagne, D.; Vichi, S.; Gougeon, R.; Alexandre, H. (−) Geosmin sorption by enological yeasts in model wine and FTIR spectroscopy characterization of the sorbent. Food Chem. 2010, 120, 531–538. [Google Scholar] [CrossRef]
- Iucci, L.; Patrignani, F.; Vallicelli, M.; Guerzoni, M.E.; Lanciotti, R. Effects of high pressure homogenization on the activity of lysozyme and lactoferrin against Listeria monocytogenes. Food Control 2007, 18, 558–565. [Google Scholar] [CrossRef]
- Comuzzo, P.; Calligaris, S.; Iacumin, L.; Ginaldi, F.; Paz, A.E.P.; Zironi, R. Potential of high pressure homogenization to induce autolysis of wine yeasts. Food Chem. 2015, 185, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Morata, A.; Escott, C.; Bañuelos, M.A.; Loira, I.; del Fresno, J.M.; González, C.; Suárez-Lepe, J.A. Contribution of Non-Saccharomyces Yeasts to Wine Freshness. A Review. Biomolecules 2020, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Martin, V.; Valera, M.J.; Medina, K.; Boido, E.; Carrau, F. Oenological Impact of the Hanseniaspora/Kloeckera Yeast Genus on Wines—A Review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef] [Green Version]
- Tufariello, M.; Fragasso, M.; Pico, J.; Panighel, A.; Castellarin, S.D.; Flamini, R.; Grieco, F. Influence of Non-Saccharomyces on Wine Chemistry: A Focus on Aroma-Related Compounds. Molecules 2021, 26, 644. [Google Scholar] [CrossRef]
- Fia, G.; Giovani, G.; Rosi, I. Study of β-glucosidase production by wine-related yeasts during alcoholic fermentation. A new rapid fluorimetric method to determine enzymatic activity. J. Appl. Microbiol. 2005, 99, 509–517. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef] [PubMed]
- De Orduña, R.M. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Ruiz, J.; Belda, I.; Kiene, F.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Santos, A.; Rauhut, D. Application of Non-Saccharomyces Yeasts in Wine Production. In Non-Conventional Yeasts: From Basic Research to Application; Sibirny, A., Ed.; Springer: Cham, Switzerland, 2019; pp. 75–89. [Google Scholar]
- Sabaghian, S.; Braschi, G.; Vannini, L.; Patrignani, F.; Samsulrizal, N.H.; Lanciotti, R. Isolation and Identification of Wild Yeast from Malaysian Grapevine and Evaluation of Their Potential Antimicrobial Activity against Grapevine Fungal Pathogens. Microorganisms 2021, 9, 2582. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Stoeckle, P.; Masneuf-Pomarède, I.; Dubourdieu, D. Impact of mixed Torulaspora delbrueckii–Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 2008, 122, 312–320. [Google Scholar] [CrossRef]
- González-Royo, E.; Pascual, O.; Kontoudakis, N.; Esteruelas, M.; Esteve-Zarzoso, B.; Mas, A.; Canals, J.M.; Zamora, F. Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 2015, 240, 999–1012. [Google Scholar] [CrossRef]
- Medina-Trujillo, L.; González-Royo, E.; Sieczkowski, N.; Heras, J.; Canals, J.M.; Zamora, F. Effect of sequential inoculation (Torulaspora delbrueckii/Saccharomyces cerevisiae) in the first fermentation on the foaming properties of sparkling wine. Eur. Food Res. Technol. 2017, 243, 681–688. [Google Scholar] [CrossRef]
- Puertas, B.; Jiménez, M.J.; Cantos-Villar, E.; Cantoral, J.M.; Rodríguez, M.E. Use of Torulaspora delbrueckii and Saccharomyces cerevisiae in semi-industrial sequential inoculation to improve quality of Palomino and Chardonnay wines in warm climates. J. Appl. Microbiol. 2017, 122, 733–746. [Google Scholar] [CrossRef]
- Ivit, N.N.; Longo, R.; Kemp, B. The Effect of Non-Saccharomyces and Saccharomyces Non-cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine. Fermentation 2020, 6, 77. [Google Scholar] [CrossRef]
- Azzolini, M.; Tosi, E.; Lorenzini, M.; Finato, F.; Zapparoli, G. Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2015, 31, 277–293. [Google Scholar] [CrossRef]
- Renault, P.; Coulon, J.; Moine, V.; Thibon, C.; Bely, M. Enhanced 3-Sulfanylhexan-1-ol Production in Sequential Mixed Fermentation with Torulaspora delbrueckii/Saccharomyces cerevisiae Reveals a Situation of Synergistic Interaction between Two Industrial Strains. Front. Microbiol. 2016, 7, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial Resources and Enological Significance: Opportunities and Benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renault, P.; Coulon, J.; de Revel, G.; Barbe, J.-C.; Bely, M. Increase of fruity aroma during mixed T. delbrueckii/S. cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 2015, 207, 40–48. [Google Scholar] [CrossRef]
- Benito, Á.; Calderón, F.; Benito, S. The Influence of Non-Saccharomyces Species on Wine Fermentation Quality Parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef] [Green Version]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction of Alcohol Content in Wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Navascués, E.; Marquina, D.; Santos, A.; Calderón, F.; Benito, S. Outlining the influence of non-conventional yeasts in wine ageing over lees. Yeast 2016, 33, 329–338. [Google Scholar] [CrossRef]
- Porter, T.J.; Divol, B.; Setati, M.E. Lachancea yeast species: Origin, biochemical characteristics and oenological significance. Food Res. Int. 2019, 119, 378–389. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Vilela, A. Lachancea thermotolerans, the Non-Saccharomyces Yeast that Reduces the Volatile Acidity of Wines. Fermentation 2018, 4, 56. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Kapsopoulou, K.; Kapaklis, A.; Spyropoulos, H. Growth and Fermentation Characteristics of a Strain of the Wine Yeast Kluyveromyces thermotolerans Isolated in Greece. World J. Microbiol. Biotechnol. 2005, 21, 1599–1602. [Google Scholar] [CrossRef]
- Du Plessis, H.; Du Toit, M.; Nieuwoudt, H.; Van der Rijst, M.; Kidd, M.; Jolly, N. Effect of Saccharomyces, Non-Saccharomyces Yeasts and Malolactic Fermentation Strategies on Fermentation Kinetics and Flavor of Shiraz Wines. Fermentation 2017, 3, 64. [Google Scholar] [CrossRef] [Green Version]
- Kapsopoulou, K.; Mourtzini, A.; Anthoulas, M.; Nerantzis, E. Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 735–739. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Santos, A.; Benito, S. Analytical impact of Metschnikowia pulcherrima in the volatile profile of Verdejo white wines. Appl. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Canonico, L.; Comitini, F.; Ciani, M. Metschnikowia pulcherrima Selected Strain for Ethanol Reduction in Wine: Influence of Cell Immobilization and Aeration Condition. Foods 2019, 8, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domizio, P.; Liu, Y.; Bisson, L.F.; Barile, D. Cell wall polysaccharides released during the alcoholic fermentation by Schizosaccharomyces pombe and S. japonicus: Quantification and characterization. Food Microbiol. 2017, 61, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Benito, S. The impacts of Schizosaccharomyces on winemaking. Appl. Microbiol. Biotechnol. 2019, 103, 4291–4312. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderón, F.; Palmero, D.; Suárez-Lepe, J.A. Physiological features of Schizosaccharomyces pombe of interest in making of white wines. Eur. Food Res. Technol. 2013, 236, 29–36. [Google Scholar] [CrossRef] [Green Version]
- Viana, F.; Gil, J.V.; Vallés, S.; Manzanares, P. Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 2009, 135, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Di Maio, S.; Genna, G.; Gandolfo, V.; Amore, G.; Ciaccio, M.; Oliva, D. Presence of Candida zemplinina in Sicilian musts and selection of a strain for wine mixed fermentations. S. Afr. J. Enol. Vitic. 2012, 33, 80–87. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Magyar, I.; Tóth, T. Comparative evaluation of some oenological properties in wine strains of Candida stellata, Candida zemplinina, Saccharomyces uvarum and Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 94–100. [Google Scholar] [CrossRef]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella bacillaris in winemaking: Opportunities and risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genovés, S.; Vallés, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef]
- Cañas, P.M.I.; García-Romero, E.; Manso, J.M.H.; Fernández-González, M. Influence of sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the quality of red wines. Eur. Food Res. Technol. 2014, 239, 279–286. [Google Scholar] [CrossRef]
- MacDonald, J.B.; Saliba, A.J.; Winetitles, B. Wine choice and drivers of consumption explored in relation to generational cohorts and methodology. J. Retail. Consum. Serv. 2013, 20, 349–357. [Google Scholar] [CrossRef]






| Compound | Concentration in Wine (mg/L) [37] | Aroma Threshold (mg/L) | Aroma Descriptors [37] | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Water | Ethanol (10% v/v) | Wine | Synth. Wine | |||||||
| Ethyl acetate | 22.5–63.5 | 0.005–5 | [38] | 7.5 | [37] | Fruit, solvent | ||||
| Isoamyl acetate | 0.1–3.4 | 0.002 | [38] | 0.03 | [37] | Banana, pear | ||||
| 2-phenylethyl acetate | 0–18.5 | 0.25 | [37] | Floral, fruity, rose | ||||||
| Isobutyl acetate | 0.01–1.6 | 0.066 | [38] | Banana, fruity | ||||||
| Exiyl acetate | 0–4.8 | 0.7 | [37] | Sweet, perfume | ||||||
| Ethyl butanoate | 0.01–1.8 | 0.001 | [38] | 0.02 | [37] | Floral, fruity | ||||
| Ethyl esanoate | 0.03–3.4 | 0.05 | [37] | Green apple | ||||||
| Ethyl octanoate | 0.05–3.8 | 0.02 | [37] | Soap | ||||||
| Ethyl decanoate | 0–2.1 | 0.2 | [37] | Floral, soap | ||||||
| Propanol | 9–6.8 | 9 | [38] | 500 | [37] | Pungent, astringent | ||||
| Butanol | 0.5–8.5 | 0.5 | [38] | 150 | [37] | Alcoholic | ||||
| Isobutanol | 9–174 | 40 | [37] | Alcoholic | ||||||
| Isoamyl alcohol | 6–490 | 0.25–0.3 | [38] | 30 | [37] | Astringent, solvent | ||||
| Hexanol | 0.3–12 | 4 | [37] | Herbaceous | ||||||
| 2- Phenylethyl alcohol | 4–197 | 10 | [37] | Floral, rose | ||||||
| Acetic acid | 100–1150 | 280 | [37] | Acidic, vinegar | ||||||
| Acetaldehyde | 10–75 | 0.015–0.12 | [38] | 100 | [37] | Unripe walnut, bruised fruit, sherry | ||||
| Diacetyl | <5 | 0.0023–0.0065 | 0.2–2.8 | [37] | Buttery | |||||
| Glycerol | 5–14 | 5.2 g/L | [37] | Odorless (slight sweet taste) | ||||||
| Linalool | 0.002–0.01 | 0.006 | [38] | 0.025 | [37] | Rose | ||||
| Geraniol | 0.001–0.044 | 0.04–0.075 | [38] | 30 | [37] | Rose | ||||
| Citronellol | 0.015–0.042 | 0.04–0.086 | [38] | 100 | [37] | Citronella | ||||
| 2-Acetyl-1-pyrroline (ACPY) | traces | 0.0001 | [37] | Mouse urine | ||||||
| 2-Acetyltetrahydropyridine (ACPTY) | 0.005–0.1 | 0.0016 | [37] | Mouse urine | ||||||
| 4-Ethylphenol | 0.012–6.5 | 0.14 | [37] | 0.6 | [37] | Medicinal, stable | ||||
| 4-Ethylguaiacol | 0.001–0.44 | 0.04 | [38] | 0.033 | [37] | 0.11 | [37] | Phenolic, sweet | ||
| 4-Vinylphenol | 0.04–0.45 | 0.02 | [37] | Drug | ||||||
| 4-Vinylguaiacol | 0.001–0.71 | 10 | [37] | Cloves, phenolic | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, P.; Braschi, G.; Siesto, G.; Patrignani, F.; Lanciotti, R. Role of Yeasts on the Sensory Component of Wines. Foods 2022, 11, 1921. https://doi.org/10.3390/foods11131921
Romano P, Braschi G, Siesto G, Patrignani F, Lanciotti R. Role of Yeasts on the Sensory Component of Wines. Foods. 2022; 11(13):1921. https://doi.org/10.3390/foods11131921
Chicago/Turabian StyleRomano, Patrizia, Giacomo Braschi, Gabriella Siesto, Francesca Patrignani, and Rosalba Lanciotti. 2022. "Role of Yeasts on the Sensory Component of Wines" Foods 11, no. 13: 1921. https://doi.org/10.3390/foods11131921
APA StyleRomano, P., Braschi, G., Siesto, G., Patrignani, F., & Lanciotti, R. (2022). Role of Yeasts on the Sensory Component of Wines. Foods, 11(13), 1921. https://doi.org/10.3390/foods11131921