
Transcriptional Profiling Reveals Adaptive Response and Tolerance to Lactic Acid Stress in Pichia kudriavzevii

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains, Media and Culture Conditions
2.2. Lactic Acid Tolerance Tests
2.3. Biomass Detection
2.4. Analysis Methods
2.5. RNA Extraction and Sequencing
2.6. Statistical Analysis
3. Results
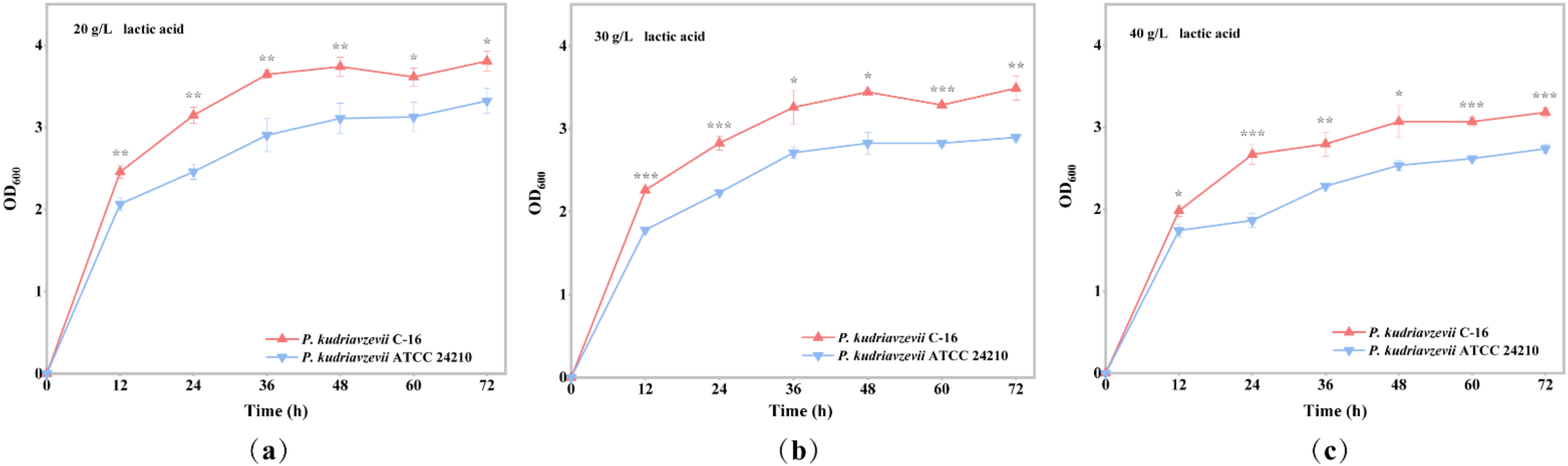
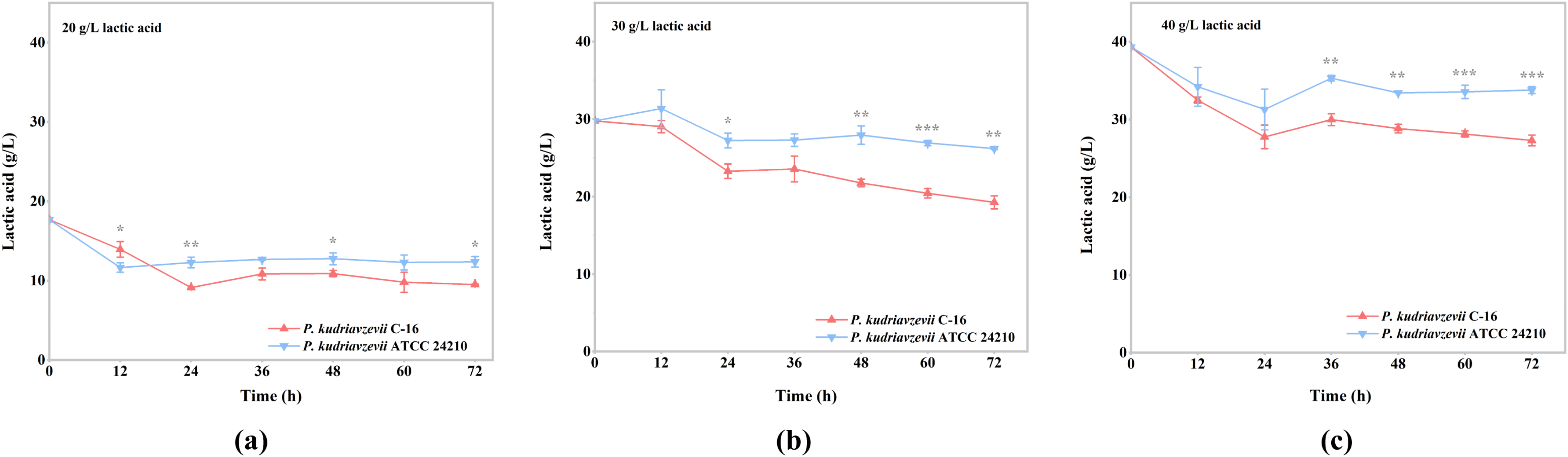
3.1. Comparison of Lactic Acid Tolerance between P. kudriavzevii C-16 and the Type Strain P. kudriavzevii ATCC 24210
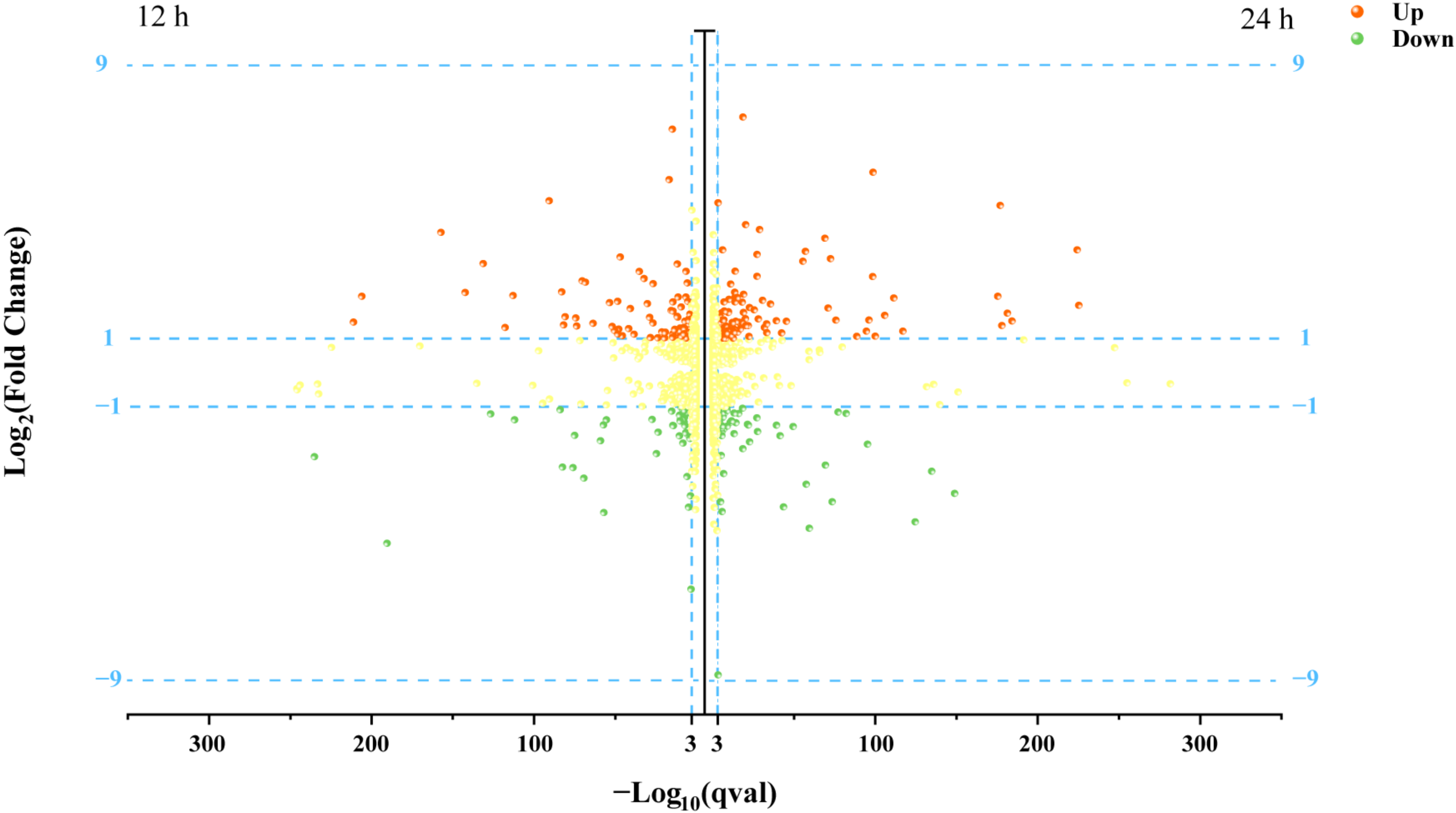
3.2. Transcriptional Profile of P. kudriavzevii C-16 under Lactic Acid Stress
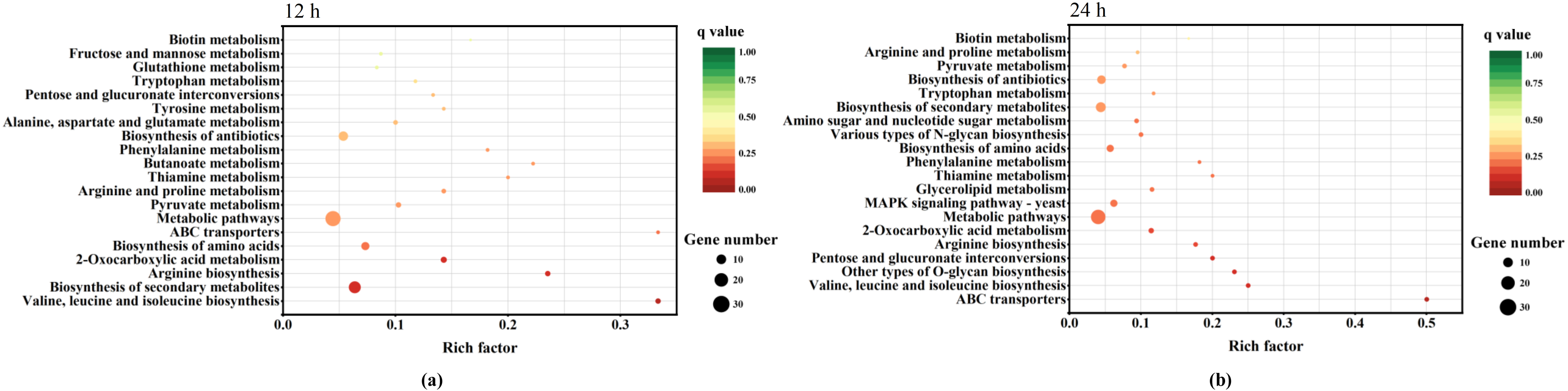
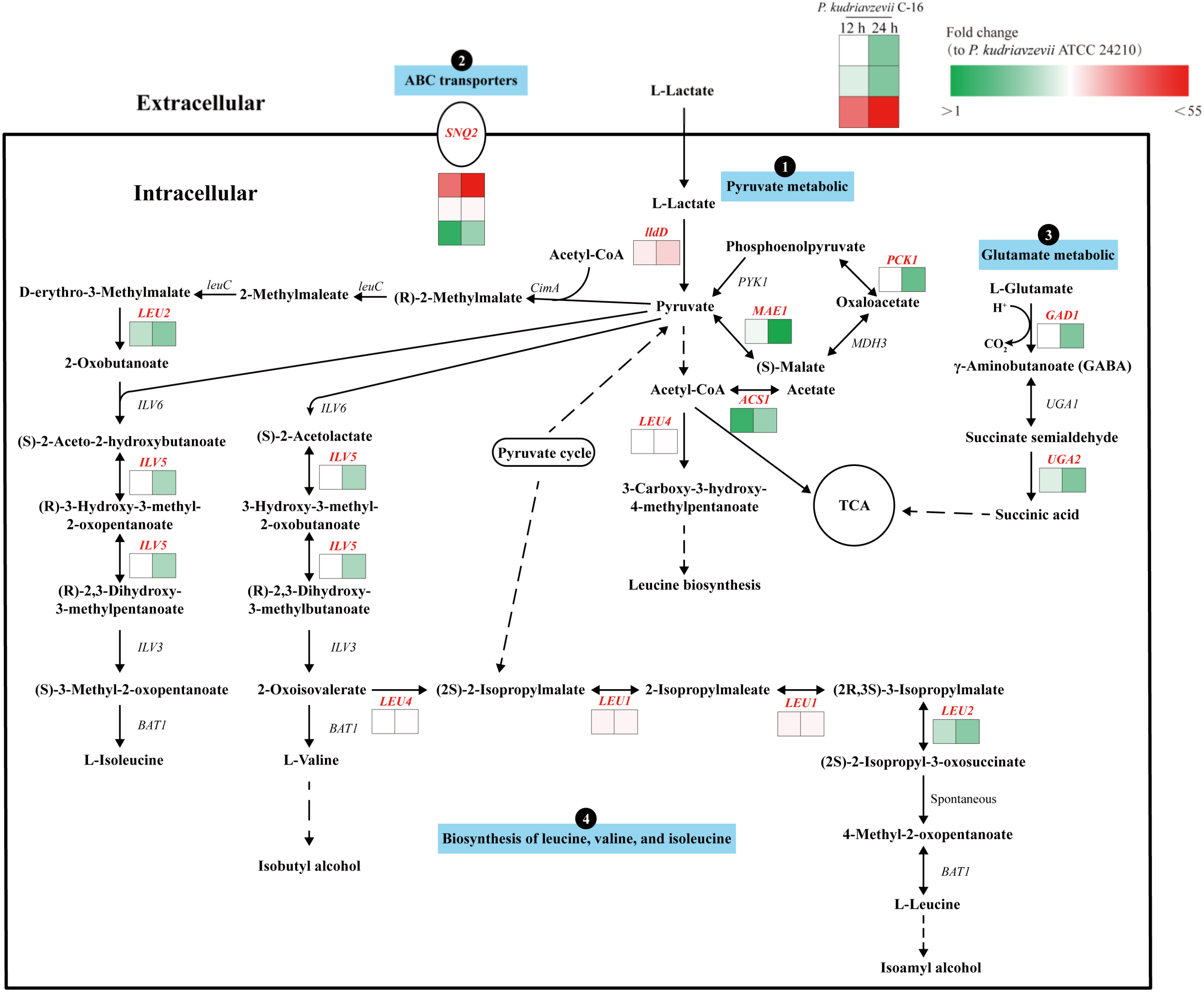
3.3. Response Mechanisms to Lactic Acid Stress in P. kudriavzevii C-16
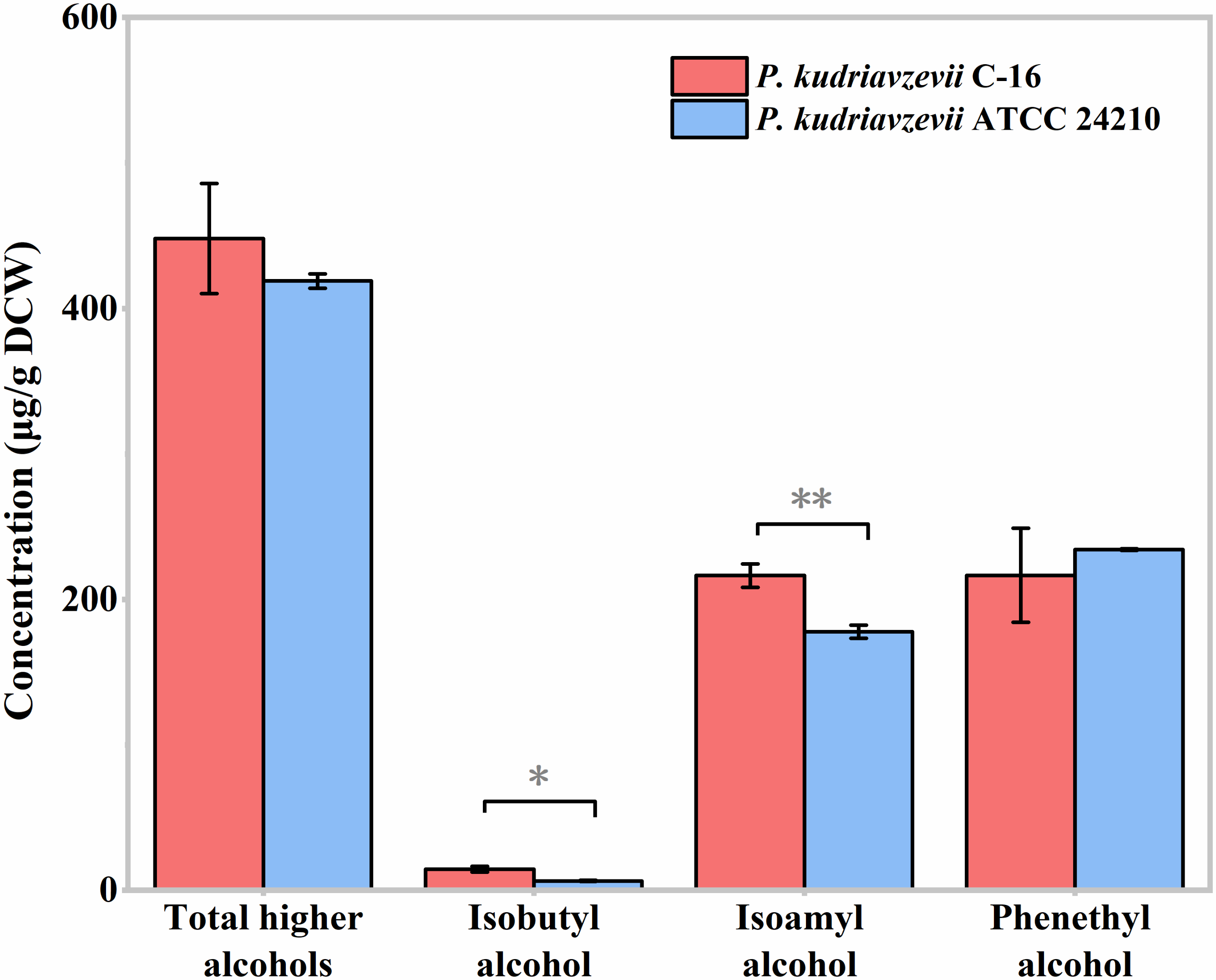
3.4. Higher Alcohols Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef]
- Gallone, B.; Mertens, S.; Gordon, J.L.; Maere, S.; Verstrepen, K.J.; Steensels, J. Origins, evolution, domestication and diversity of Saccharomyces beer yeasts. Curr. Opin. Biotechnol. 2018, 49, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Steensels, J.; Gallone, B.; Voordeckers, K.; Verstrepen, K.J. Domestication of industrial microbes. Curr. Biol. 2019, 29, R381–R393. [Google Scholar] [CrossRef]
- Liu, H.; Sun, B. Effect of Fermentation Processing on the Flavor of Baijiu. J. Agric. Food Chem. 2018, 66, 5425–5432. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Liu, Y.; Shu, L.; He, Y. Study on metabolites of Bacillus producing soy sauce-like aroma in Jiang-flavor Chinese spirits. Food Sci. Nutr. 2020, 8, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Hai, D.; Yan, Z.; Yan, X. Unraveling core functional microbiota in traditional solid-state fermentation by high-throughput amplicons and metatranscriptomics sequencing. Front. Microbiol. 2017, 8, 1294. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, H.; Starrenburg, M.J.; Molenaar, D.; Kleerebezem, M.; van Hylckama Vlieg, J.E. Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution. Genome Res. 2012, 22, 115–124. [Google Scholar] [CrossRef]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.-I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome sequencing and analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef]
- Deng, N.; Du, H.; Xu, Y. Cooperative response of Pichia kudriavzevii and Saccharomyces cerevisiae to lactic acid stress in Baijiu fermentation. J. Agric. Food Chem. 2020, 68, 4903–4911. [Google Scholar] [CrossRef]
- Gao, W.; Fan, W.; Xu, Y. Characterization of the key odorants in light aroma type Chinese liquor by gas chromatography–olfactometry, quantitative measurements, aroma recombination, and omission studies. J. Agric. Food Chem. 2014, 62, 5796–5804. [Google Scholar] [CrossRef]
- Lyu, C.; Zhao, W.; Peng, C.; Hu, S.; Fang, H.; Hua, Y.; Yao, S.; Huang, J.; Mei, L. Exploring the contributions of two glutamate decarboxylase isozymes in Lactobacillus brevis to acid resistance and γ-aminobutyric acid production. Microb. Cell Fact. 2018, 17, 180. [Google Scholar] [CrossRef]
- Liu, Y.; Tang, H.; Lin, Z.; Xu, P. Mechanisms of acid tolerance in bacteria and prospects in biotechnology and bioremediation. Biotechnol. Adv. 2015, 33, 1484–1492. [Google Scholar] [CrossRef]
- Usami, Y.; Uemura, S.; Mochizuki, T.; Morita, A.; Shishido, F.; Inokuchi, J.-i.; Abe, F. Functional mapping and implications of substrate specificity of the yeast high-affinity leucine permease Bap2. Biochim. Biophys. Acta Biomembr. 2014, 1838, 1719–1729. [Google Scholar] [CrossRef]
- Pérez-Ortín, J.E.; Querol, A.; Puig, S.; Barrio, E. Molecular characterization of a chromosomal rearrangement involved in the adaptive evolution of yeast strains. Genome Res. 2002, 12, 1533–1539. [Google Scholar] [CrossRef]
- Yuasa, N.; Nakagawa, Y.; Hayakawa, M.; Iimura, Y. Distribution of the sulfite resistance gene SSU1-R and the variation in its promoter region in wine yeasts. J. Biosci. Bioeng. 2004, 98, 394–397. [Google Scholar] [CrossRef]
- Sicard, D.; Legras, J.-L. Bread, beer and wine: Yeast domestication in the Saccharomyces sensu stricto complex. C. R. Biol. 2011, 334, 229–236. [Google Scholar] [CrossRef]
- Leng, W.; Gao, R.; Wu, X.; Zhou, J.; Sun, Q.; Yuan, L. Genome sequencing of cold-adapted Planococcus bacterium isolated from traditional shrimp paste and protease identification. J. Sci. Food Agric. 2021, 101, 3225–3236. [Google Scholar] [CrossRef]
- Legras, J.L.; Galeote, V.; Bigey, F.; Camarasa, C.; Marsit, S.; Nidelet, T.; Sanchez, I.; Couloux, A.; Guy, J.; Franco-Duarte, R.; et al. Adaptation of S. cerevisiae to fermented food environments reveals remarkable genome plasticity and the footprints of domestication. Mol. Biol. Evol. 2018, 35, 1712–1727. [Google Scholar] [CrossRef]
- Stansen, C.; Uy, D.; Delaunay, S.; Eggeling, L.; Goergen, J.L.; Wendisch, V.F. Characterization of a Corynebacterium glutamicum lactate utilization operon induced during temperature-triggered glutamate production. Appl. Environ. Microbiol. 2005, 71, 5920–5928. [Google Scholar] [CrossRef]
- Gao, C.; Jiang, T.; Dou, P.; Ma, C.; Li, L.; Kong, J.; Xu, P. NAD-independent L-lactate dehydrogenase is required for L-lactate utilization in Pseudomonas stutzeri SDM. PLoS ONE 2012, 7, e36519. [Google Scholar] [CrossRef] [Green Version]
- Guiard, B.; Lederer, F.; Jacq, C. More similarity between bakers’ yeast L-(+)-lactate dehydrogenase and liver microsomal cytochrome b5. Nature 1975, 255, 422–423. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Gu, H.; Gao, Q.; Liu, Z.L.; Bao, J. Transcriptome analysis of Zymomonas mobilis ZM4 reveals mechanisms of tolerance and detoxification of phenolic aldehyde inhibitors from lignocellulose pretreatment. Biotechnol. Biofuels 2015, 8, 153. [Google Scholar] [CrossRef] [PubMed]
- Casal, M.; Queirós, O.; Talaia, G.; Ribas, D.; Paiva, S. Carboxylic acids plasma membrane transporters in Saccharomyces cerevisiae. In Yeast Membrane Transport; Ramos, J., Sychrová, H., Kschischo, M., Eds.; Springer: Cham, Switzerland, 2016; Volume 892, pp. 229–251. [Google Scholar]
- Balzi, E.; Goffeau, A. Yeast multidrug resistance: The PDR network. J. Bioenerg. Biomembr. 1995, 27, 71–76. [Google Scholar] [CrossRef]
- Rogers, B.; Decottignies, A.; Kolaczkowski, M.; Carvajal, E.; Balzi, E.; Goffeau, A. The pleitropic drug ABC transporters from Saccharomyces cerevisiae. J. Mol. Microbiol. Biotechnol. 2001, 3, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Hirata, D.; Yano, K.; Miyahara, K.; Miyakawa, T. Saccharomyces cerevisiae YDR1, which encodes a member of the ATP-binding cassette (ABC) superfamily, is required for multidrug resistance. Curr. Genet. 1994, 26, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Feehily, C.; Karatzas, K.A.G. Role of glutamate metabolism in bacterial responses towards acid and other stresses. J. Appl. Microbiol. 2013, 114, 11–24. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W. Escherichia coli acid resistance: Tales of an amateur acidophile. Nat. Rev. Microbiol. 2004, 2, 898–907. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bown, A.W.; McLean, M.D. Metabolism and functions of gamma-aminobutyric acid. Trends Plant Sci. 1999, 4, 446–452. [Google Scholar] [CrossRef]
- Fernández, M.; Zúñiga, M. Amino acid catabolic pathways of lactic acid bacteria. Crit. Rev. Microbiol. 2006, 32, 155–183. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, H.; Fu, Y.; Deng, N.; Xu, Y. Transcriptional Profiling Reveals Adaptive Response and Tolerance to Lactic Acid Stress in Pichia kudriavzevii. Foods 2022, 11, 2725. https://doi.org/10.3390/foods11182725
Du H, Fu Y, Deng N, Xu Y. Transcriptional Profiling Reveals Adaptive Response and Tolerance to Lactic Acid Stress in Pichia kudriavzevii. Foods. 2022; 11(18):2725. https://doi.org/10.3390/foods11182725
Chicago/Turabian StyleDu, Hai, Yan Fu, Nan Deng, and Yan Xu. 2022. "Transcriptional Profiling Reveals Adaptive Response and Tolerance to Lactic Acid Stress in Pichia kudriavzevii" Foods 11, no. 18: 2725. https://doi.org/10.3390/foods11182725
APA StyleDu, H., Fu, Y., Deng, N., & Xu, Y. (2022). Transcriptional Profiling Reveals Adaptive Response and Tolerance to Lactic Acid Stress in Pichia kudriavzevii. Foods, 11(18), 2725. https://doi.org/10.3390/foods11182725