
Nitric Oxide Made a Major Contribution to the Improvement of Quality in Button Mushrooms (Agaricus bisporus) by the Combined Treatment of Nitric Oxide with 1-MCP


Abstract
:
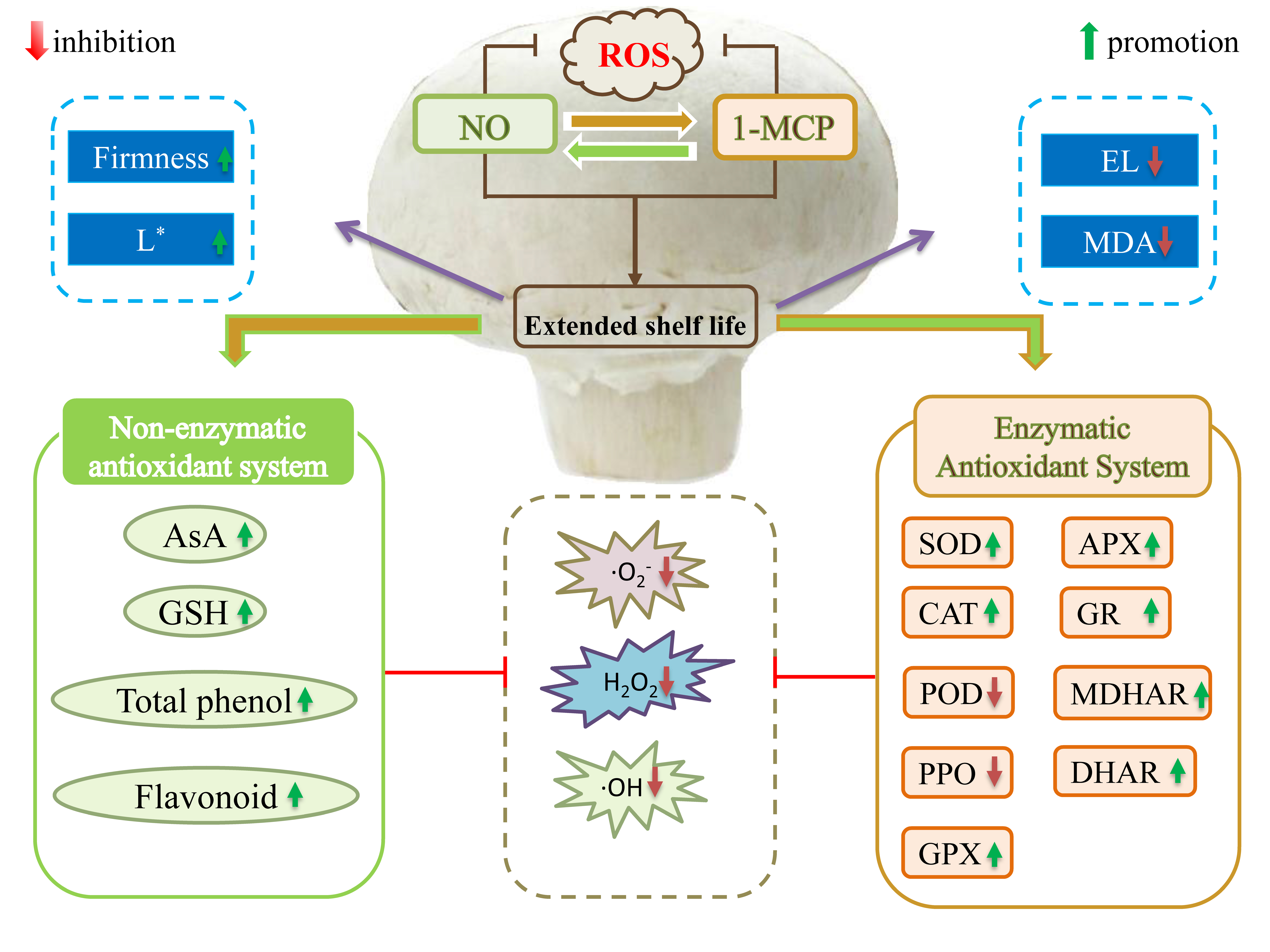{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatment
2.2. Firmness and Color Measurements
2.3. Electrolyte Leakage (EL) Measurement
2.4. Malondialdehyde (MDA) Content Measurement
2.5. Superoxide (O2·−), Hydrogen Peroxide (H2O2), and Hydroxyl Radical (·OH) Content Measurements
2.6. Enzyme Extraction and Assays
2.7. Total Phenolic and Total Flavonoid Contents Measurements
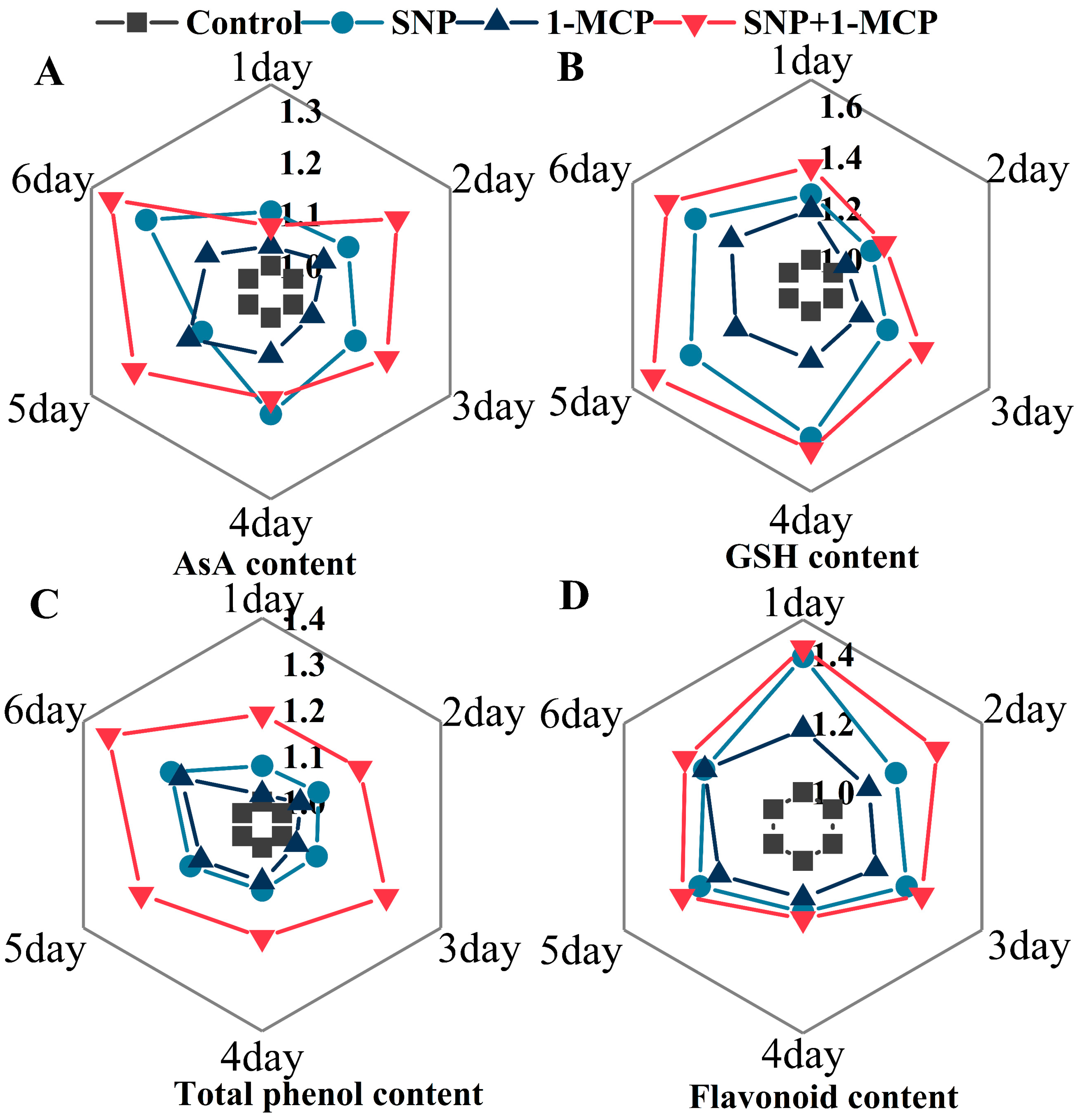
2.8. The Ascorbic Acid (AsA) and Glutathione (GSH) Contents Measurements
2.9. Statistical Analysis
3. Results
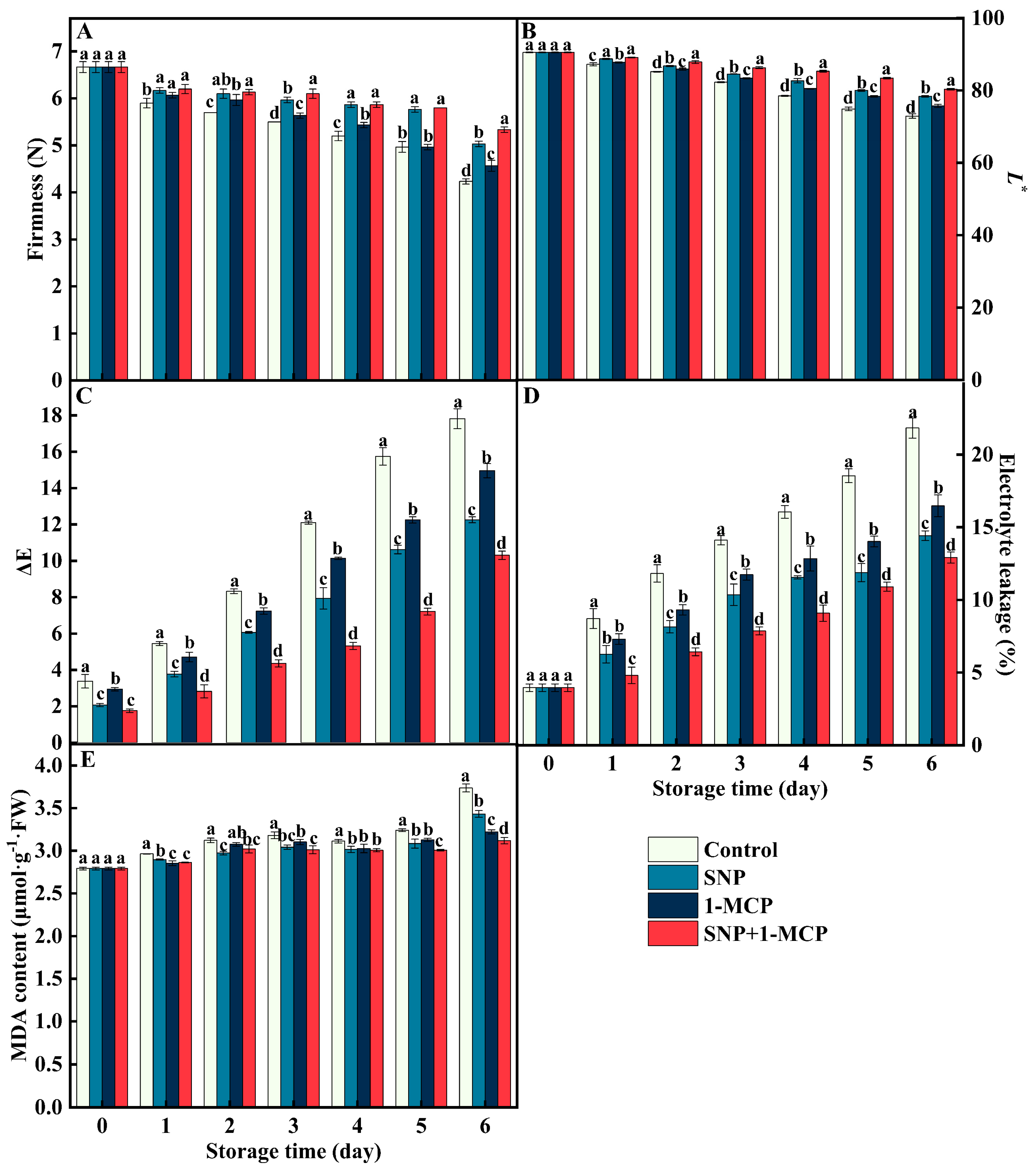
3.1. Firmness, Lightness(L*), EL, and MDA Content
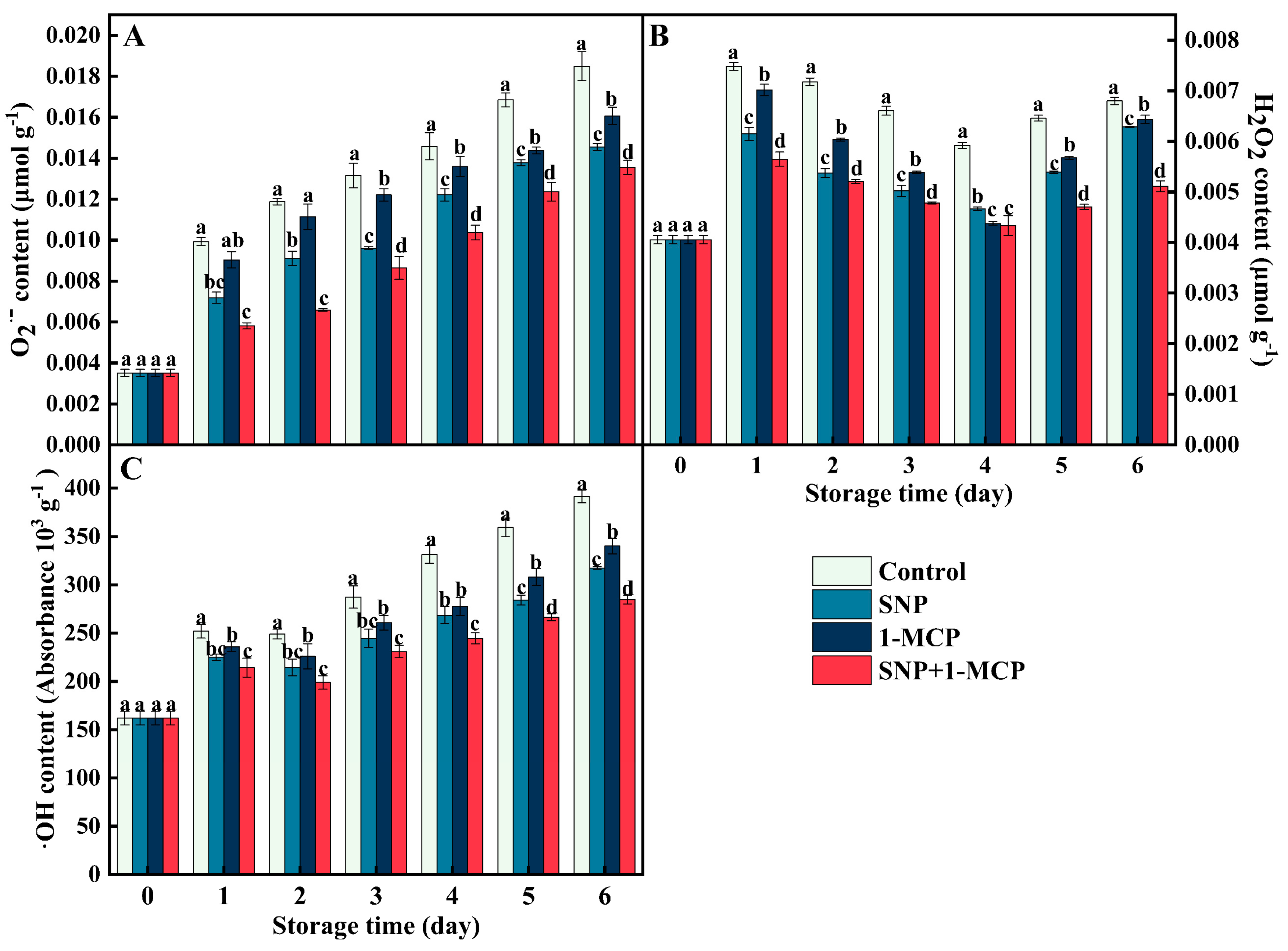
3.2. Contents of O2·−, H2O2, and ·OH
3.3. Activities of SOD, CAT, POD, and PPO
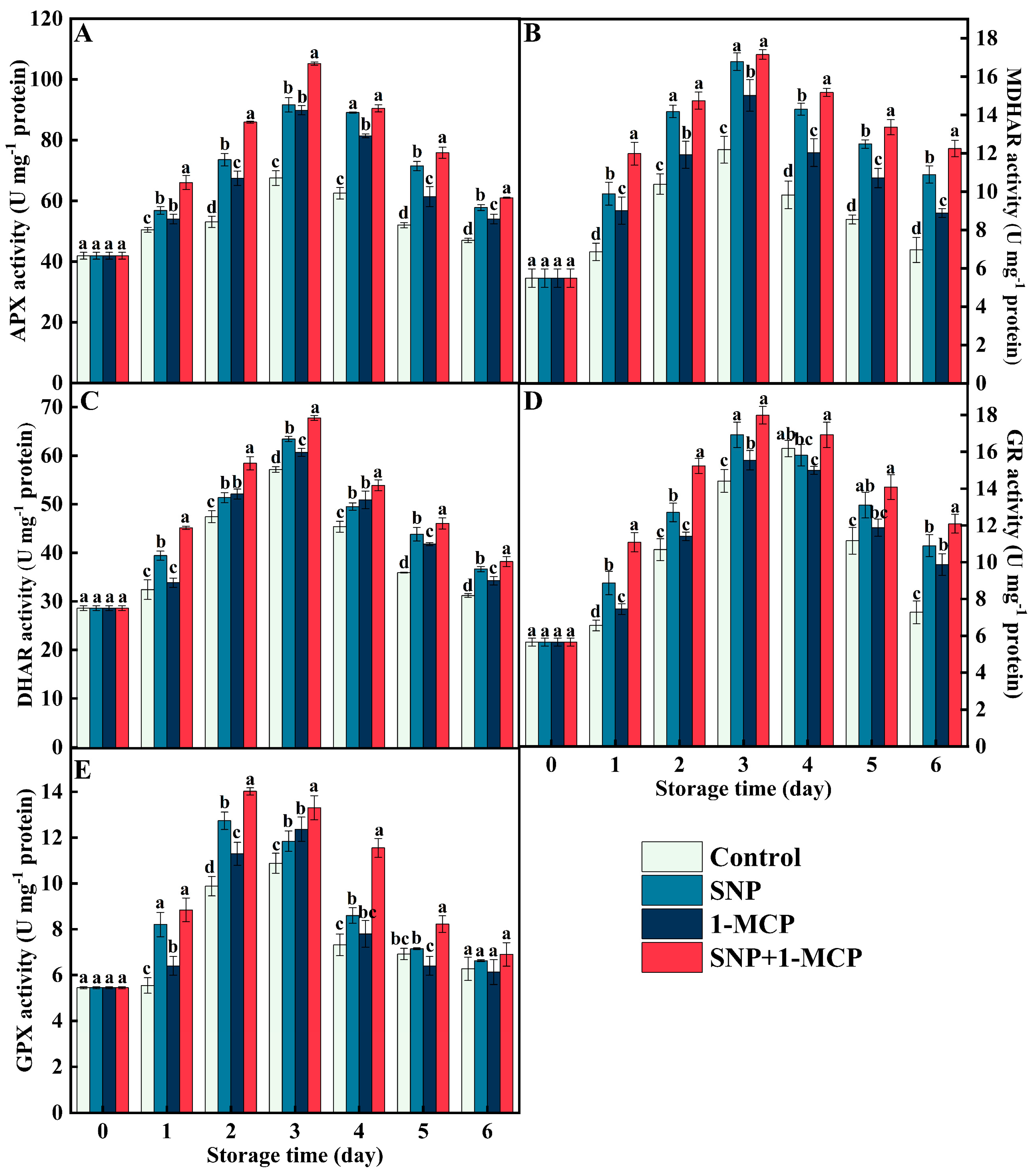
3.4. The Ascorbate–Glutathione Cycle (AsA-GSH)-Related Enzyme Activity
3.5. Contents of Antioxidant Substances
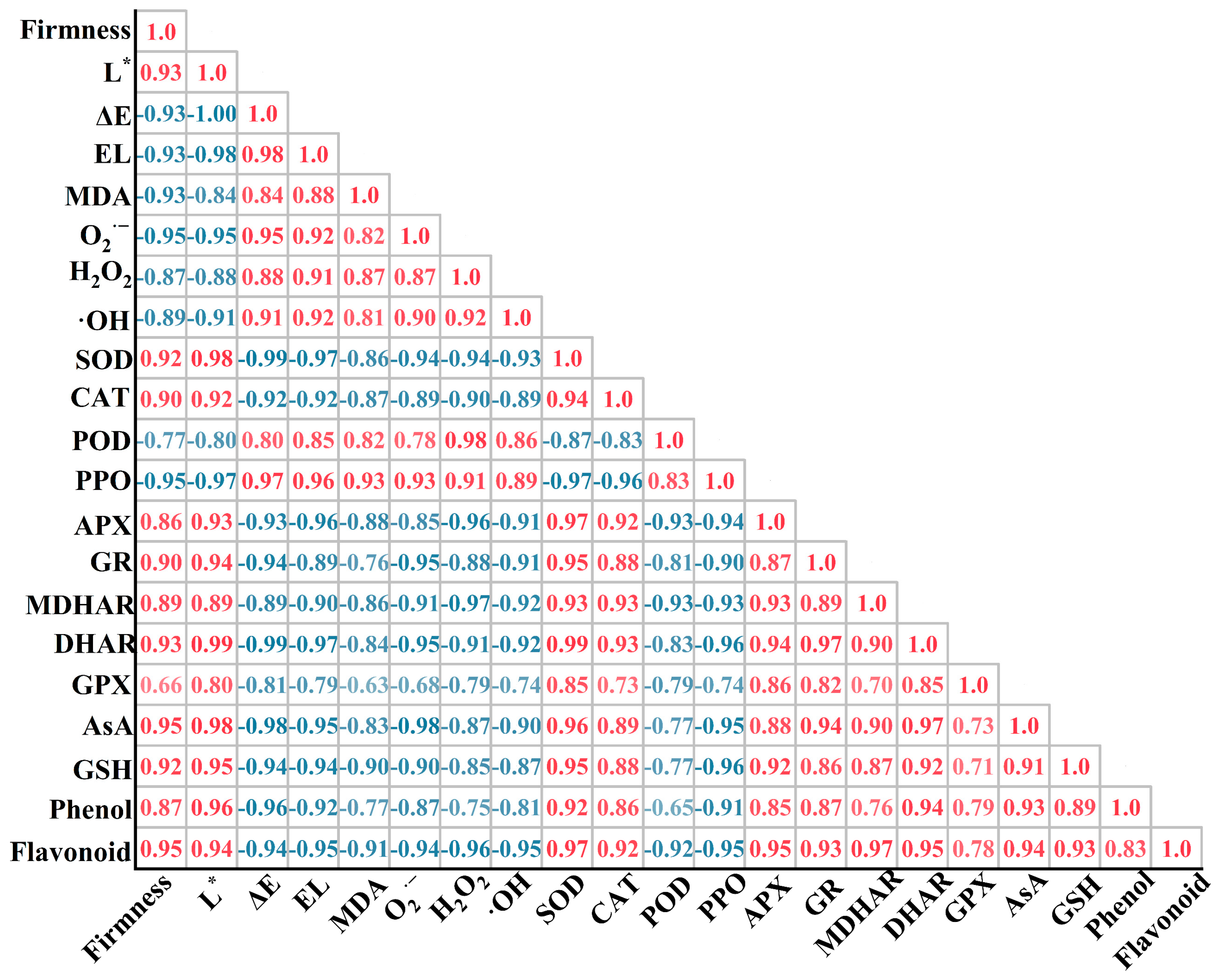
3.6. Correlation between Quality, ROS Damage, Antioxidant Enzymes, and Antioxidants of Agaricus bisporus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sami, R.; Elhakem, A.; Almushhin, A.; Alharbi, M.; Almatrafi, M.; Benajiba, N.; Fikry, M.; Helal, M. Enhancement in physicochemical parameters and microbial populations of mushrooms as influenced by nano-coating treatments. Sci. Rep. 2021, 11, 7915. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Chen, X.; Xin, G.; Qin, S.; Chen, M.; Jiang, F. Effect of 1-methylcyclopropene (1-MCP) on quality of button mushrooms (Agaricus bisporus) packaged in different packaging materials. Postharvest Biol. Technol. 2020, 159, 111023. [Google Scholar] [CrossRef]
- Gowen, A.A.; O’Donnell, C.P.; Taghizadeh, M.; Gaston, E.; O’Gorman, A.; Cullen, P.J.; Frias, J.M.; Esquerre, C.; Downey, G. Hyperspectral imaging for the investigation of quality deterioration in sliced mushrooms (Agaricus bisporus) during storage. Sens. Instrum. Food Qual. Saf. 2008, 2, 133–143. [Google Scholar] [CrossRef]
- Flaherty, E.J.; Lum, G.B.; DeEll, J.R.; Subedi, S.; Shelp, B.J.; Bozzo, G.G. Metabolic alterations in postharvest pear fruit as influenced by 1-Methylcyclopropene and controlled atmosphere storage. J. Agric. Food Chem. 2018, 66, 12989–12999. [Google Scholar] [CrossRef]
- Ma, G.; Zhang, L.; Kato, M.; Ikoma, Y.; Yamawaki, K.; Asai, T.; Nishikawa, F.; Ikoma, Y.; Matsumoto, H. Effect of 1-methylcyclopropene on the expression of genes for ascorbate metabolism in postharvest cauliflower. J. Jpn. Soc. Hortic. Sci. 2011, 80, 512–520. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Lei, H.; Ma, X.; Lai, T.; Song, H.; Shi, X.; Li, J. Antifungal activity of 1-methylcyclopropene (1-MCP) against anthracnose (Colletotrichum gloeosporioides) in postharvest mango fruit and its possible mechanisms of action. Int. J. Food Microbiol. 2017, 241, 1–6. [Google Scholar] [CrossRef]
- Zhang, J.; Lu, J.; Mantri, N.; Jiang, L.; Ying, S.; Chen, S.; Feng, X.; Cao, Y.; Chen, Z.; Ren, L.; et al. An effective combination storage technology to prolong storability, preserve high nutrients and antioxidant ability of astringent persimmon. Sci. Hortic. 2018, 241, 304–312. [Google Scholar] [CrossRef]
- Sahay, S.; Gupta, M. An update on nitric oxide and its benign role in plant responses under metal stress. Nitric Oxide 2017, 67, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.; Wang, Y.; Li, B.; Qin, G.; Tian, S. Defense responses of tomato fruit to exogenous nitric oxide during postharvest storage. Postharvest Biol. Technol. 2011, 62, 127–132. [Google Scholar] [CrossRef]
- Shi, K.; Liu, Z.; Wang, J.; Zhu, S.; Huang, D. Nitric oxide modulates sugar metabolism and maintains the quality of red raspberry during storage. Sci. Hortic. 2019, 256, 108611. [Google Scholar] [CrossRef]
- Meitha, K.; Pramesti, Y.; Suhandono, S. Reactive oxygen species and antioxidants in postharvest vegetables and fruits. Int. J. Food Sci. Technol. 2020, 2020, 8817778. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Liu, M.; Zhou, J. Inhibition by nitric oxide of ethylene biosynthesis and lipoxygenase activity in peach fruit during storage. Postharvest Biol. Technol. 2006, 42, 41–48. [Google Scholar] [CrossRef]
- Ni, X.; Yu, J.; Shao, P.; Yu, J.; Chen, H.; Gao, H. Preservation of Agaricus bisporus freshness with using innovative ethylene manipulating active packaging paper. Food Chem. 2020, 345, 128757. [Google Scholar] [CrossRef]
- Rodrı’guez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A Copper Cofactor for the Ethylene Receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef]
- Gergoff Grozeff, G.E.; Alegre, M.L.; Senn, M.E.; Chaves, A.R.; Simontacchi, M.; Bartoli, C.G. Combination of nitric oxide and 1-MCP on postharvest life of the blueberry (Vaccinium spp.) fruit. Postharvest Biol. Technol. 2017, 133, 72–80. [Google Scholar] [CrossRef]
- Steelheart, C.; Alegre, M.L.; Vera Bahima, J.; Senn, M.E.; Simontacchi, M.; Bartoli, C.G.; Gergoff Grozeff, G.E. Nitric oxide improves the effect of 1-methylcyclopropene extending the tomato (Lycopersicum esculentum L.) fruit postharvest life. Sci. Hortic. 2019, 255, 193–201. [Google Scholar] [CrossRef]
- Gholami, R.; Ahmadi, E.; Farris, S. Shelf life extension of white mushrooms (Agaricus bisporus) by low temperatures conditioning, modified atmosphere, and nanocomposite packaging material. Food Packag. Shelf Life 2017, 14, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Park, D.H.; Park, J.J.; Olawuyi, I.F.; Lee, W.Y. Quality of White mushroom (Agaricus bisporus) under argon- and nitrogen-based controlled atmosphere storage. Sci. Hortic. 2020, 265, 109229. [Google Scholar] [CrossRef]
- Gu, C.S.; Yang, Y.H.; Shao, Y.F.; Wu, K.W.; Liu, Z.L. The effects of exogenous salicylic acid on alleviating cadmium toxicity in Nymphaea tetragona Georgi. S. Afr. J. Bot. 2018, 114, 267–271. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Z.; Sun, Y.; Wang, X.; Li, L. Combined antioxidant and sensory effects of active chitosan/zein film containing α-tocopherol on Agaricus bisporus. Food Packag. Shelf Life 2020, 24, 100470. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, J.; Chen, Y.; Wei, J.; Wu, B. Nitric oxide treatment maintains postharvest quality of table grapes by mitigation of oxidative damage. Postharvest Biol. Technol. 2019, 152, 9–18. [Google Scholar] [CrossRef]
- Zhu, F.; Chen, J.; Xiao, X.; Zhang, M.; Yun, Z.; Zeng, Y.; Xu, J.; Cheng, Y.; Deng, X. Salicylic acid treatment reduces the rot of postharvest citrus fruit by inducing the accumulation of H2O2, primary metabolites and lipophilic polymethoxylated flavones. Food Chem. 2016, 207, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yun, J.; Zhang, Y.; Bi, Y.; Zhao, F.; Niu, Y. Effects of ozone fumigation combined with nano-film packaging on the postharvest storage quality and antioxidant capacity of button mushrooms (Agaricus bisporus). Postharvest Biol. Tecnol. 2021, 176, 111501. [Google Scholar] [CrossRef]
- Hu, Y.H.; Zhou, Y.; Liu, J.A.; Wang, Q.; Lin, J.C.; Shi, Y. Effect of 4-methoxycinnamic acid on the postharvest browning of mushrooms (Agaricus bisporus). J. Food Process. Preserv. 2020, 44, e14735. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous silicon attenuates cadmium-induced oxidative stress in Brassica napus L. by modulating AsA-GSH pathway and glyoxalase system. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Huan, C.; Jiang, L.; An, X.; Kang, R.; Yu, M.; Ma, R.; Yu, Z. Potential role of glutathione peroxidase gene family in peach fruit ripening under combined postharvest treatment with heat and 1-MCP. Postharvest Biol. Technol. 2016, 111, 175–184. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Nie, X.; Zhang, R.; Cheng, L.; Li, S.; Zhao, X.; Chen, X. Combining the biocontrol yeast Pichia kluyveri with UV-C treatment to control postharvest decay of king oyster mushrooms (Pleurotus eryngii) caused by Lactococcus lactis subsp. lactis. Biol. Control 2020, 149, 104327. [Google Scholar] [CrossRef]
- Huang, D.; Wang, Y.; Zhang, D.; Dong, Y.; Meng, Q.; Zhu, S.; Zhang, L. Strigolactone maintains strawberry quality by regulating phenylpropanoid, NO, and H2S metabolism during storage. Postharvest Biol. Technol. 2021, 178, 111546. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Wang, H.; Chen, G.; Shi, L.; Lin, H.; Chen, Y.; Lin, Y.; Fan, Z. Influences of 1-methylcyclopropene-containing papers on the metabolisms of membrane lipids in Anxi persimmons during storage. Food Qual. Saf. 2020, 4, 143–150. [Google Scholar] [CrossRef]
- Mayer, A.M.; Harel, E. Polyphenol oxidases in plants. Phytochemistry 1979, 18, 193–215. [Google Scholar] [CrossRef]
- Xia, R.; Wang, L.; Xin, G.; Bao, X.; Sun, L.; Xu, H.; Hou, Z. Preharvest and postharvest applications of 1-MCP affect umami taste and aroma profiles of mushrooms (Flammulina velutipes). LWT Food Sci. Technol. 2021, 144, 111176. [Google Scholar] [CrossRef]
- Li, L.; Kitazawa, H.; Zhang, X.; Zhang, L.; Sun, Y.; Wang, X.; Liu, Z.; Guo, Y.; Yu, S. Melatonin retards senescence via regulation of the electron leakage of postharvest white mushroom (Agaricus bisporus). Food Chem. 2021, 340, 127833. [Google Scholar] [CrossRef]
- Karuppanapandian, T.; Moon, J.-C.; Kim, C.; Manoharan, K.; Kim, W. Reactive oxygen species in plants: Their generation, signal transduction, and scavenging mechanisms. Aust. J. Crop Sci. 2011, 5, 709–725. [Google Scholar] [CrossRef] [Green Version]
- Corpas, F.J.; Fernandez-Ocana, A.; Carreras, A.; Valderrama, R.; Luque, F.; Esteban, F.J.; Rodriguez-Serrano, M.; Chaki, M.; Pedrajas, J.R.; Sandalio, L.M.; et al. The expression of different superoxide dismutase forms is cell-type dependent in olive (Olea europaea L.) leaves. Plant Cell Physiol. 2006, 47, 984–994. [Google Scholar] [CrossRef] [Green Version]
- Marί, M.; Morales, A.; Colell, A.; GarcίaRuiz, C.; Fernández-Checa, J.C. Mitochondrial glutathione, a key survival antioxidant. Antioxid. Redox Signal. 2009, 11, 2685–2699. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Li-qin, L.; Jun-feng, G. Effects of 1-methylcyclopropene on NO content, NOS activity, and H2O2. Agric. Sci. China 2011, 10, 797–804. [Google Scholar] [CrossRef]
- Wang, H.; Li, Y.; Hou, J.; Huang, J.; Liang, W. Nitrate reductase-mediated nitric oxide production alleviates Al-induced inhibition of root elongation by regulating the ascorbate-glutathione cycle in soybean roots. Plant Soil 2016, 410, 453–465. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Ryan, L. Improving public health?: The role of antioxidant-rich fruit and vegetable beverages. Food Res. Int. 2011, 44, 3135–3148. [Google Scholar] [CrossRef]
- Dong, J.; Zhang, M.; Lu, L.; Sun, L.; Xu, M. Nitric oxide fumigation stimulates flavonoid and phenolic accumulation and enhances antioxidant activity of mushroom. Food Chem. 2012, 135, 1220–1225. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, S.; Dong, Y. Improving storability, physiological disorders, and antioxidant properties of ‘Bartlett’ and ‘d’Anjou’ pears (Pyrus communis L.) by pre-harvest 1-methylcyclopropene spraying. Int. J. Food Sci. Technol. 2020, 56, 115–125. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Yang, Z.; Cui, J.; Zhu, S. Nitric Oxide Made a Major Contribution to the Improvement of Quality in Button Mushrooms (Agaricus bisporus) by the Combined Treatment of Nitric Oxide with 1-MCP. Foods 2022, 11, 3147. https://doi.org/10.3390/foods11193147
Wang X, Yang Z, Cui J, Zhu S. Nitric Oxide Made a Major Contribution to the Improvement of Quality in Button Mushrooms (Agaricus bisporus) by the Combined Treatment of Nitric Oxide with 1-MCP. Foods. 2022; 11(19):3147. https://doi.org/10.3390/foods11193147
Chicago/Turabian StyleWang, Xiaoyu, Zhifeng Yang, Jinxia Cui, and Shuhua Zhu. 2022. "Nitric Oxide Made a Major Contribution to the Improvement of Quality in Button Mushrooms (Agaricus bisporus) by the Combined Treatment of Nitric Oxide with 1-MCP" Foods 11, no. 19: 3147. https://doi.org/10.3390/foods11193147
APA StyleWang, X., Yang, Z., Cui, J., & Zhu, S. (2022). Nitric Oxide Made a Major Contribution to the Improvement of Quality in Button Mushrooms (Agaricus bisporus) by the Combined Treatment of Nitric Oxide with 1-MCP. Foods, 11(19), 3147. https://doi.org/10.3390/foods11193147