
Chitosan Increases Lysine Content through Amino Acid Transporters in Flammulina filiformis



Abstract
:Highlights
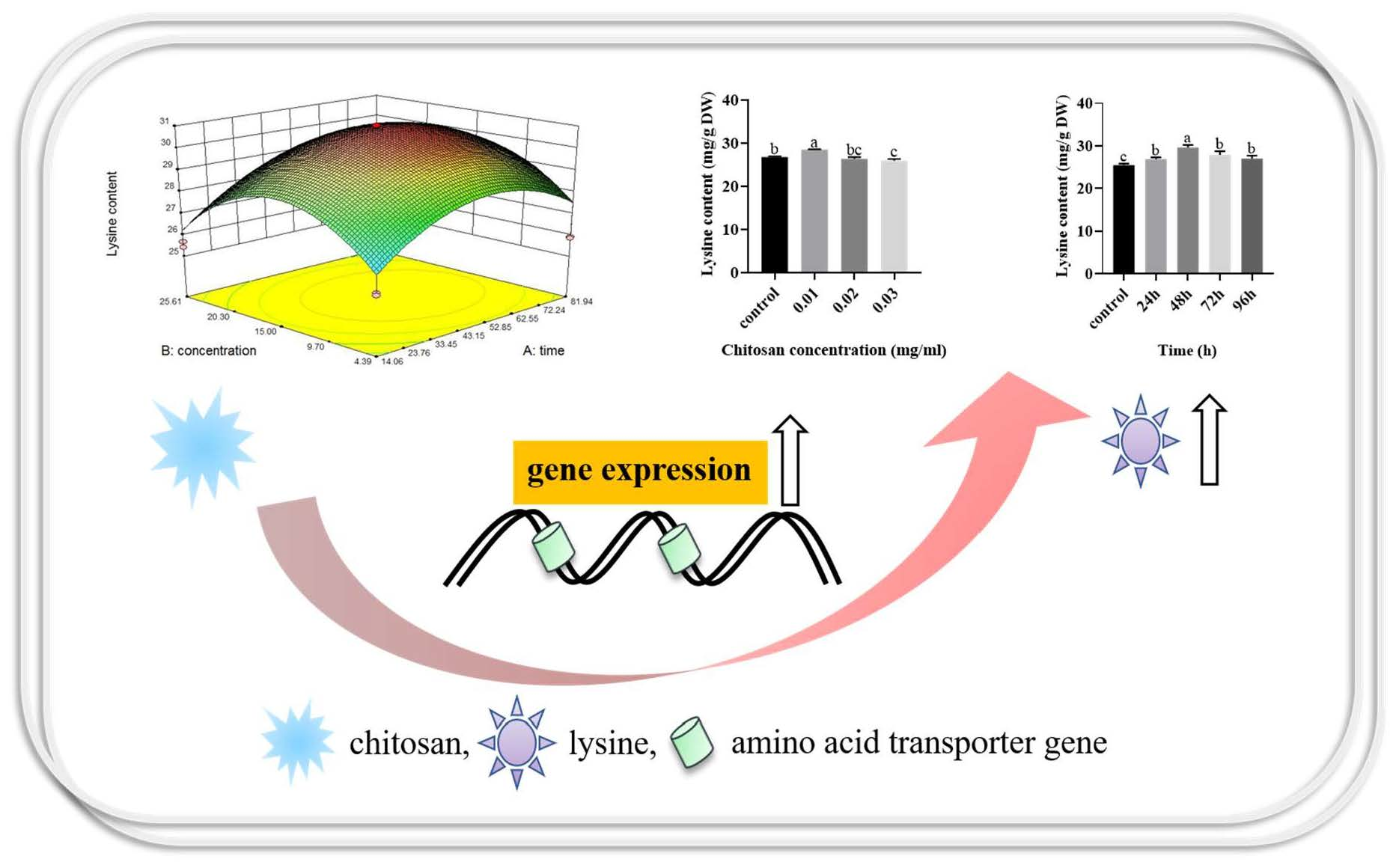
- Chitosan treatment increased the lysine content of F. filiformis. Results from this study provide insights for industrial-scale trials potentially resulting in higher-quality F. filiformis.
- Response surface methodology (RSM) was used to optimize the conditions of chitosan-induced amino acid content in F. filiformis for the first time.
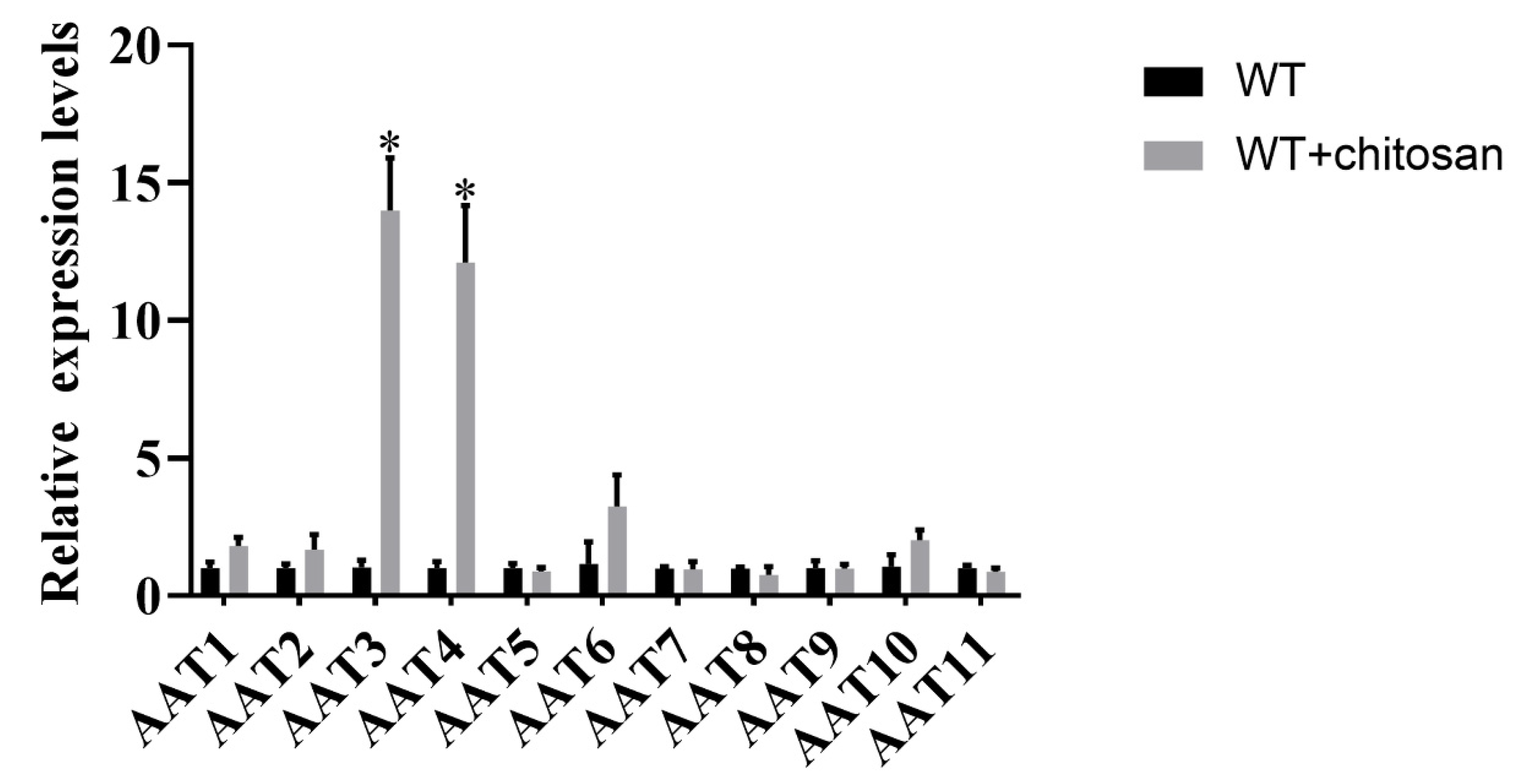
- The amino acid transporters in F. filiformis were systematically screened for the first time. The involvement of AAT3 and AAT4 genes in chitosan-induced lysine biosynthesis in F. filiformis was investigated by gene-silenced strains.
Abstract

1. Introduction
2. Materials and Methods
2.1. Strains, Culture Conditions and Induction Treatment with Chitosan of Mycelium of F. filiformis
2.2. Cultivation and Induction Treatment with Chitosan of Fruiting Body of F. filiformis
2.3. Determination of Mycelial Biomass, Growth of Fruiting Body, Total Protein and Lysine Content
2.4. Optimization for Chitosan-Induced Lysine Accumulation in F. filiformis by Response Surface Methodology
2.5. RNA Extraction and Gene Expression Analysis
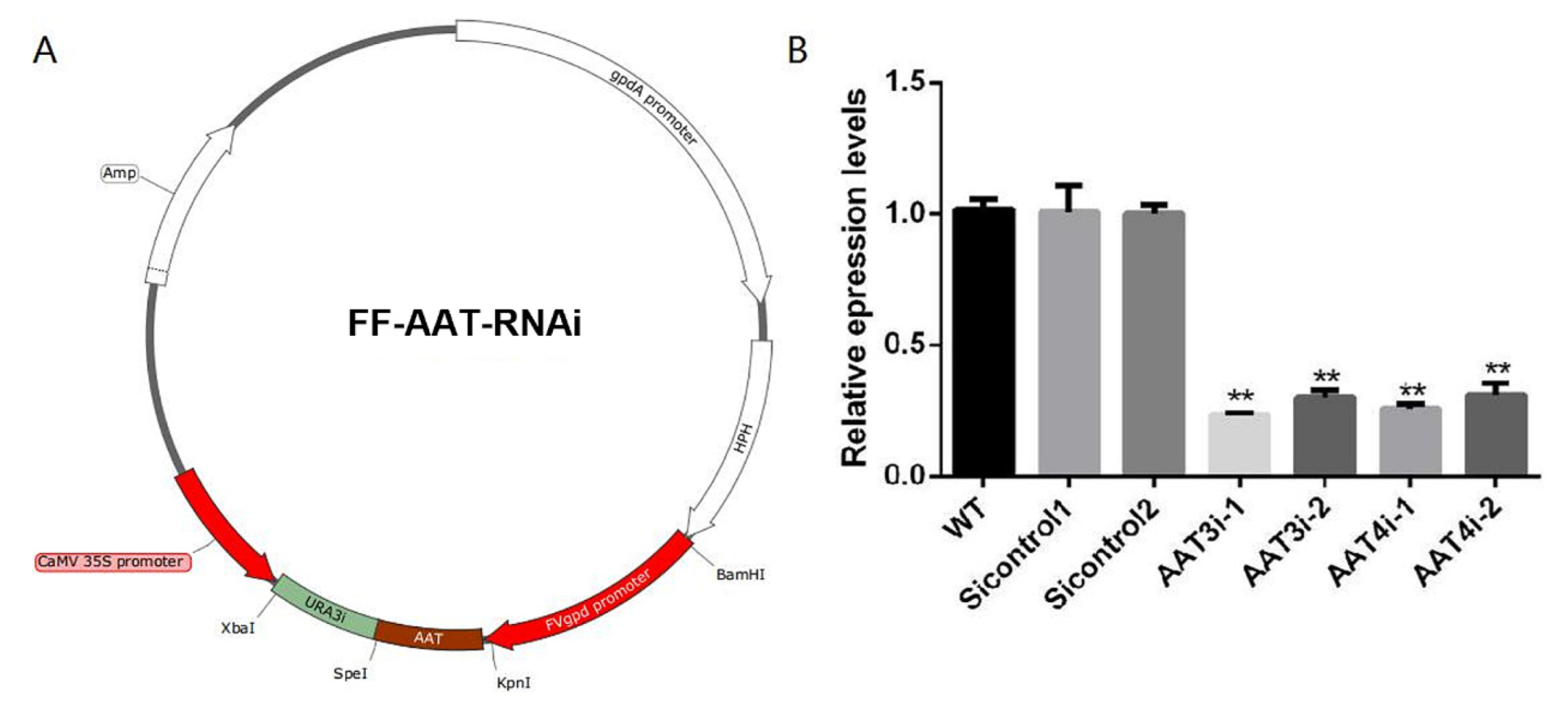
2.6. Construction of an AAT-Silencing Vector and Transformation
2.7. Statistical Analysis
3. Results
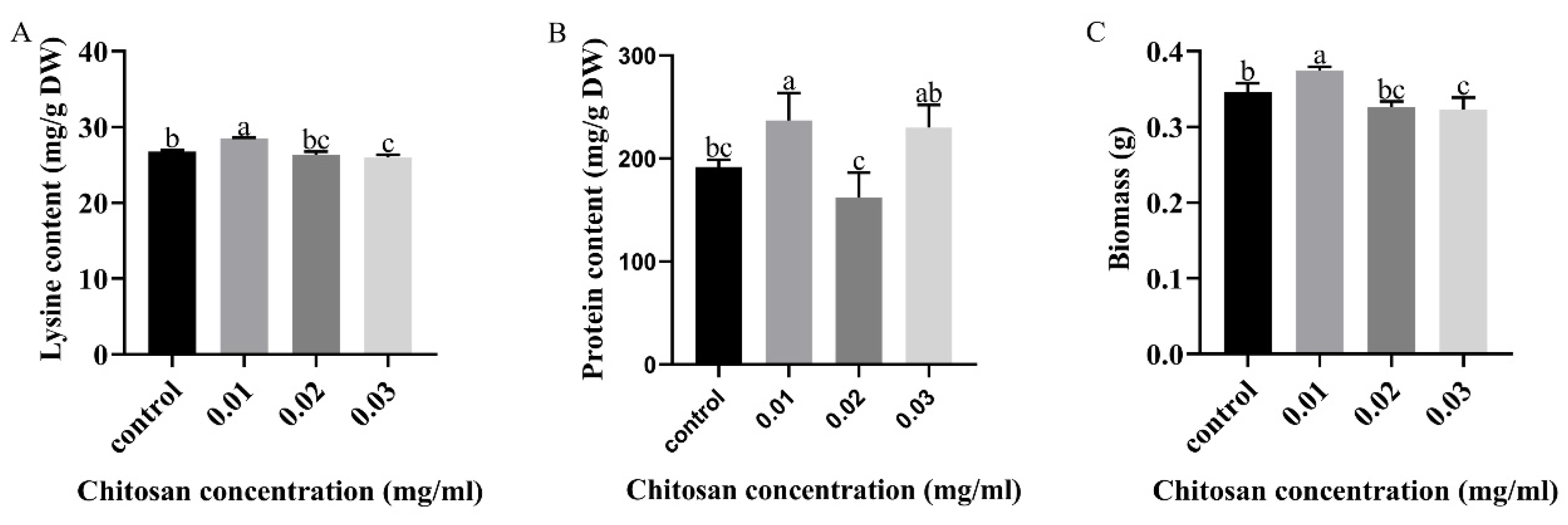
3.1. Effects of Chitosan Treatment for Different Concentrations on Lysine and Protein Content of Mycelium of F. filiformis
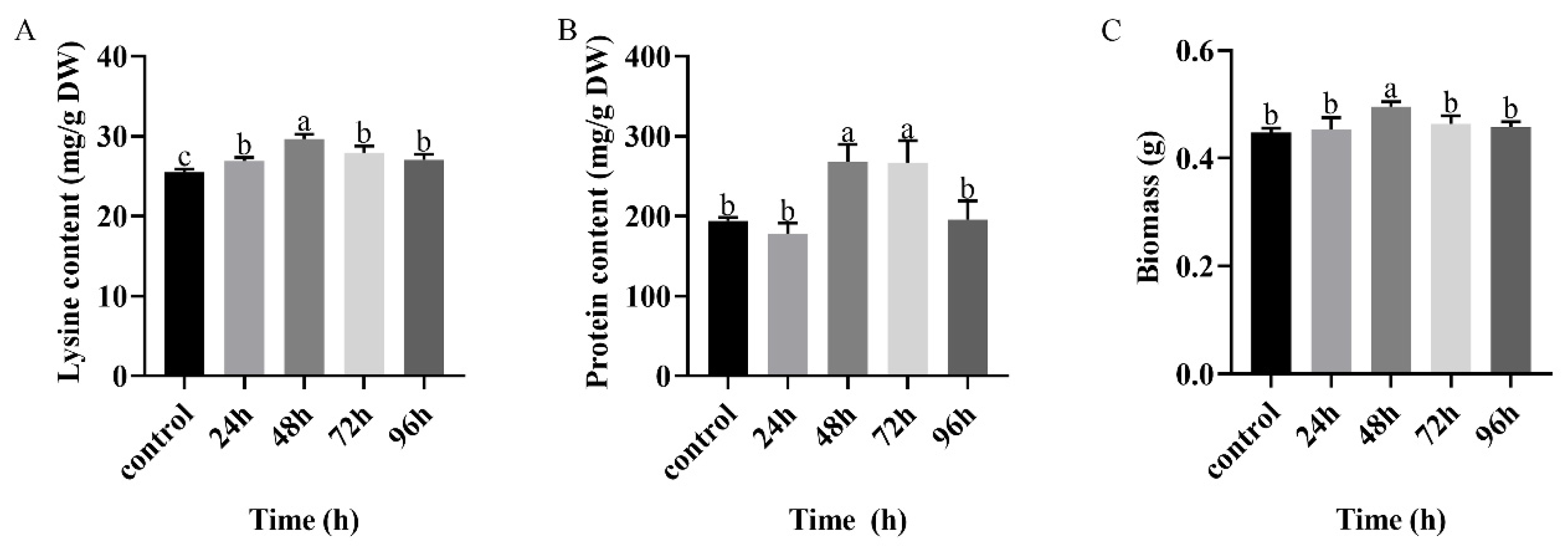
3.2. Effects of Chitosan Treatment Duration on Lysine and Protein Content of Mycelium of F. filiformis
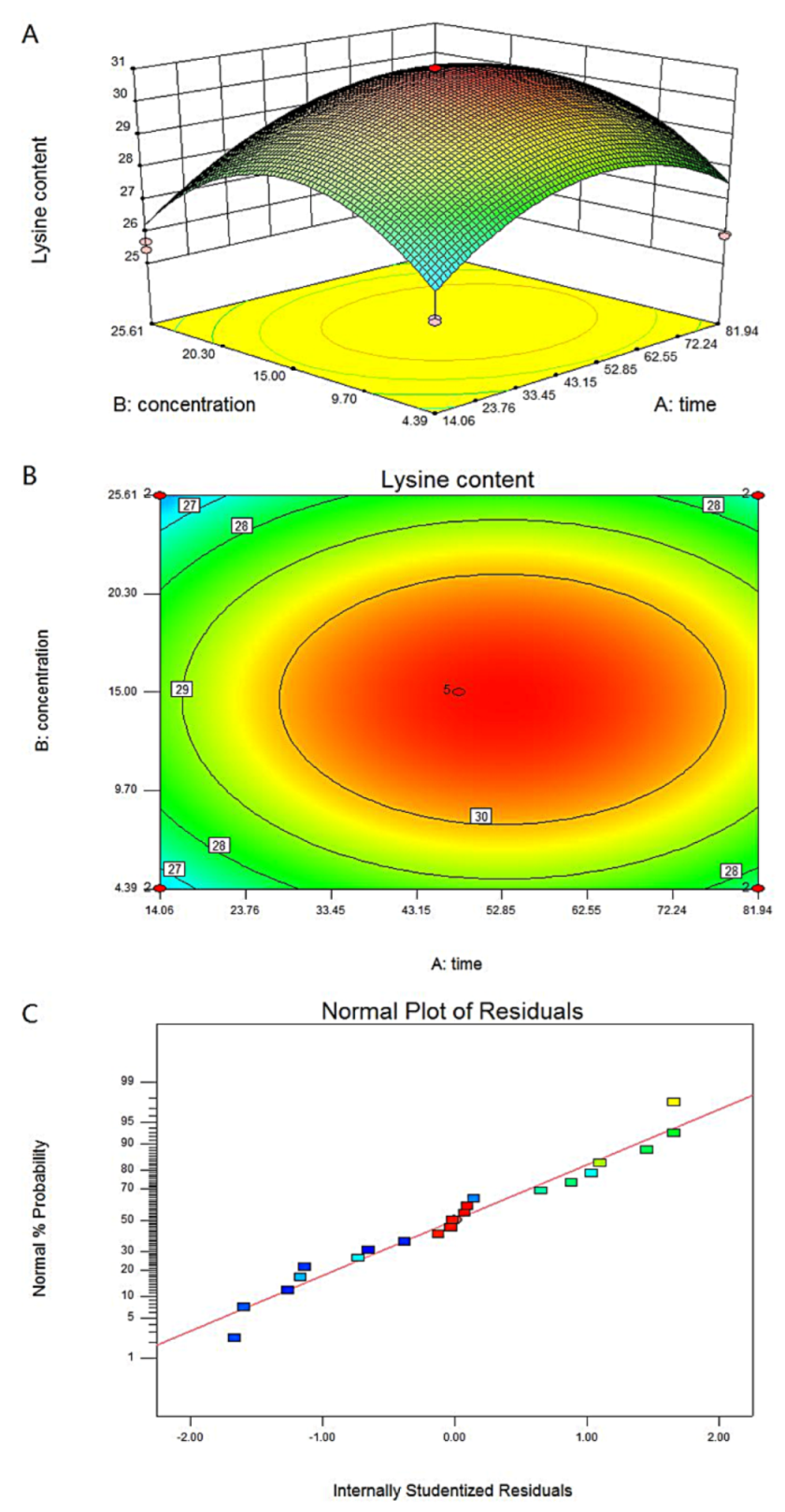
3.3. Response Surface Methodology to Optimize Chitosan Treatment Conditions
3.4. Chitosan Treatment Promotes the Growth of the Fruiting Body of F. filiformis
3.5. Genetic Screening of Amino Acid Transporters Induced by Chitosan
3.6. Construction of AAT3 and AAT4 Genes Silencing Strains of F. filiformis
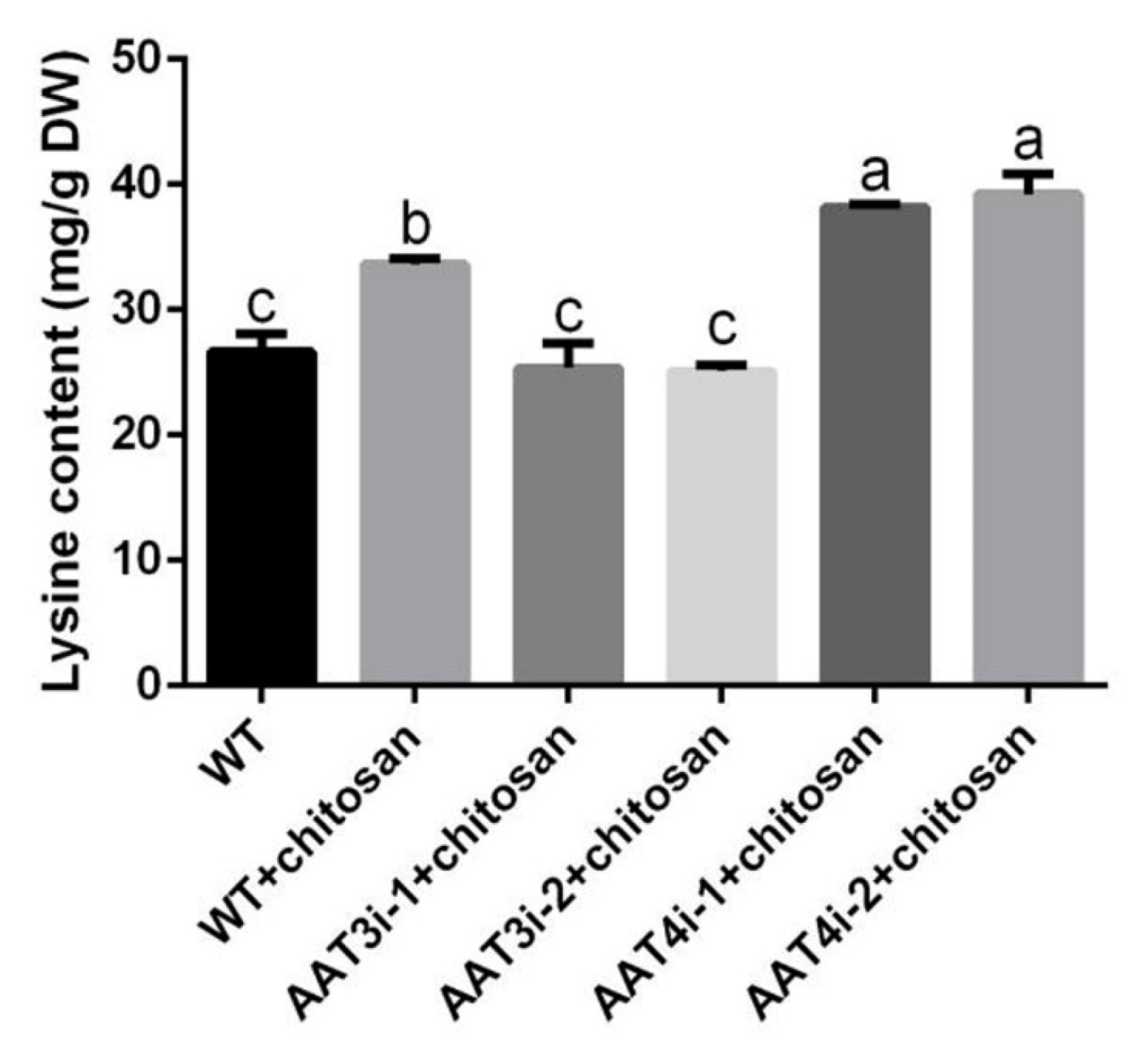
3.7. Changes to Lysine Content in AAT3 and AAT4 Silencing Strains under Optimal Chitosan Induction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, Z.; Xiao, Y.; Wu, H. Selenium accumulation, speciation, and its effect on nutritive value of Flammulina velutipes (Golden needle mushroom). Food Chem. 2021, 350, 128667. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, J.-M.; Tang, Y.-J.; Ma, K.; Li, B.; Zeng, X.; Liu, X.-B.; Li, Y.; Yang, Z.-L.; Xu, W.-N.; et al. Genome-wide analysis and prediction of genes involved in the biosynthesis of polysaccharides and bioactive secondary metabolites in high-temperature-tolerant wild Flammulina filiformis. BMC Genom. 2020, 21, 719. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.K.; Das, A.K.; Banerjee, R.; Pateiro, M.; Nanda, P.K.; Gadekar, Y.P.; Biswas, S.; McClements, D.J.; Lorenzo, J.M. Application of Enoki Mushroom (Flammulina Velutipes) Stem Wastes as Functional Ingredients in Goat Meat Nuggets. Foods 2020, 9, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukushima, M.; Ohashi, T.; Fujiwara, Y.; Sonoyama, K.; Nakano, M. Cholesterol-Lowering Effects of Maitake (Grifola frondosa) Fiber, Shiitake (Lentinus edodes) Fiber, and Enokitake (Flammulina velutipes) Fiber in Rats. Exp. Biol. Med. 2001, 226, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.H.; Liu, X.M.; Chen, Z.Y.; Liao, S.T.; Zou, Y.X. Isolation, purification and identification of nine chemical compounds from Flammulina velutipes fruiting bodies. Food Chem. 2013, 141, 2873–2879. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Carbonero, E.R.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Characterization of a heterogalactan: Some nutritional values of the edible mushroom Flammulina velutipes. Food Chem. 2008, 108, 329–333. [Google Scholar] [CrossRef]
- Xu, H.Y.; Andi, B.; Qian, J.H.; West, A.H.; Cook, P.F. The alpha-aminoadipate pathway for lysine biosynthesis in fungi. Cell Biochem. Biophys. 2006, 46, 43–64. [Google Scholar] [CrossRef]
- Liu, J.Y.; Li, Q.Z.; Jiang, P.Y.; Xu, Z.; Zhang, D.; Zhang, L.J.; Zhang, M.Y.; Yu, H.L.; Song, C.Y.; Tan, Q.; et al. Overexpression of the saccharopine dehydrogenase gene improves lysine biosynthesis in Flammulina velutipes. J. Basic Microbiol. 2019, 59, 890–900. [Google Scholar] [CrossRef]
- Yin, J.; Han, H.; Li, Y.; Liu, Z.; Zhao, Y.; Fang, R.; Huang, X.; Zheng, J.; Ren, W.; Wu, F.; et al. Lysine Restriction Affects Feed Intake and Amino Acid Metabolism via Gut Microbiome in Piglets. Cell. Physiol. Biochem. 2017, 44, 1749–1761. [Google Scholar] [CrossRef]
- Liu, G.-Q.; Zhang, K.-C. Enhancement of polysaccharides production in Ganoderma lucidum by the addition of ethyl acetate extracts from Eupolyphaga sinensis and Catharsius molossus. Appl. Microbiol. Biotechnol. 2007, 74, 572–577. [Google Scholar] [CrossRef]
- Yang, H.-L.; Wu, T.-X.; Zhang, K.-C. Enhancement of mycelial growth and polysaccharide production in Ganoderma lucidum (the Chinese medicinal fungus, ‘Lingzhi’) by the addition of ethanol. Biotechnol. Lett. 2004, 26, 841–844. [Google Scholar] [CrossRef] [PubMed]
- Ren, A.; Li, X.-B.; Miao, Z.-G.; Shi, L.; Jaing, A.-L.; Zhao, M.-W. Transcript and metabolite alterations increase ganoderic acid content in Ganoderma lucidum using acetic acid as an inducer. Biotechnol. Lett. 2014, 36, 2529–2536. [Google Scholar] [CrossRef] [PubMed]
- Kurita, K. Chemistry and application of chitin and chitosan. Polym. Degrad. Stab. 1998, 59, 117–120. [Google Scholar] [CrossRef]
- Karakosta, L.K.; Vatavali, K.A.; Kosma, I.S.; Badeka, A.V.; Kontominas, M.G. Combined Effect of Chitosan Coating and Laurel Essential Oil (Laurus nobilis) on the Microbiological, Chemical, and Sensory Attributes of Water Buffalo Meat. Foods 2022, 11, 1664. [Google Scholar] [CrossRef]
- Jia, X.C.; Rajib, M.R.; Yin, H. Recognition Pattern, Functional Mechanism and Application of Chitin and Chitosan Oligosaccharides in Sustainable Agriculture. Curr. Pharm. Des. 2020, 26, 3508–3521. [Google Scholar] [CrossRef]
- Li, J.H.; Guo, Z.X.; Luo, Y.; Wu, X.M.; An, H.M. Chitosan Can Induce Rosa roxburghii Tratt. against Sphaerotheca sp. and Enhance Its Resistance, Photosynthesis, Yield, and Quality. Horticulturae 2021, 7, 289. [Google Scholar] [CrossRef]
- Hyde, R.; Taylor, P.M.; Hundal, H.S. Amino acid transporters: Roles in amino acid sensing and signalling in animal cells. Biochem. J. 2003, 373, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.P.; Du, G.C.; Zhou, J.W.; Chen, J. Regulation of Sensing, Transportation, and Catabolism of Nitrogen Sources in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2018, 82, e00040-17. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.E.; Song, Z.Y.; Lu, K.; Lian, X.M.; Cai, H.M. Molecular characterization, expression and functional analysis of the amino acid transporter gene family (OsAATs) in rice. Acta Physiol. Plant 2012, 34, 1943–1962. [Google Scholar] [CrossRef]
- Guo, N.; Xue, D.; Zhang, W.; Zhao, J.M.; Xue, C.C.; Yan, Q.; Xue, J.Y.; Wang, H.T.; Zhang, Y.M.; Xing, H. Overexpression of GmProT1 and GmProT2 increases tolerance to drought and salt stresses in transgenic Arabidopsis. J. Integr. Agric. 2016, 15, 1727–1743. [Google Scholar] [CrossRef]
- Peng, B.; Kong, H.; Li, Y.; Wang, L.; Zhong, M.; Sun, L.; Gao, G.; Zhang, Q.; Luo, L.; Wang, G.; et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 2014, 5, 4847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, B.; Andrade, P.B.; Silva, B.M.; Baptista, P.; Seabra, R.M.; Valentão, P. Comparative Study on Free Amino Acid Composition of Wild Edible Mushroom Species. J. Agr. Food Chem. 2008, 56, 10973–10979. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, C.; Xie, B.; Zhang, L.; Yan, S.; Wang, W.; Tao, Y.; Li, S. A Single Transcription Factor (PDD1) Determines Development and Yield of Winter Mushroom (Flammulina velutipes). Appl. Env. Microbiol. 2019, 85, e01735-19. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Martos, S.; Nguyen, H.H.; Nguyen, P.N.; Hristozova, N.; Macossay-Castillo, M.; Kovacs, D.; Bekesi, A.; Oemig, J.S.; Maes, D.; Pauwels, K.; et al. Quantification of Intrinsically Disordered Proteins: A Problem Not Fully Appreciated. Front Mol. Biosci. 2018, 5, 83. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wan, X.-f.; Liu, B.-l.; Yu, T.; Yan, N.; Chai, X.-S.; Li, Y.-m.; Chen, G.-x. Determination of lysine content based on an in situ pretreatment and headspace gas chromatographic measurement technique. Anal. Bioanal. Chem. 2018, 410, 3111–3117. [Google Scholar] [CrossRef]
- Fitzpatrick, W.H. Spectrophotometric Determination of Amino Acids by the Ninhydrin Reaction. Science 1949, 109, 469. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Casta, A.; Wang, R.; Lozada, E.; Fan, W.; Kane, S.; Ge, Q.Y.; Gu, W.; Orren, D.; Luo, J.Y. Regulation of WRN protein cellular localization and enzymatic activities by SIRT1-mediated deacetylation. J. Biol. Chem. 2008, 283, 7590–7598. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shi, L.; Zhang, T.J.; Xu, C.; Ren, A.; Jiang, A.L.; Yu, H.S.; Zhao, M.W. Development of a dual promoter-mediated gene silencing system in Flammulina velutipes. Mycoscience 2017, 58, 181–187. [Google Scholar] [CrossRef]
- Shi, L.; Chen, D.D.; Xu, C.; Ren, A.; Yu, H.S.; Zhao, M.W. Highly-efficient liposome-mediated transformation system for the basidiomycetous fungus Flammulina velutipes. J. Gen. Appl. Microbiol. 2017, 63, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandasamy, P.; Gyimesi, G.; Kanai, Y.; Hediger, M.A. Amino acid transporters revisited: New views in health and disease. Trends Biochem. Sci. 2018, 43, 752–789. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Lou, H.; Hu, J.; Liu, Z.; Chen, Q. Macrofungi: A review of cultivation strategies, bioactivity, and application of mushrooms. Compr Rev. Food Sci. Food Saf. 2020, 19, 2333–2356. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, Y. Influences of environmental factors on fruiting body induction, development and maturation in mushroom-forming fungi. Fungal Biol. Rev. 2018, 32, 236–248. [Google Scholar] [CrossRef]
- Cao, P.F.; Wu, C.G.; Dang, Z.H.; Shi, L.; Jiang, A.L.; Ren, A.; Zhao, M.W. Effects of Exogenous Salicylic Acid on Ganoderic Acid Biosynthesis and the Expression of Key Genes in the Ganoderic Acid Biosynthesis Pathway in the Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes). Int. J. Med. Mushrooms 2017, 19, 65–73. [Google Scholar] [CrossRef]
- Vi, M.; Yang, X.Q.; Zeng, X.L.; Chen, R.A.; Guo, L.Q.; Lin, J.F.; He, Q.Y.; Zheng, Q.W.; Wei, T. Improvement of Nutritional and Bioactive Compound Production by Lion’s Mane Medicinal Mushroom, Hericium erinaceus (Agaricomycetes), by Spraying Growth Regulators. Int. J. Med. Mushrooms 2018, 20, 271–281. [Google Scholar] [CrossRef]
- Yang, W.J.; Wu, Y.Y.; Hu, Q.H.; Pei, F.; Mariga, A.M. Preharvest treatment of Agaricus bisporus with methyl jasmonate inhibits postharvest deterioration. Lwt-Food Sci. Technol. 2019, 106, 158–163. [Google Scholar] [CrossRef]
- Gao, Y.; Wu, Y. Recent advances of chitosan-based nanoparticles for biomedical and biotechnological applications. Int. J. Biol. Macromol. 2022, 203, 379–388. [Google Scholar] [CrossRef]
- Campos, E.V.R.; Oliveira, J.L.; Fraceto, L.F. Poly(ethylene glycol) and Cyclodextrin-Grafted Chitosan: From Methodologies to Preparation and Potential Biotechnological Applications. Front Chem. 2017, 5, 93. [Google Scholar] [CrossRef] [Green Version]
- Sathiyabama, M.; Bernstein, N.; Anusuya, S. Chitosan elicitation for increased curcumin production and stimulation of defence response in turmeric (Curcuma longa L.). Ind. Crops Prod. 2016, 89, 87–94. [Google Scholar] [CrossRef]
- Perez-Balibrea, S.; Moreno, D.A.; Garcia-Viguera, C. Improving the phytochemical composition of broccoli sprouts by elicitation. Food Chem. 2011, 129, 35–44. [Google Scholar] [CrossRef]
- Samarah, N.H.; Al-Quraan, N.A.; Massad, R.S.; Welbaum, G.E. Treatment of bell pepper (Capsicum annuum L.) seeds with chitosan increases chitinase and glucanase activities and enhances emergence in a standard cold test. Sci. Hortic. 2020, 269, 109393. [Google Scholar] [CrossRef]
- Wang, T.; Chen, Y.; Zhang, M.; Chen, J.; Liu, J.; Han, H.; Hua, X. Arabidopsis AMINO ACID PERMEASE1 Contributes to Salt Stress-Induced Proline Uptake from Exogenous Sources. Front Plant Sci. 2017, 8, 2182. [Google Scholar] [CrossRef] [Green Version]
- Santiago, J.P.; Tegeder, M. Connecting Source with Sink: The Role of Arabidopsis AAP8 in Phloem Loading of Amino Acids. Plant Physiol. 2016, 171, 508–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dündar, E.; Bush, D.R. BAT1, a bidirectional amino acid transporter in Arabidopsis. Planta 2009, 229, 1047–1056. [Google Scholar] [CrossRef]
- Svennerstam, H.; Ganeteg, U.; Bellini, C.; Näsholm, T. Comprehensive Screening of Arabidopsis Mutants Suggests the Lysine Histidine Transporter 1 to Be Involved in Plant Uptake of Amino Acids. Plant Physiol. 2007, 143, 1853–1860. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Gao, Y.; Chen, Z.; Xu, G.; Zhang, X.; Li, H.; Shi, J.; Koffas, M.A.G.; Xu, Z. High-yield production of l-serine through a novel identified exporter combined with synthetic pathway in Corynebacterium glutamicum. Microb. Cell Factories 2020, 19, 115. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor 1 | Factor 2 | Response 1 | |
|---|---|---|---|
| Run | A: Time (h) | B: Concentration (μg/mL) | Lysine Content (mg/g DW) |
| 1 | 81.94 | 25.61 | 26.77 |
| 2 | 0.00 | 15.00 | 27.38 |
| 3 | 14.06 | 25.61 | 25.67 |
| 4 | 48.00 | 0.00 | 27.76 |
| 5 | 81.94 | 4.39 | 25.92 |
| 6 | 48.00 | 15.00 | 30.88 |
| 7 | 48.00 | 15.00 | 30.89 |
| 8 | 14.06 | 4.39 | 25.77 |
| 9 | 48.00 | 30.00 | 26.92 |
| 10 | 14.06 | 25.61 | 25.43 |
| 11 | 0.00 | 15.00 | 27.58 |
| 12 | 96.00 | 15.00 | 29.65 |
| 13 | 48.00 | 30.00 | 26.14 |
| 14 | 48.00 | 0.00 | 27.94 |
| 15 | 14.06 | 4.39 | 25.66 |
| 16 | 81.94 | 25.61 | 26.39 |
| 17 | 48.00 | 15.00 | 30.98 |
| 18 | 81.94 | 4.39 | 25.86 |
| 19 | 48.00 | 15.00 | 30.79 |
| 20 | 96.00 | 15.00 | 29.16 |
| 21 | 48.00 | 15.00 | 31 |
| Source | Sum of Squares | DF | Mean Squares | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|
| Model | 69.77 | 5 | 13.95 | 12.47 | <0.0001 |
| Residual | 16.78 | 15 | 1.12 | ||
| Lack of fit | 16.18 | 3 | 5.39 | 108.24 | <0.0001 |
| Pure error | 0.60 | 12 | 0.050 | ||
| Cor total | 86.55 | 20 |
| Factor | Coefficient Estimate | Standard Error | %95 CI Low | %95 CI High | F Value | p-Value (Prob > F) |
|---|---|---|---|---|---|---|
| Intercept | 30.91 | 0.46 | 29.90 | 31.92 | ||
| A-Time | 0.49 | 0.26 | −0.073 | 1.05 | 3.45 | 0.0831 |
| B-Concentration | −0.17 | 0.26 | −0.73 | 0.40 | 0.40 | 0.5354 |
| AB | 0.21 | 0.37 | −0.58 | 1.01 | 0.33 | 0.5760 |
| A2 | −1.70 | 0.32 | −2.41 | −1.00 | 26.78 | 0.0001 |
| B2 | −2.33 | 0.32 | −3.03 | −1.63 | 50.08 | <0.0001 |
| Chitosan Concentration | Average Lysine Content (mg/g) | Lysine Content Increase Ratio (%) | Average Growth Length (cm) | Growth Length Increase Ratio (%) |
|---|---|---|---|---|
| Control | 9.58 ± 0.31 c | - | 13.06 ± 0.25 c | - |
| 10 μg/mL | 10.46 ± 0.15 b | 9.19 | 14.48 ± 0.19 b | 10.87 |
| 14.61 μg/mL | 11.05 ± 0.23 a | 15.34 | 16.11 ± 0.11 a | 23.35 |
| 20 μg/mL | 9.61 ± 0.26 c | 0.31 | 12.93 ± 0.18 d | −1.00 |
| 30 μg/mL | 8.57 ± 0.29 d | −10.54 | 11.80 ± 0.16 e | −9.65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, L.; Ma, Z.; Qiu, H.; Liu, X.; Wu, T.; Ge, F.; Liu, R.; Zhu, J.; Shi, L.; Jiang, A.; et al. Chitosan Increases Lysine Content through Amino Acid Transporters in Flammulina filiformis. Foods 2022, 11, 2163. https://doi.org/10.3390/foods11142163
Tian L, Ma Z, Qiu H, Liu X, Wu T, Ge F, Liu R, Zhu J, Shi L, Jiang A, et al. Chitosan Increases Lysine Content through Amino Acid Transporters in Flammulina filiformis. Foods. 2022; 11(14):2163. https://doi.org/10.3390/foods11142163
Chicago/Turabian StyleTian, Li, Zhaodi Ma, Hao Qiu, Xiaotian Liu, Tao Wu, Feng Ge, Rui Liu, Jing Zhu, Liang Shi, Ailiang Jiang, and et al. 2022. "Chitosan Increases Lysine Content through Amino Acid Transporters in Flammulina filiformis" Foods 11, no. 14: 2163. https://doi.org/10.3390/foods11142163
APA StyleTian, L., Ma, Z., Qiu, H., Liu, X., Wu, T., Ge, F., Liu, R., Zhu, J., Shi, L., Jiang, A., Yu, H., & Ren, A. (2022). Chitosan Increases Lysine Content through Amino Acid Transporters in Flammulina filiformis. Foods, 11(14), 2163. https://doi.org/10.3390/foods11142163