
In-Silico Analysis and Antidiabetic Effect of α-Amylase and α-Glucosidase Inhibitory Peptides from Lupin Protein Hydrolysate: Enzyme-Peptide Interaction Study Using Molecular Docking Approach

 ,
, 
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Preparation of Lupin Protein Hydrolysates (LPH)
2.3. Fractionation of the Hydrolysate Samples
2.4. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis
2.5. Determination of α-Amylase Inhibitory Activity
2.6. Determination of α-Glucosidase Inhibitory Activity
2.7. Identification of Peptides Implied in α-Amylase and α-Glucosidase Inhibitory Activities Using Liquid Chromatography-Mass Spectrometry of Quadrupole Time-of-Flight (LC-MS QTOF) and In-Silico Approach
2.8. Molecular Docking
2.9. Statistical Analysis
3. Results and Discussions
3.1. SDS-PAGE Analysis
3.2. Inhibition of the α-Amylase Inhibitory Activity
3.3. Inhibition of the α-Glucosidase Inhibitory Activity
3.4. Identification and Selection of α-Amylase and α-Glucosidase Inhibitory Peptides from Selected Lupin Protein Hydrolysates
3.5. Molecular Interaction of Lupin Protein-Derived α-Glucosidase Inhibitory Peptides with α-Glucosidase
3.6. Molecular Interaction of Lupin Protein-Derived α-Amylase Inhibitory Peptides with α-Amylase
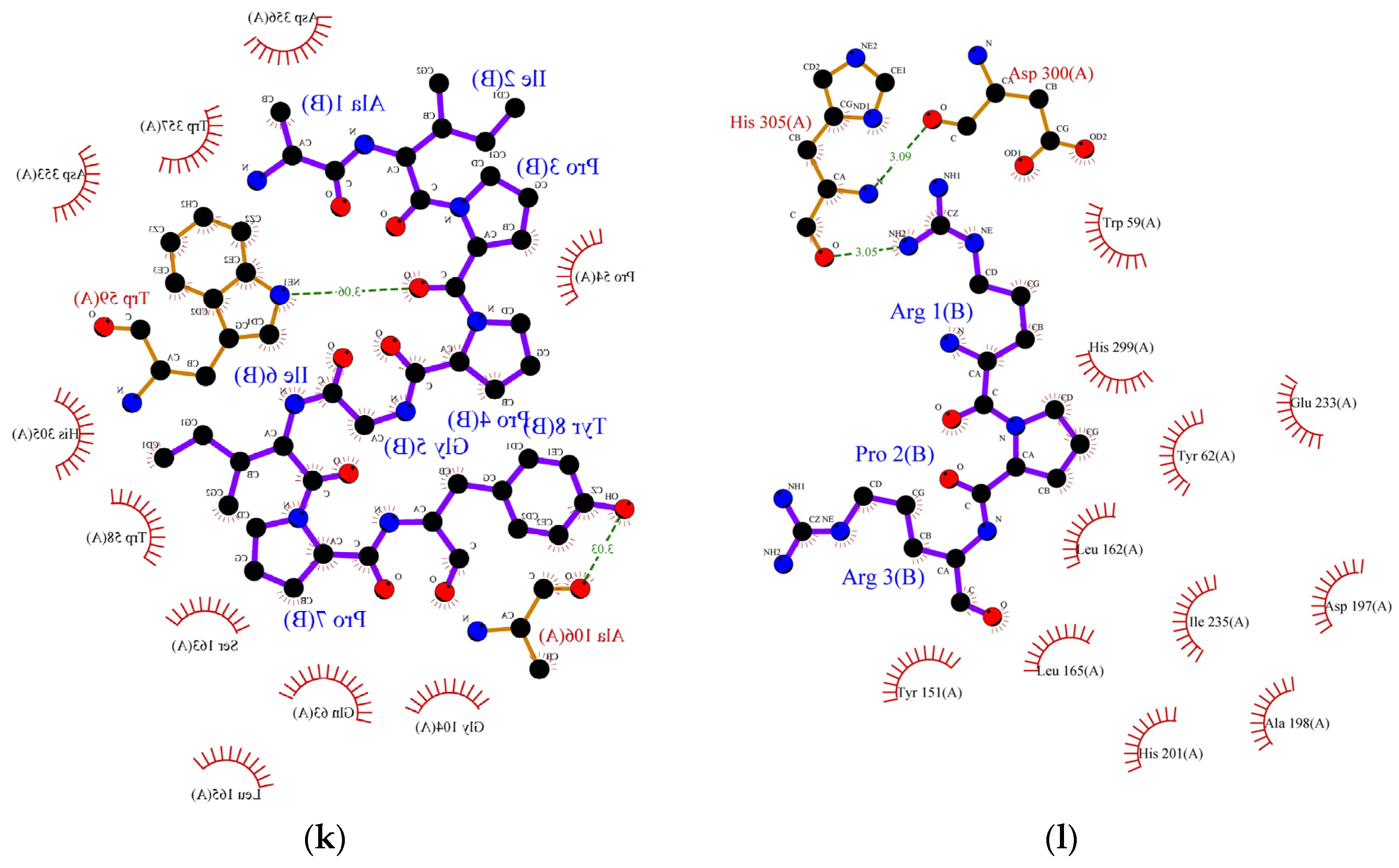
3.7. Mechanism of Molecular Binding and Molecular Docking of Novel Peptides with α-Amylase and α-Glucosidase
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IDF. Diabetes Atlas Tenth Edition; IDF: Brussels, Belgium, 2021. [Google Scholar]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, O.S.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Antioxidant and enzymes inhibitory properties of Amaranth leaf protein hydrolyzates and ultrafiltration peptide fractions. J. Food Biochem. 2021, 45, e13396. [Google Scholar] [CrossRef] [PubMed]
- Patil; Mandal, S.; Tomar, S.K.; Anand, S. Food protein-derived bioactive peptides in management of type 2 diabetes. Eur. J. Nutr. 2015, 54, 863–880. [Google Scholar] [CrossRef] [PubMed]
- Fadimu, G.J.; Gill, H.; Farahnaky, A.; Truong, T. Improving the enzymolysis efficiency of lupin protein by ultrasound pretreatment: Effect on antihypertensive, antidiabetic and antioxidant activities of the hydrolysates. Food Chem. 2022, 383, 132457. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.J.; Karam, S.L.; Wallia, A.; Kang, R.H.; Cooper, A.J.; Lancki, N.; Moran, M.R.; Liss, D.T.; Prospect, T.A.; Ackermann, R.T. Association of second-line Antidiabetic medications with cardiovascular events among insured adults with Type 2 diabetes. JAMA Netw. Open 2018, 1, 186125. [Google Scholar] [CrossRef] [PubMed]
- Nourmohammadi, E.; Mahoonak, A.S. Health implications of bioactive peptides: A review. Int. J. Vitam. Nutr. Res. 2019, 88, 319–343. [Google Scholar] [CrossRef]
- Singh, B.P.; Aluko, R.E.; Hati, S.; Solanki, D. Bioactive peptides in the management of lifestyle-related diseases: Current trends and future perspectives. Crit. Rev. Food Sci. Nutr. 2022, 62, 4593–4606. [Google Scholar] [CrossRef]
- Guo, X.; Shang, W.; Strappe, P.; Zhou, Z.; Blanchard, C. Peptides derived from lupin proteins confer potent protection against oxidative stress. J. Sci. Food Agric. 2018, 98, 5225–5234. [Google Scholar] [CrossRef]
- Kamran, F.; Phillips, M.; Reddy, N. Functional properties of Australian blue lupin (Lupinus angustifolius) protein and biological activities of protein hydrolysates. Legume Sci. 2021, 3, e65. [Google Scholar] [CrossRef]
- Fadimu, G.J.; Farahnaky, A.; Gill, H.; Truong, T. Influence of ultrasonic pretreatment on structural properties and biological activities of lupin protein hydrolysate. Int. J. Food Sci. Technol. 2022, 57, 1729–1738. [Google Scholar] [CrossRef]
- Lammi, C.; Bollati, C.; Lecca, D.; Abbracchio, M.P.; Arnoldi, A. Lupin peptide T9 (GQEQSHQDEGVIVR) modulates the mutant PCSK9D374Y Pathway: In vitro characterization of its dual hypocholesterolemic behavior. Nutrients 2019, 11, 1665. [Google Scholar] [CrossRef]
- Muñoz, E.B.; Luna-Vital, D.A.; Fornasini, M.; Baldeón, M.E.; de Mejia, E.G. Gamma-conglutin peptides from Andean lupin legume (Lupinus mutabilis Sweet) enhanced glucose uptake and reduced gluconeogenesis in vitro. J. Funct. Foods 2018, 45, 339–347. [Google Scholar] [CrossRef]
- Patil, S.P.; Goswami, A.; Kalia, K.; Kate, A.S. Plant-derived bioactive peptides: A treatment to cure diabetes. Int. J. Pept. Res. Ther. 2020, 26, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Fadimu, G.J.; Gill, H.; Farahnaky, A.; Truong, T. Investigating the impact of ultrasound pretreatment on the physicochemical, structural and antioxidant properties of lupin protein hydrolysates. Food Bioproc. Technol. 2021, 14, 2004–2019. [Google Scholar] [CrossRef]
- Laemmli, U. SDS-PAGE Laemmli method. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Wickramaratne, M.N.; Punchihewa, J.; Wickramaratne, D. In-vitro alpha-amylase inhibitory activity of the leaf extracts of Adenanthera pavonina. BMC Complement Altern. Med. 2016, 16, 466. [Google Scholar] [CrossRef] [Green Version]
- Lankatillake, C.; Luo, S.; Flavel, M.; Lenon, G.B.; Gill, H.; Huynh, T.; Dias, D.A. Screening natural product extracts for potential enzyme inhibitors: Protocols, and the standardisation of the usage of blanks in α-amylase, α-glucosidase and lipase assays. Plant Methods 2021, 17, 3. [Google Scholar] [CrossRef]
- Sarah, S.; Faradalila, W.; Salwani, M.; Amin, I.; Karsani, S.; Sazili, A. LC–QTOF-MS identification of porcine-specific peptide in heat treated pork identifies candidate markers for meat species determination. Food Chem. 2016, 199, 157–164. [Google Scholar] [CrossRef]
- Lamiable, A.; Thévenet, P.; Rey, J.; Vavrusa, M.; Derreumaux, P.; Tufféry, P. PEP-FOLD3: Faster de novo structure prediction for linear peptides in solution and in the complex. Nucleic Acids Res. 2016, 44, W449–W454. [Google Scholar] [CrossRef] [Green Version]
- Madhavi Sastry, G.; Adzhigirey, M.; Day, T.; Annabhimoju, R.; Sherman, W. Protein and ligand preparation: Parameters, protocols, and influence on virtual screening enrichments. J. Comput.-Aided Mol. Des. 2013, 27, 221–234. [Google Scholar] [CrossRef]
- Honorato, R.V.; Koukos, P.I.; Jiménez-García, B.; Tsaregorodtsev, A.; Verlato, M.; Giachetti, A.; Rosato, A.; Bonvin, A.M. Structural biology in the clouds: The WeNMR-EOSC ecosystem. Front. Mol. Biosci. 2021, 8, 729513. [Google Scholar] [CrossRef]
- Awosika, T.O.; Aluko, R.E. Inhibition of the in-vitro activities of α-amylase, α-glucosidase and pancreatic lipase by yellow field pea (Pisum sativum L.) protein hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Aluko, R.E. Food protein-derived peptides: Production, isolation, and purification. In Proteins in Food Processing; Elsevier: Amsterdam, The Netherlands, 2018; pp. 389–412. [Google Scholar]
- Girgih, A.T.; Udenigwe, C.C.; Li, H.; Adebiyi, A.P.; Aluko, R.E. Kinetics of enzyme inhibition and antihypertensive effects of hemp seed (Cannabis sativa L.) protein hydrolysates. J. Am. Chem. Soc. 2011, 88, 1767–1774. [Google Scholar] [CrossRef]
- Wei, D.; Fan, W.; Xu, Y. In vitro production and identification of angiotensin-converting enzyme (ACE) inhibitory peptides derived from distilled spent grain prolamin isolate. Foods 2019, 8, 390. [Google Scholar] [CrossRef] [Green Version]
- Trabuco, L.G.; Lise, S.; Petsalaki, E.; Russell, R.B. PepSite: Prediction of peptide-binding sites from protein surfaces. Nucleic Acids Res. 2012, 40, W423–W427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermans, M.; Kroos, M.; Van Beeumen, J.; Oostra, B.; Reuser, A. Human lysosomal alpha-glucosidase. Characterization of the catalytic site. J. Biol. Chem. 1991, 266, 13507–13512. [Google Scholar] [CrossRef]
- Bruckmann, C.; Repo, H.; Kuokkanen, E.; Xhaard, H.; Heikinheimo, P. Systematic Structure-Activity Study on Potential Chaperone Lead Compounds for Acid α-Glucosidase. ChemMedChem 2012, 7, 1943–1953. [Google Scholar] [CrossRef]
- Kamal, H.; Mudgil, P.; Bhaskar, B.; Fisayo, A.F.; Gan, C.-Y.; Maqsood, S. Amaranth proteins as potential source of bioactive peptides with enhanced inhibition of enzymatic markers linked with hypertension and diabetes. J. Cereal Sci. 2021, 101, 103308. [Google Scholar] [CrossRef]
- Roig-Zamboni, V.; Cobucci-Ponzano, B.; Iacono, R.; Ferrara, M.C.; Germany, S.; Bourne, Y.; Parenti, G.; Moracci, M.; Sulzenbacher, G. Structure of human lysosomal acid α-glucosidase–a guide for the treatment of Pompe disease. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Liu, L.; Shin, H.-d.; Chen, R.R.; Li, J.; Du, G.; Chen, J. Structure-based engineering of histidine residues in the catalytic domain of α-amylase from Bacillus subtilis for improved protein stability and catalytic efficiency under acidic conditions. J. Biotechnol. 2013, 164, 59–66. [Google Scholar] [CrossRef]
- Yadav, J.K.; Prakash, V. Stabilization of α-amylase, the key enzyme in carbohydrates properties alterations, at low pH. Int. J. Food Prop. 2011, 14, 1182–1196. [Google Scholar] [CrossRef]
- Buisson, G.; Duee, E.; Haser, R.; Payan, F. Three-dimensional structure of porcine pancreatic alpha-amylase at 2.9 A resolution. Role of calcium in structure and activity. EMBO J. 1987, 6, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Siow, H.-L.; Gan, C.-Y. Extraction, identification, and structure-activity relationship of antioxidative and α-amylase inhibitory peptides from cumin seeds (Cuminum cyminum). J. Funct. Foods 2016, 22, 1–12. [Google Scholar] [CrossRef]
- Yu, X.; Cai, X.; Li, S.; Luo, L.; Wang, J.; Wang, M.; Zeng, L. Studies on the interactions of theaflavin-3, 3′-digallate with bovine serum albumin: Multi-spectroscopic analysis and molecular docking. Food Chem. 2022, 366, 130422. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Hwang, K.Y.; Oh, K.H.; Kim, Y.H.; Lee, J.Y.; Kim, K. Discovery of novel α-glucosidase inhibitors based on the virtual screening with the homology-modeled protein structure. Bioorg. Med. Chem. 2008, 16, 284–292. [Google Scholar] [CrossRef]
- Siow, H.-L.; Lim, T.S.; Gan, C.-Y. Development of a workflow for screening and identification of α-amylase inhibitory peptides from food source using an integrated Bioinformatics-phage display approach: Case study–Cumin seed. Food Chem. 2017, 214, 67–76. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 Values (mg/mL) | |
|---|---|---|
| α-Amylase | α-Glucosidase | |
| ACT | 1.66 ± 0.01 e | 1.65 ± 0.02 d |
| A1kDa | 4.87 ± 0.48 a | 4.51 ± 0.16 a |
| A5kDa | 3.58 ± 0.13 cd | 4.27 ± 0.15 ab |
| A10kDa | 3.85 ± 0.13 c | 3.78 ± 0.19 c |
| FCT | 1.98 ± 0.01 e | 1.91 ± 0.02 d |
| F1kDa | 3.52 ± 0.28 cd | 4.37 ± 0.02 a |
| F5kDa | 4.38 ± 0.01 b | 4.49 ± 0.02 a |
| F10kDa | 3.19 ± 0.04 d | 4.07 ± 0.04 b |
| Peptide Sequence | Peptide Ranker Score | PepSite2 p-Value | Reactive Residues in Peptide | Bound Residues of α-Glucosidase (5NN3) |
|---|---|---|---|---|
| FP | 0.993916 | 0.0004915 | F1, P2 | Asp282, Trp376 *, Trp481, Met519 *, Phe525, Asp616 * |
| RW | 0.978386 | 0.01098 | R1, W2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp518, Asp518 *, Asp616 *, His674 * |
| SPRRF | 0.919692 | 0.001218 | S1, P2, R3, R4 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 *, Phe649 *, His674 * |
| PMLL | 0.89733 | 0.004202 | P1, M2, L3, L4 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Asp616 * |
| ML | 0.894564 | 0.02483 | M1, L2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Asp616 *, His674 * |
| AIPINNPGKL | 0.807084 | 0.003297 | P3, I4, N5, N6, P7, K9 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Trp613 *, Asp616 *, Phe649 *, His674 * |
| MLLL | 0.756994 | 0.04779 | M1, L2, L3, L4 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Asp616 *, Phe649 *, His674 * |
| AIPPGIPY | 0.753808 | 0.008538 | P3, P4, G5, I6, P7 | Asp282, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Arg600 *, Trp613 *, Asp616 *, Phe649 *, His674 * |
| HSDADFIL | 0.683272 | 0.02868 | H1, S2, D3, A4, D5, F6 | Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Asp616 *, Phe649 *, His674 * |
| RLL | 0.607206 | 0.02058 | R1, L2, L3 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Asp616 *, Phe649 *, His674 * |
| FE | 0.589707 | 0.1065 | F1, E2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Trp613 *, Asp616 *, Phe649 *, His674 * |
| LR | 0.569984 | 0.009125 | L1, R2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Asp616 *, Phe649 *, His674 * |
| RR | 0.565498 | 0.001149 | R1, R2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Arg600 *, Asp616 *, Phe649 *, His674 * |
| LRL | 0.564172 | 0.03196 | L1, R2, L3 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Arg600 *, Trp613 *, Asp616 *, Phe649 *, His674 * |
| SVPGCT | 0.501911 | 0.02842 | S1, P3, G4, C5 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 *, Phe649 * |
| LLPH | 0.501446 | 0.001936 | L1, L2, P3, H4 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Asp616 *, Phe649 *, His674 * |
| Peptide Sequence | Peptide Ranker Score | PepSite2 p-Value | Reactive Residues in Peptide | Bound Residues of α-Glucosidase (5NN3) |
|---|---|---|---|---|
| FP | 0.993916 | 0.0004915 | F1, P2 | Asp282, Trp376 *, Trp481, Met519 *, Phe525, Asp616 * |
| ML | 0.894564 | 0.02483 | M1, L2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Asp616 *, His674 * |
| PPGIP | 0.838217 | 0.001023 | P1, P2, G3, I4 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 *, Phe649 * |
| TF | 0.826678 | 0.01348 | T1, F2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Arg600 *, Asp616 * |
| AIPINNPGKL | 0.807084 | 0.003297 | P3, I4, N5, N6, P7, K9 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Trp613 *, Asp616 * |
| LP | 0.79612 | 0.001344 | L1, P2 | Asp282, Trp376 *, Trp481, Met519 *, Phe525, Asp616 *, Phe649 * |
| RPR | 0.722632 | 0.0002172 | R1, P2, R3 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 * |
| LRP | 0.722289 | 0.001416 | L1, R2, P3 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 * |
| FE | 0.589707 | 0.1065 | F1, E2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Trp613 *, Asp616 *, Phe649 *, His674 * |
| RPH | 0.582675 | 0.0001519 | R1, P2, H3 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 *, Phe649 * |
| YL | 0.57536 | 0.1541 | Y1, L2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Asp616 *, Phe649 * |
| LR | 0.569984 | 0.009125 | L1, R2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Asp616 *, Phe649 *, His674 * |
| RR | 0.565498 | 0.001149 | R1, R2 | Trp376 *, Asp404 *, Ile441 *, Trp481, Trp516 *, Asp518 *, Met519 *, Arg600 *, Asp616 *, Phe649 *, His674 * |
| NVLSGFDPQF | 0.514393 | 0.006611 | N1, L3, D10, P8, Q9, F10 | Asp282, Trp376 *, Asp404 *, Ile441 *, Trp481, Asp518 *, Met519 *, Phe525, Arg600 *, Asp616 *, Trp618 |
| Peptide Sequence | Peptide Ranker Score | PepSite2 p-Value | Reactive Residues in Peptide | Bound Residues of α-Amylase (1SMD) |
|---|---|---|---|---|
| FP | 0.993916 | 0.0001681 | F1, P2 | Trp58 *, Trp59 *, Asp300 * |
| RW | 0.978386 | 0.01257 | R1, W2 | His15, Gln41, Trp58 *, Tyr62 *, Arg195 *, Asn298, His299 *, Asp300 *, Arg337 |
| SPRRF | 0.919692 | 0.0007243 | P2, R3, R4, F5 | His15, Phe17, Gln41, Val42, Ser43, Pro44, Trp58 *, Trp59 *, Tyr62 *, Asp96 *, His299 *, Asp300 * |
| PMLL | 0.89733 | 0.00183 | P1, M2, L3, L4 | Phe17, Glu18, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 *, Tyr342 |
| ML | 0.894564 | 0.02935 | M1, L2 | Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| AIPINNPGKL | 0.807084 | 0.003841 | I2, P3, I4, N5, N6, P7 | Phe17, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| MLLL | 0.756994 | 0.03077 | M1, L2, L3, L4 | His15, Phe17, Glu18, Gln41, Val42, Ser43, Pro44, Trp58 *, Trp59 *, Tyr62 *, Asp96 *, His299 *, Asp300 *, Tyr342 |
| AIPPGIPY | 0.753808 | 0.01494 | I2, P3, P4, P7, Y8 | Phe17, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 *, His305 * |
| HSDADFIL | 0.683272 | 0.0521 | A4, D5, F6, I7, L8 | Phe17, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| RLL | 0.607206 | 0.022 | R1, L2, L3 | Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| FE | 0.589707 | 0.08928 | F1, E2 | His15, Gln41, Val42, Ser43, Pro44, Tyr62 *, Asp96 *, Arg195 *, His299 *, Arg337 |
| LR | 0.569984 | 0.0167 | L1, R2 | Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| RR | 0.565498 | 0.003765 | R1, R2 | Phe17, Trp58 *, Tyr62 *, His299 *, Asp300 * |
| LRL | 0.564172 | 0.02945 | L1, R2, L3 | Phe17, Glu18, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 *, Tyr342 |
| SVPGCT | 0.501911 | 0.003133 | V2, P3, G4, C5, T6 | Phe17, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 *, His305 *, Lys352, Asp356 |
| LLPH | 0.501446 | 0.001419 | L1, L2, P3, H4 | Phe17, Glu18, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 *, Tyr342 |
| Peptide Sequence | Peptide Ranker Score | PepSite2 p-Value | Reactive Residues in Peptide | Bound Residues of α-Amylase (1SMD) |
|---|---|---|---|---|
| FP | 0.993916 | 0.0001681 | F1, P2 | Trp58 *, Trp59 *, Asp300 * |
| ML | 0.894564 | 0.02935 | M1, L2 | Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| PPGIP | 0.838217 | 0.001008 | P1, P2, G3, I4 | Trp58 *, Trp59 *, Asp300 *, HIs305 * |
| TF | 0.826678 | 0.02906 | T1, F2 | Trp58 *, Trp59 *, Asp300 * |
| AIPINNPGKL | 0.807084 | 0.003841 | I2, P3, I4, N5, N6, P7 | Phe17, Trp58 *, Trp59 *, His299 *, Asp300 * |
| LP | 0.79612 | 0.0001394 | L1, P2 | Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| RPR | 0.722632 | 0.0001414 | R1, P2, R3 | Phe17, Trp58 *, Trp59 *, Tyr62 *, His299 *, Asp300 * |
| LRP | 0.722289 | 0.0003887 | L1, R2, P3 | Phe17, Glu18, Trp58 *, Trp59 *, Tyr62 *, His299 *, Tyr342 |
| FE | 0.589707 | 0.08928 | F1, E2 | His15, Gln41, Val42, Ser43, Pro44, Tyr62 *, Asp96 *, Arg195 *, His299 *, Arg337 |
| RPH | 0.582675 | 0.0002215 | R1, P2, H3 | Trp58 *, Trp59 *, Tyr62 *, Asp300 * |
| YL | 0.57536 | 0.03237 | Y1, L2 | Trp58 *, Trp59 *, His299 *, Asp300 * |
| LR | 0.569984 | 0.0167 | L1, R2 | Trp58 *, Trp59 *, His299 *, Asp300 * |
| EGDIIAIPPGIP | 0.568358 | |||
| RR | 0.565498 | 0.003765 | R1, R2 | Phe17, Trp58 *, Tyr62 *, His299 *, Asp300 * |
| NVLSGFDPQF | 0.514393 | 0.01987 | V2, L3, S4, F6, P8, F10 | His15, Gln41, Val42, Ser43, Pro44, Trp58 *, Trp59 *, Tyr62 *, Asp96 * |
| Sequence | Binding Affinity (kcal/mol) | Hydrophobicity | Hydrogen Bond | Hydrophobic Interaction |
|---|---|---|---|---|
| α-glucosidase | ||||
| RPR | −6.1 | +11.66 | Asp616 | Ser676, Leu650, Phe649, Leu678, Trp376 * |
| LRP | −6.2 | +8.60 | Asp616 | Leu650, Ser676, Trp376 *, Trp481, Asp282, Phe525 |
| RR | −6.4 | +11.52 | Asp616, Asp518 | Trp376 *, Leu650, Phe649 *, Trp613 |
| SPRRF | −6.6 | +10.41 | Arg411 | Leu677, Ser379, Asp404 *, Asn417, Leu405, Asp419, Met408, Ser410 |
| FE | −6.0 | +9.82 | ND | Arg600, Leu650, Phe649 *, Asp616 *, Trp376 *, Trp481, Met519 |
| PPGIP | −5.3 | +8.35 | ND | Leu650, Leu678, Trp376 *, Trp481 |
| α-amylase | ||||
| ML | −6.9 | +5.98 | Asp300 | Asp356, Trp58 *, Trp59 *, Tyr62 *, Asp197, His305 |
| LP | −7.0 | +6.79 | Trp59, His101 | Ser163, Leu162, Leu165, Gln63, Tyr62 *, Asp197, Asp300 * |
| MLLL | −8.2 | +3.48 | Asp300 | Lys352, Asp356, His305, Ile235, Gly306, His299 *, Glu233, Asp197, Gln63, Tyr62 *, Leu165, Trp58 *, Trp59 * |
| SPRRF | −9.1 | +10.41 | His305, Asp356, Tyr151, Gly306, Asp96, His299, Glu233, Asp300, Asp197 | Ser163, Leu162, Leu165, Trp58 *, Trp59 *, Tyr62 *, Arg195 |
| AIPPGIPY | −8.4 | +7.02 | Trp59, Ala106 | Asp356, Trp357, Asp353, His305 *, Trp58 *, Ser163, Leu165, Gln63, Gly104, Pro54 |
| RPR | −8.2 | +11.66 | Asp300, His305 | Trp59 *, His299 *, Tyr62 *, Glu233, Leu162, Tyr151, Leu165, His201, Ile235, Asp197, Ala198 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fadimu, G.J.; Farahnaky, A.; Gill, H.; Olalere, O.A.; Gan, C.-Y.; Truong, T. In-Silico Analysis and Antidiabetic Effect of α-Amylase and α-Glucosidase Inhibitory Peptides from Lupin Protein Hydrolysate: Enzyme-Peptide Interaction Study Using Molecular Docking Approach. Foods 2022, 11, 3375. https://doi.org/10.3390/foods11213375
Fadimu GJ, Farahnaky A, Gill H, Olalere OA, Gan C-Y, Truong T. In-Silico Analysis and Antidiabetic Effect of α-Amylase and α-Glucosidase Inhibitory Peptides from Lupin Protein Hydrolysate: Enzyme-Peptide Interaction Study Using Molecular Docking Approach. Foods. 2022; 11(21):3375. https://doi.org/10.3390/foods11213375
Chicago/Turabian StyleFadimu, Gbemisola J., Asgar Farahnaky, Harsharn Gill, Olusegun A. Olalere, Chee-Yuen Gan, and Tuyen Truong. 2022. "In-Silico Analysis and Antidiabetic Effect of α-Amylase and α-Glucosidase Inhibitory Peptides from Lupin Protein Hydrolysate: Enzyme-Peptide Interaction Study Using Molecular Docking Approach" Foods 11, no. 21: 3375. https://doi.org/10.3390/foods11213375
APA StyleFadimu, G. J., Farahnaky, A., Gill, H., Olalere, O. A., Gan, C. -Y., & Truong, T. (2022). In-Silico Analysis and Antidiabetic Effect of α-Amylase and α-Glucosidase Inhibitory Peptides from Lupin Protein Hydrolysate: Enzyme-Peptide Interaction Study Using Molecular Docking Approach. Foods, 11(21), 3375. https://doi.org/10.3390/foods11213375