
Antibiofilm and Antiquorum Sensing Potential of Lactiplantibacillus plantarum Z057 against Vibrio parahaemolyticus

 , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Strain and Culture Conditions
2.2. Co-Culture of L. plantarum Z057 and V. parahaemolyticus
2.3. Determination of the Minimum Inhibitory Concentration (MIC)
2.4. Biofilm Inhibition Assay
2.5. Measurement of the Metabolic Activity of Biofilms
2.6. Evaluation of the Physicochemical Properties of V. parahaemolyticus Treated with Z057-E
2.7. Swarming and Swimming Motilities
2.8. Autoinducer-2 (AI-2) Bioassay
2.9. Analysis of Extracellular Polymer Matrix in Biofilm
2.10. Visualization of the Biofilms by Fluorescence Microscopy
2.11. Visualization of the Biofilms Using Scanning Electron Microscopy (SEM)
2.12. Analysis of the Gene Expression Using RT-qPCR
2.13. Statistical Analysis
3. Results
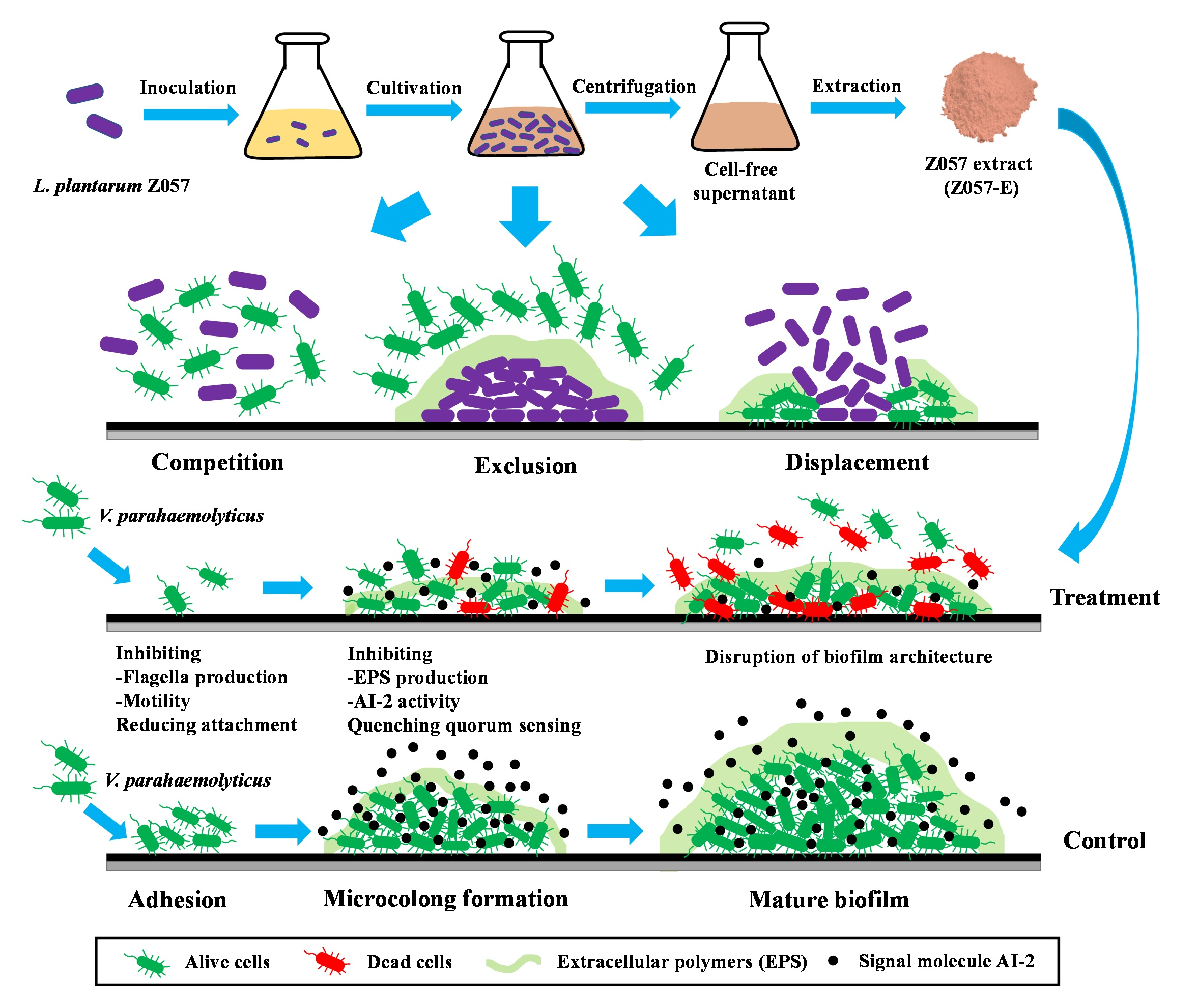
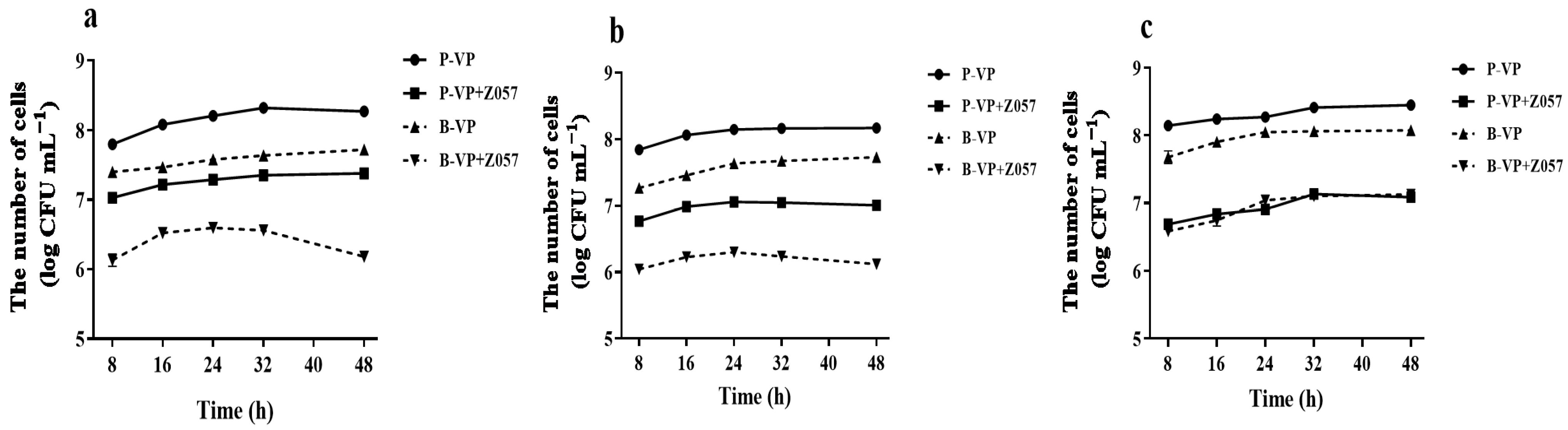
3.1. L. plantarum Z057-Mediated Inhibition on V. parahaemolyticus Biofilm
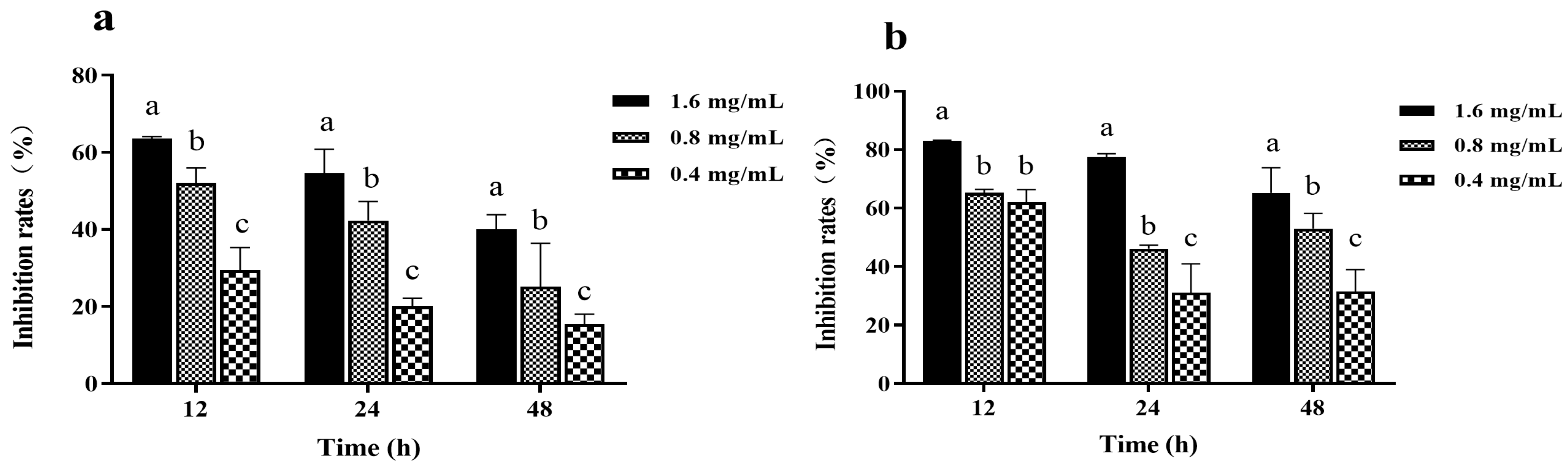
3.2. Effects of Z057-E on Biofilm Formation of V. parahaemolyticus
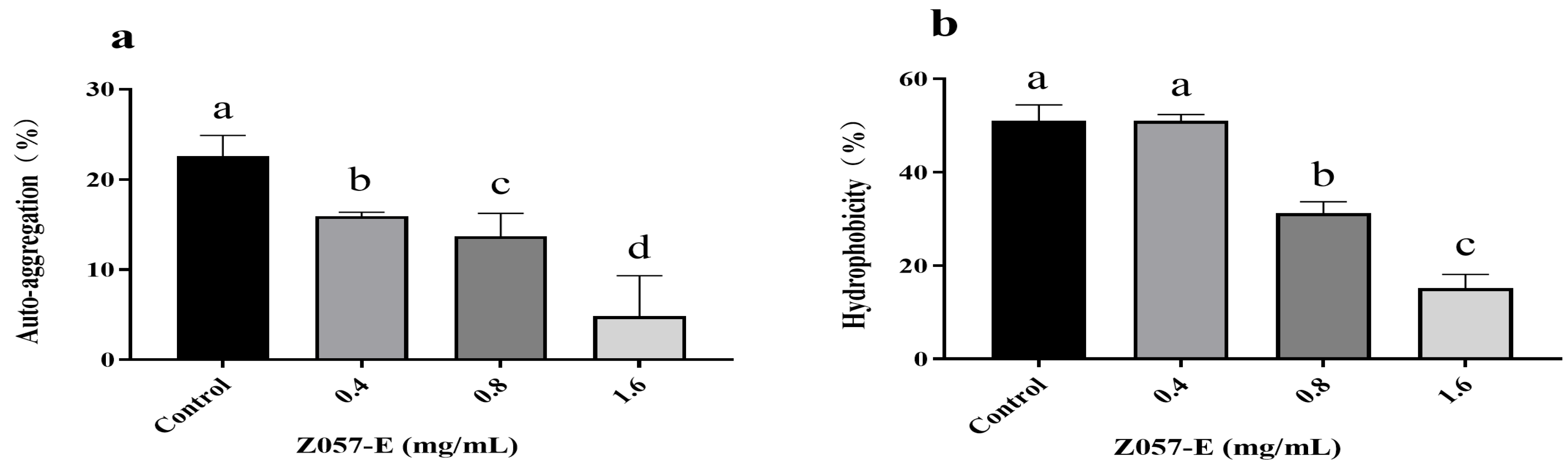
3.3. Effects of Z057-E on the Auto-Aggregation and Cell Surface Hydrophobicity of V. parahaemolyticus
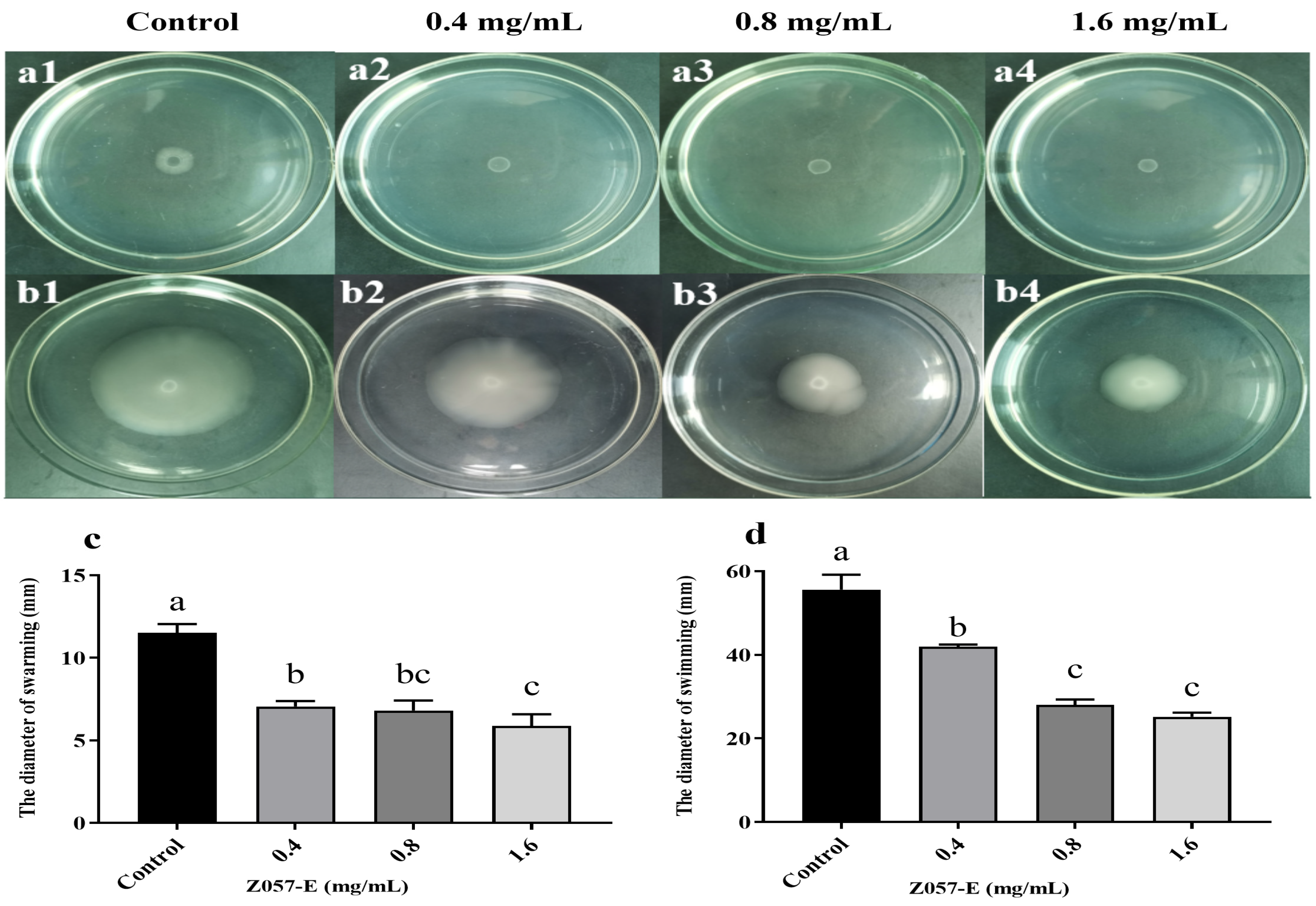
3.4. Effects of Z057-E on the Swarming and Swimming Abilities of V. parahaemolyticus
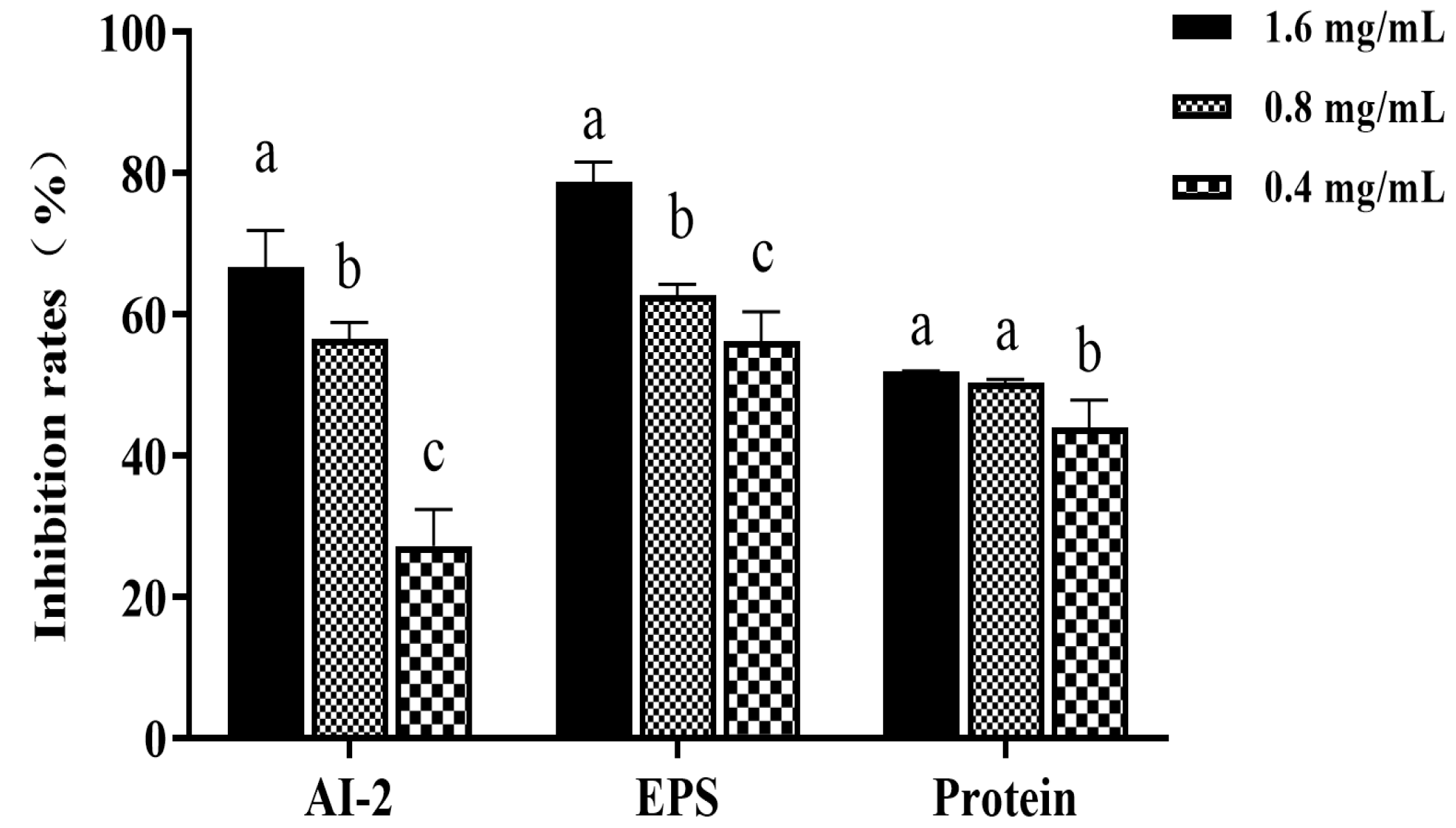
3.5. Effect of Z057-E on the Activity of Signal Molecule AI-2 of V. parahaemolyticus
3.6. Effect of Z057-E on EPS, Extracellular Protein of V. parahaemolyticus Biofilm
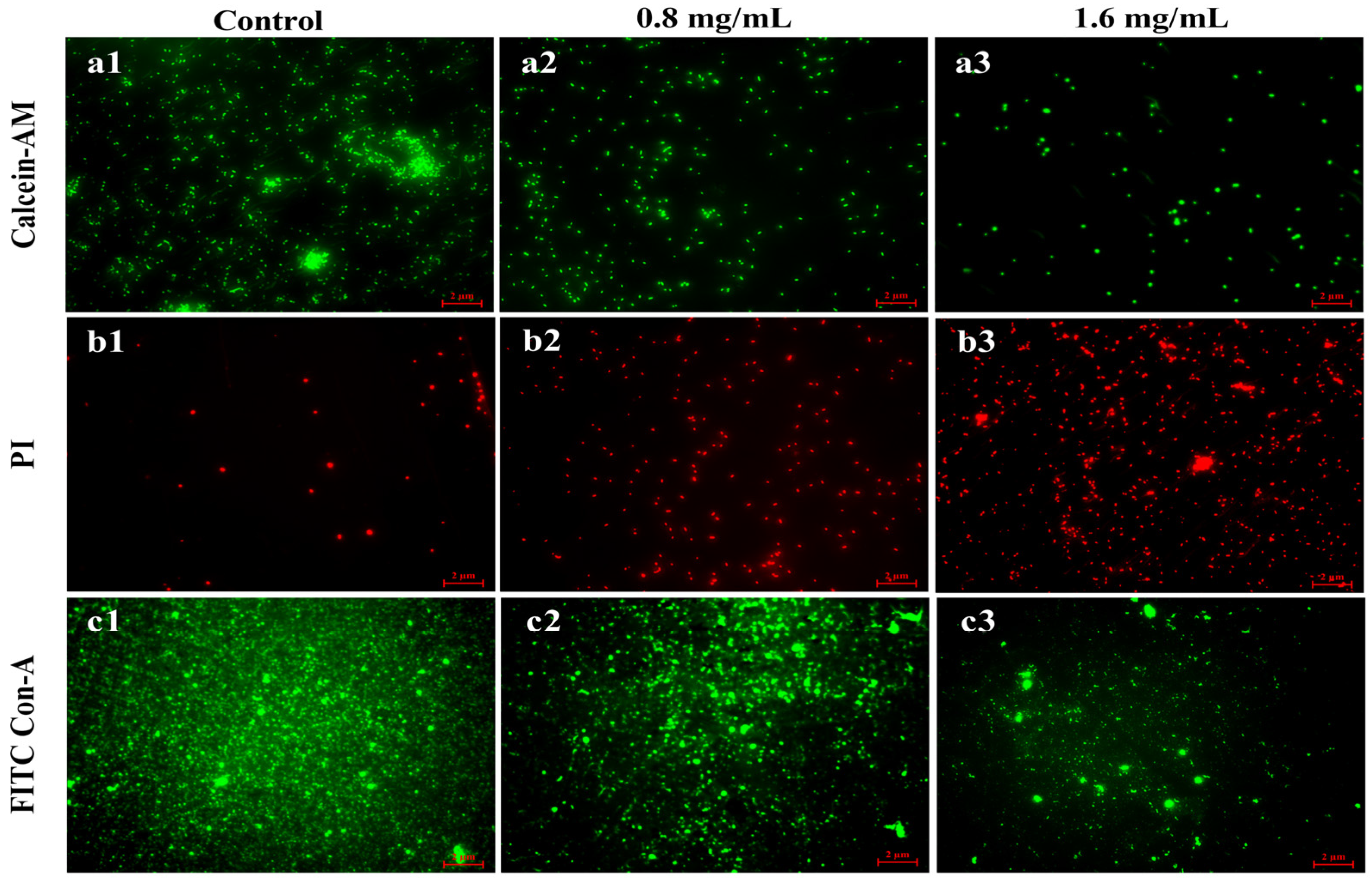
3.7. Fluorescence Microscope Visualization of Biofilm
3.8. Scanning Electron Microscope (SEM) Visualization of the Changes of Biofilm Structures
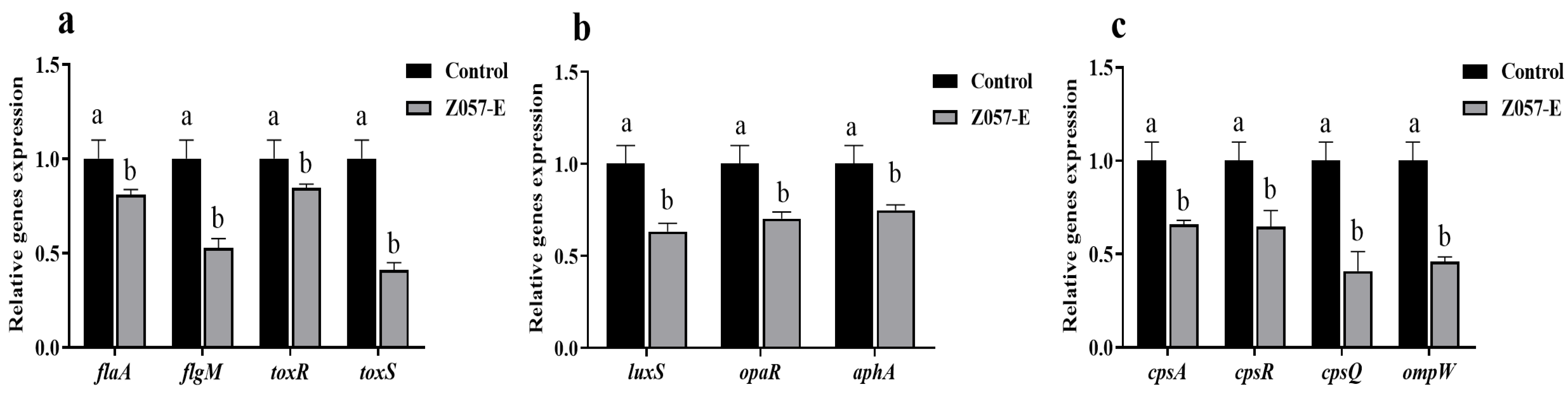
3.9. Z057-E Modulated Expressions of Biofilm-Related Genes and QS-Related Genes in V. parahaemolyticus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hubbard, T.P.; Chao, M.C.; Abel, S.; Blondel, C.J.; Wiesch, P.A.Z.; Zhou, X.; Davis, B.M.; Waldor, M.K. Genetic analysis of Vibrio parahaemolyticus intestinal colonization. Proc. Natl. Acad. Sci. USA 2016, 113, 6283–6288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sahlany, S.T.G. Effect of mentha piperita essential oil against Vibrio spp. isolated from local cheeses. Pak. J. Food Sci. 2016, 26, 65–71. [Google Scholar]
- Xie, T.; Xu, X.; Wu, Q.; Zhang, J.; Cheng, J. Prevalence, molecular characterization, and antibiotic susceptibility of Vibrio parahaemolyticus from ready-to-eat foods in China. Front. Microbiol. 2016, 7, 549–559. [Google Scholar] [CrossRef]
- Farisa Banu, S.; Rubini, D.; Murugan, R.; Vadivel, V.; Gowrishankar, S.; Pandian, S.K.; Nithyanand, P. Exploring the antivirulent and sea food preservation efficacy of essential oil combined with DNase on Vibrio parahaemolyticus. LWT 2018, 95, 107–115. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef]
- Rutherford, S.T.; van Kessel, J.C.; Shao, Y.; Bassler, B.L. AphA and LuxR/HapR reciprocally control quorum sensing in vibrios. Genes Dev. 2011, 25, 397–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, T.; Liao, Z.; Lei, H.; Fang, X.; Wang, J.; Zhong, Q. Antibacterial activity of food-grade chitosan against Vibrio parahaemolyticus biofilms. Microb. Pathog. 2017, 110, 291–297. [Google Scholar] [CrossRef]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Zhong, Y.; Gu, X.; Yuan, J.; Saeed, A.F.; Wang, S. The pathogenesis, detection, and prevention of Vibrio parahaemolyticus. Front. Microbiol. 2015, 6, 144–157. [Google Scholar] [CrossRef]
- Zhu, S.; Kojima, S.; Homma, M. Structure, gene regulation and environmental response of flagella in Vibrio. Front. Microbiol. 2013, 4, 410–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, L.N.; Shi, C.Z.; Luo, C.X.; Hu, C.Y.; Meng, Y.H. Phloretin inhibits biofilm formation by affecting quorum sensing under different temperature. LWT 2020, 131, 109668–109676. [Google Scholar] [CrossRef]
- Pang, X.; Wong, C.; Chung, H.-J.; Yuk, H.-G. Biofilm formation of Listeria monocytogenes and its resistance to quaternary ammonium compounds in a simulated salmon processing environment. Food Control 2019, 98, 200–208. [Google Scholar] [CrossRef]
- Ashrafudoulla, M.; Mizan, M.F.R.; Park, H.; Byun, K.-H.; Lee, N.; Park, S.H.; Ha, S.-D. Genetic Relationship, Virulence Factors, Drug Resistance Profile and Biofilm Formation Ability of Vibrio parahaemolyticus Isolated from Mussel. Front. Microbiol. 2019, 10, 513–527. [Google Scholar] [CrossRef]
- Zheng, X.; Guo, J.; Rao, H.; Guo, D.; Huang, Y.; Xu, Y.; Liang, S.; Xia, X.; Shi, C. Antibacterial and antibiofilm activity of coenzyme Q0 against Vibrio parahaemolyticus. Food Control 2020, 109, 106955–106963. [Google Scholar] [CrossRef]
- Danh, L.T.; Han, L.N.; Triet, N.D.A.; Zhao, J.; Mammucari, R.; Foster, N. Comparison of Chemical Composition, Antioxidant and Antimicrobial Activity of Lavender (Lavandula angustifolia L.) Essential Oils Extracted by Supercritical CO2, Hexane and Hydrodistillation. Food Bioprocess Technol. 2012, 6, 3481–3489. [Google Scholar] [CrossRef]
- Shangguan, W.; Xie, T.; Zhang, R.; Lu, C.; Han, X.; Zhong, Q. Anti-biofilm potential of kefir-derived Lactobacillus paracasei L10 against Vibrio parahaemolyticus. Lett. Appl. Microbiol. 2021, 73, 750–758. [Google Scholar] [CrossRef]
- Lu, C.; Liu, H.; Shangguan, W.; Chen, S.; Zhong, Q. Antibiofilm activities of the cinnamon extract against Vibrio parahaemolyticus and Escherichia coli. Arch. Microbiol. 2021, 203, 125–135. [Google Scholar] [CrossRef]
- Kariyawasam, K.M.G.M.M.; Lee, N.-K.; Paik, H.-D. Fermented dairy products as delivery vehicles of novel probiotic strains isolated from traditional fermented Asian foods. J. Food Sci. Technol. 2021, 58, 2467–2478. [Google Scholar] [CrossRef] [PubMed]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs from Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters during Food-Animal Production. Front. Microbiol. 2019, 10, 57–74. [Google Scholar] [CrossRef] [Green Version]
- Kumariya, R.; Garsa, A.K.; Rajput, Y.S.; Sood, S.K.; Akhtar, N.; Patel, S. Bacteriocins: Classification, synthesis, mechanism of action and resistance development in food spoilage causing bacteria. Microb. Pathog. 2019, 128, 171–177. [Google Scholar] [CrossRef]
- Wan, L.Y.M.; Chen, Z.J.; Shah, N.P.; El-Nezami, H. Modulation of Intestinal Epithelial Defense Responses by Probiotic Bacteria. Crit. Rev. Food Sci. Nutr. 2016, 56, 2628–2641. [Google Scholar] [CrossRef]
- Rasheed, H.A.; Tuoheti, T.; Zhang, Y.; Azi, F.; Tekliye, M.; Dong, M. Purification and partial characterization of a novel bacteriocin produced by bacteriocinogenic Lactobacillus fermentum BZ532 isolated from Chinese fermented cereal beverage (Bozai). LWT 2020, 124, 109113–109121. [Google Scholar] [CrossRef]
- Woo, J.; Ahn, J. Probiotic-mediated competition, exclusion and displacement in biofilm formation by food-borne pathogens. Lett. Appl. Microbiol. 2013, 56, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Ibarreche, M.; Castellano, P.; Leclercq, A.; Vignolo, G. Control of Listeria monocytogenes biofilms on industrial surfaces by the bacteriocin-producing Lactobacillus sakei CRL1862. FEMS Microbiol. Lett. 2016, 363, 118–124. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.W.; Luo, H.Z.; Jiang, H.; Jian, T.K.; Chen, Z.Q.; Jia, A.Q. Hordenine: A novel quorum sensing inhibitor and antibiofilm agent against Pseudomonas aeruginosa. J. Agric. Food Chem. 2018, 66, 1620–1628. [Google Scholar] [CrossRef]
- Bazargani, M.M.; Rohloff, J. Antibiofilm activity of essential oils and plant extracts against Staphylococcus aureus and Escherichia coli biofilms. Food Control 2016, 61, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Salaheen, S.; Nguyen, C.; Hewes, D.; Biswas, D. Cheap extraction of antibacterial compounds of berry pomace and their mode of action against the pathogen Campylobacter jejuni. Food Control 2014, 46, 174–181. [Google Scholar] [CrossRef]
- Packiavathy, I.A.S.V.; Sasikumar, P.; Pandian, S.K.; Ravi, A.V. Prevention of quorum-sensing-mediated biofilm development and virulence factors production in Vibrio spp. by curcumin. Appl. Microbiol. Biotechnol. 2013, 97, 10177–10187. [Google Scholar] [CrossRef]
- Yan, X.; Gu, S.; Cui, X.; Shi, Y.; Wen, S.; Chen, H.; Ge, J. Antimicrobial, anti-adhesive and anti-biofilm potential of biosurfactants isolated from Pediococcus acidilactici and Lactobacillus plantarum against Staphylococcus aureus CMCC26003. Microb. Pathog. 2019, 127, 12–20. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.; Abdel-Samie, M.A.; Cui, H.; Lin, L. Unraveling the inhibitory mechanism of clove essential oil against Listeria monocytogenes biofilm and applying it to vegetable surfaces. LWT 2020, 134, 110210–110219. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, D.; Hua, Z.; Sun, H.; Zheng, Z.; Xia, X.; Shi, C. Attenuation of Multiple Vibrio parahaemolyticus Virulence Factors by Citral. Front. Microbiol. 2019, 10, 894–904. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Tan, L.; Guo, L.; Zhang, P.; Malakar, P.K.; Ahmed, F.; Liu, H.; Wang, J.J.; Zhao, Y. Acidic electrolyzed water more effectively breaks down mature Vibrio parahaemolyticus biofilm than DNase I. Food Control 2020, 117, 107312–107321. [Google Scholar] [CrossRef]
- Chen, B.; Huang, J.; Li, H.; Zeng, Q.-H.; Wang, J.J.; Liu, H.; Pan, Y.; Zhao, Y. Eradication of planktonic Vibrio parahaemolyticus and its sessile biofilm by curcumin-mediated photodynamic inactivation. Food Control 2020, 113, 107181–107191. [Google Scholar] [CrossRef]
- Merino, L.; Trejo, F.M.; De Antoni, G.; Golowczyc, M.A. Lactobacillus strains inhibit biofilm formation of Salmonella sp. isolates from poultry. Food Res. Int. 2019, 123, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Bai, F.; Sun, M.; Lv, X.; Li, X.; Zhang, D.; Du, H. Lactobacillus crustorum ZHG 2-1 as novel quorum-quenching bacteria reducing virulence factors and biofilms formation of Pseudomonas aeruginosa. LWT 2020, 117, 108696–108704. [Google Scholar] [CrossRef]
- Winkelstroter, L.K.; Gomes, B.C.; Thomaz, M.R.S.; Souza, V.M.; De Martinis, E.C.P. Lactobacillus sakei 1 and its bacteriocin influence adhesion of Listeria monocytogenes on stainless steel surface. Food Control 2011, 22, 1404–1407. [Google Scholar] [CrossRef]
- Rendueles, O.; Ghigo, J.-M. Multi-species biofilms: How to avoid unfriendly neighbors. FEMS Microbiol. Rev. 2012, 36, 972–989. [Google Scholar] [CrossRef]
- Hammami, I.; Rhouma, A.; Jaouadi, B.; Rebai, A.; Nesme, X. Optimization and biochemical characterization of a bacteriocin from a newly isolated Bacillus subtilis strain 14B for biocontrol of Agrobacterium spp. strains. Lett. Appl. Microbiol. 2009, 48, 253–260. [Google Scholar] [CrossRef]
- Ibarreche, M.P.; Castellano, P.; Vignolo, G. Evaluation of anti-Listeria meat borne Lactobacillus for biofilm formation on selected abiotic surfaces. Meat Sci. 2014, 96, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Jeniel, E.N.; Sanchez, H.; Michael, T.C.; David, R.A. Genetic basis of Candida biofilm resistance due to drug sequestering matrix glucan. J. Infect. Dis. 2010, 202, 171–175. [Google Scholar]
- Shen, Y.; Huang, C.; Monroy, G.L.; Janjaroen, D.; Derlon, N.; Lin, J.; Espinosa-Marzal, R.; Morgenroth, E.; Boppart, S.A.; Ashbolt, N.J.; et al. Response of Simulated Drinking Water Biofilm Mechanical and Structural Properties to Long-Term Disinfectant Exposure. Environ. Sci. Technol. 2016, 50, 1779–1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onbas, T.; Osmanagaoglu, O.; Kiran, F. Potential Properties of Lactobacillus plantarum F-10 as a Bio-control Strategy for Wound Infections. Probiotics Antimicrob. Proteins 2019, 11, 1110–1123. [Google Scholar] [CrossRef]
- Wang, N.; Yuan, L.; Sadiq, F.A.; He, G. Inhibitory effect of Lactobacillus plantarum metabolites against biofilm formation by Bacillus licheniformis isolated from milk powder products. Food Control 2019, 106, 106721–106732. [Google Scholar] [CrossRef]
- Kanmani, P.; Satish kumar, R.; Yuvaraj, N.; Paari, K.A.; Pattukumar, V.; Arul, V. Production and purification of a novel exopolysaccharide from lactic acid bacterium Streptococcus phocae PI80 and its functional characteristics activity in vitro. Bioresour. Technol. 2011, 102, 4827–4833. [Google Scholar] [CrossRef] [PubMed]
- Rana, S.; Bhawal, S.; Kumari, A.; Kapila, S.; Kapila, R. pH-dependent inhibition of AHL-mediated quorum sensing by cell-free supernatant of lactic acid bacteria in Pseudomonas aeruginosa PAO1. Microb. Pathog. 2020, 142, 104105–104113. [Google Scholar] [CrossRef] [PubMed]
- Ritter, A.; Com, E.; Bazire, A.; Goncalves, M.D.S.; Delage, L.; Le Pennec, G.; Pineau, C.; Dreanno, C.; Compere, C.; Dufour, A. Proteomic studies highlight outer-membrane proteins related to biofilm development in the marine bacterium Pseudoalteromonas sp. D41. Proteomics 2012, 12, 3180–3192. [Google Scholar] [CrossRef]
- Chen, Y.-C.; Ho, H.-O.; Su, C.-H.; Sheu, M.-T. Anticancer Effects of Taiwanofungus camphoratus Extracts, Isolated Compounds and its Combinational use. J. Exp. Clin. Med. 2010, 2, 274–281. [Google Scholar] [CrossRef]
- Wang, F.; He, Q.; Yin, J.; Xu, S.; Hu, W.; Gu, L. BrlR from Pseudomonas aeruginosa is a receptor for both cyclic di-GMP and pyocyanin. Nat. Commun. 2018, 9, 2563–2577. [Google Scholar] [CrossRef]
- Guvener, Z.T.; McCarter, L.L. Multiple Regulators Control Capsular Polysaccharide Production in Vibrio parahaemolyticus. J. Bacteriol. 2003, 185, 5431–5441. [Google Scholar] [CrossRef] [Green Version]
- Aschtgen, M.-S.; Brennan, C.A.; Nikolakakis, K.; Cohen, S.; McFall-Ngai, M.; Ruby, E.G. Insights into flagellar function and mechanism from the squid–vibrio symbiosis. NPJ Biofilms Microbiomes 2019, 5, 32–42. [Google Scholar] [CrossRef]
- Loh, J.T.; Forsyth, M.H.; Cover, T.L. Growth phase regulation of flaA expression in Helicobacter pylori is luxS dependent. Infect. Immun. 2004, 72, 5506–5510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, D.; Yang, Z.; Zheng, X.; Kang, S.; Yang, Z.; Xu, Y.; Shi, C.; Tian, H.; Xia, X. Thymoquinone Inhibits Biofilm Formation and Attachment-Invasion in Host Cells of Vibrio parahaemolyticus. Foodborne Pathog. Dis. 2019, 16, 671–678. [Google Scholar] [CrossRef] [PubMed]
- Wadhams, G.H.; Armitage, J.P. Making sense of it all: Bacterial chemotaxis. Nat. Rev. Mol. Cell Biol. 2004, 5, 1024–1037. [Google Scholar] [CrossRef]
- Zhang, L.; Weng, Y.; Wu, Y.; Wang, X.; Yin, Z.; Yang, H.; Yang, W.; Zhang, Y. H-NS is an activator of exopolysaccharide biosynthesis genes transcription in Vibrio parahaemolyticus. Microb. Pathog. 2018, 116, 164–167. [Google Scholar] [CrossRef]
- Zhang, L.; Orth, K. Virulence determinants for Vibrio parahaemolyticus infection. Curr. Opin. Microbiol. 2013, 16, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Ashrafudoulla, M.; Mizan, M.F.R.; Ha, A.J.-W.; Park, S.H.; Ha, S.-D. Antibacterial and antibiofilm mechanism of eugenol against antibiotic resistance Vibrio parahaemolyticus. Food Microbiol. 2020, 91, 103500–103513. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′–3′) | References |
|---|---|---|
| 16S rRNA | F-GCCTTCGGGAACTCTGAGACAG R-GCTCGTTGCGGGACTTAACCCAA | This study |
| flaA | F-CGGACTAAACCGTATCGCTGAAA R-GGCTGCCCATAGAAAGCATTACA | [32] |
| flgM | F-ATGCGAATTCCATGGCAGGTATAGATAATATA R-ATGCCTCGAGGCTTTTGCCTTGCAATTCGTT | [32] |
| luxS | F-GGATTTTGTTCTGGCTTTCCACTT R-GGGATGTCGCACTGGTTTTTAC | [32] |
| opaR | F-TGTCTACCAACCGCACTAACC R-GCTCTTTCAACTCGGCTTCAC | [32] |
| aphA | F-ACACCCAACCGTTCGTGATG R-GTTGAAGGCGTTGCGTAGTAAG | [33] |
| cpsA | F-GAGAGCGGCAACCTATATCG R-GCGGTCAAACAAAGGGTAAAC | [33] |
| cpsR | F-TTGGAGTCGCACTCTGGTCAA R-TGCACGCGACACACCAAGTT | [33] |
| cpsQ | F-GCCTGAAATCCTAATGCTC R-AGTGTCAGAAGGTGTATCAAC | [33] |
| ompW | F-TCGTGTCACCAAGTGTTTTCG R-CGTGGCTGAATGGTGTTGC | [32] |
| toxR | F-CAACGAAAGCCGTATACTCCTG R-CTCAAAACCTTGCTCACGCC | [34] |
| toxS | F-ATTTTTCTGAAGCGCAACTACG R-CCGTAGAACCGTGATTTAGGCT | [34] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, X.; Chen, Q.; Zhang, X.; Chen, X.; Luo, D.; Zhong, Q. Antibiofilm and Antiquorum Sensing Potential of Lactiplantibacillus plantarum Z057 against Vibrio parahaemolyticus. Foods 2022, 11, 2230. https://doi.org/10.3390/foods11152230
Han X, Chen Q, Zhang X, Chen X, Luo D, Zhong Q. Antibiofilm and Antiquorum Sensing Potential of Lactiplantibacillus plantarum Z057 against Vibrio parahaemolyticus. Foods. 2022; 11(15):2230. https://doi.org/10.3390/foods11152230
Chicago/Turabian StyleHan, Xiangpeng, Qingying Chen, Xingguo Zhang, Xiaolan Chen, Dongsheng Luo, and Qingping Zhong. 2022. "Antibiofilm and Antiquorum Sensing Potential of Lactiplantibacillus plantarum Z057 against Vibrio parahaemolyticus" Foods 11, no. 15: 2230. https://doi.org/10.3390/foods11152230
APA StyleHan, X., Chen, Q., Zhang, X., Chen, X., Luo, D., & Zhong, Q. (2022). Antibiofilm and Antiquorum Sensing Potential of Lactiplantibacillus plantarum Z057 against Vibrio parahaemolyticus. Foods, 11(15), 2230. https://doi.org/10.3390/foods11152230