
Studies on Simultaneous Enrichment and Detection of Escherichia coli O157:H7 during Sample Shipment

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Media Preparation
2.2. E. coli O157:H7 Culture Preparation
2.3. Phage Purification Procedure
2.5. Nano-Glo® Reagent Preparation
2.6. Effects of Phage Concentration on Time to Detection
2.7. Characterization of the Growth of E. coli O157:H7 and Corresponding ΦV10nluc Lysogen
2.8. E. coli O157:H7 Detection in Raw Ground Beef
2.8.1. Detection of E. coli O157:H7 in Raw Ground Beef in Stomacher Bags
2.8.2. Detection of E. coli O157:H7 in Raw Ground Beef in Nalgene Bottles
3. Results and Discussion
3.1. Phage Purification
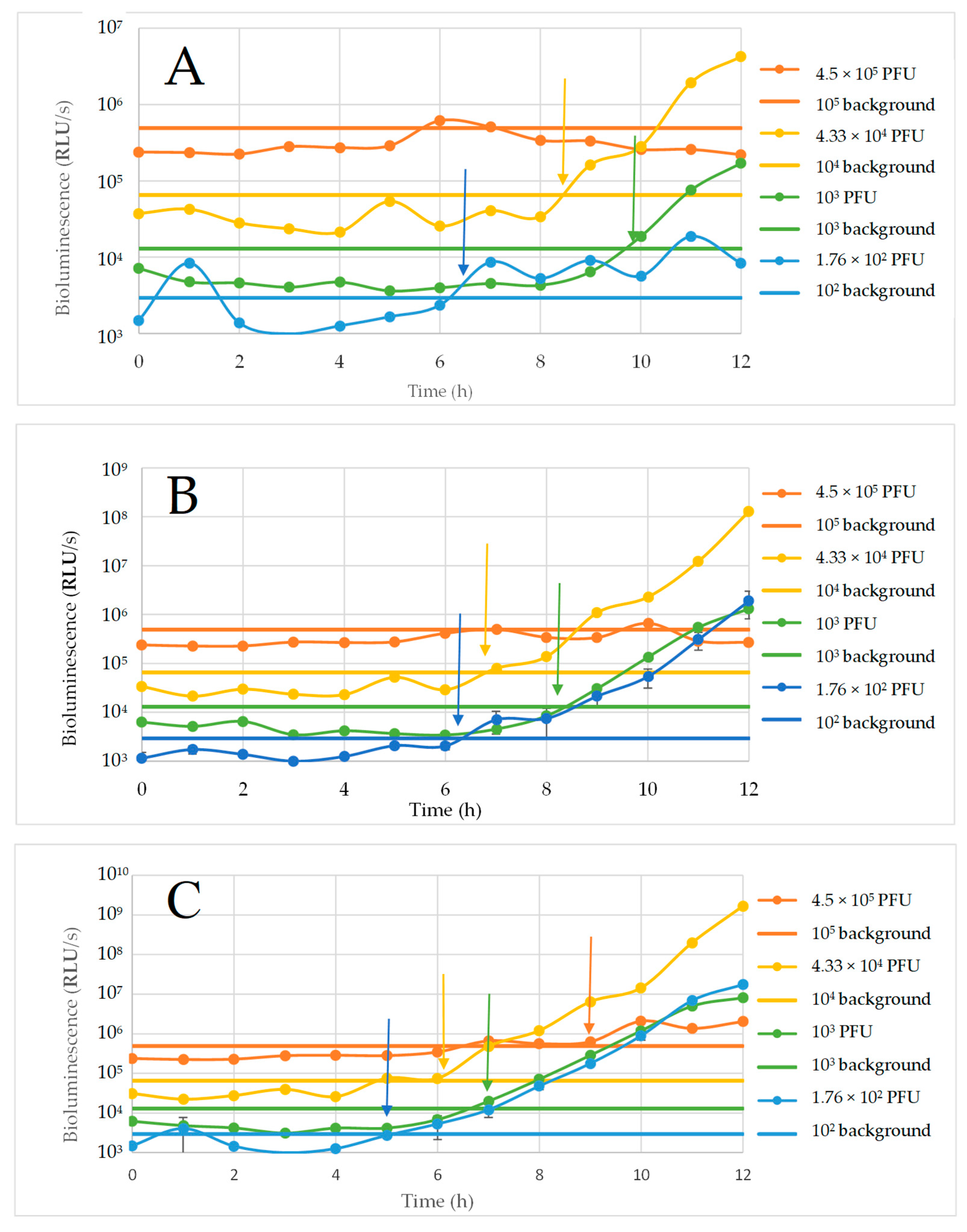
3.2. E. coli O157:H7 Detection by ΦV10nluc Phage in LB
3.3. Characterization of the Growth of E. coli O157:H7 and ΦV10nluc Lysogen
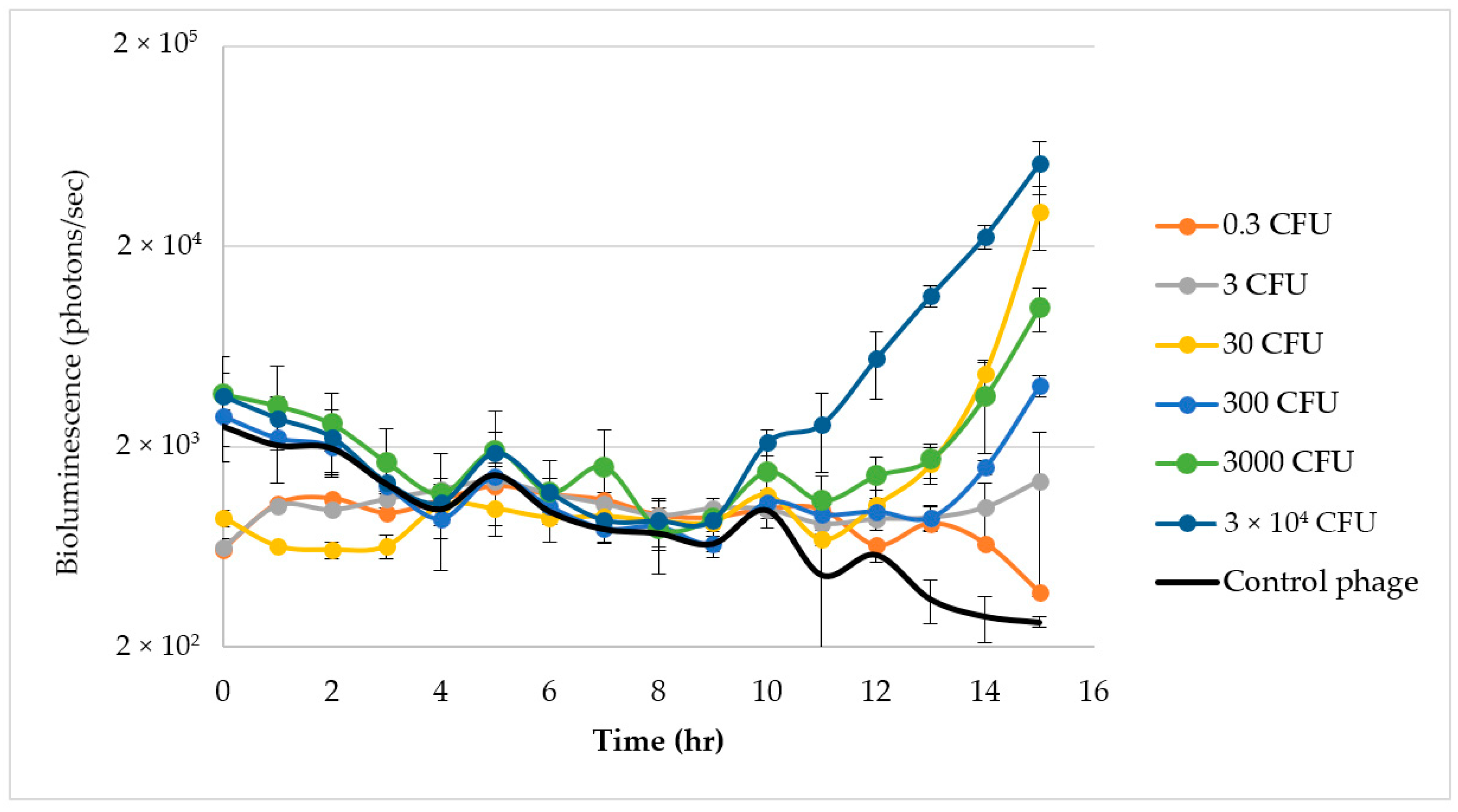
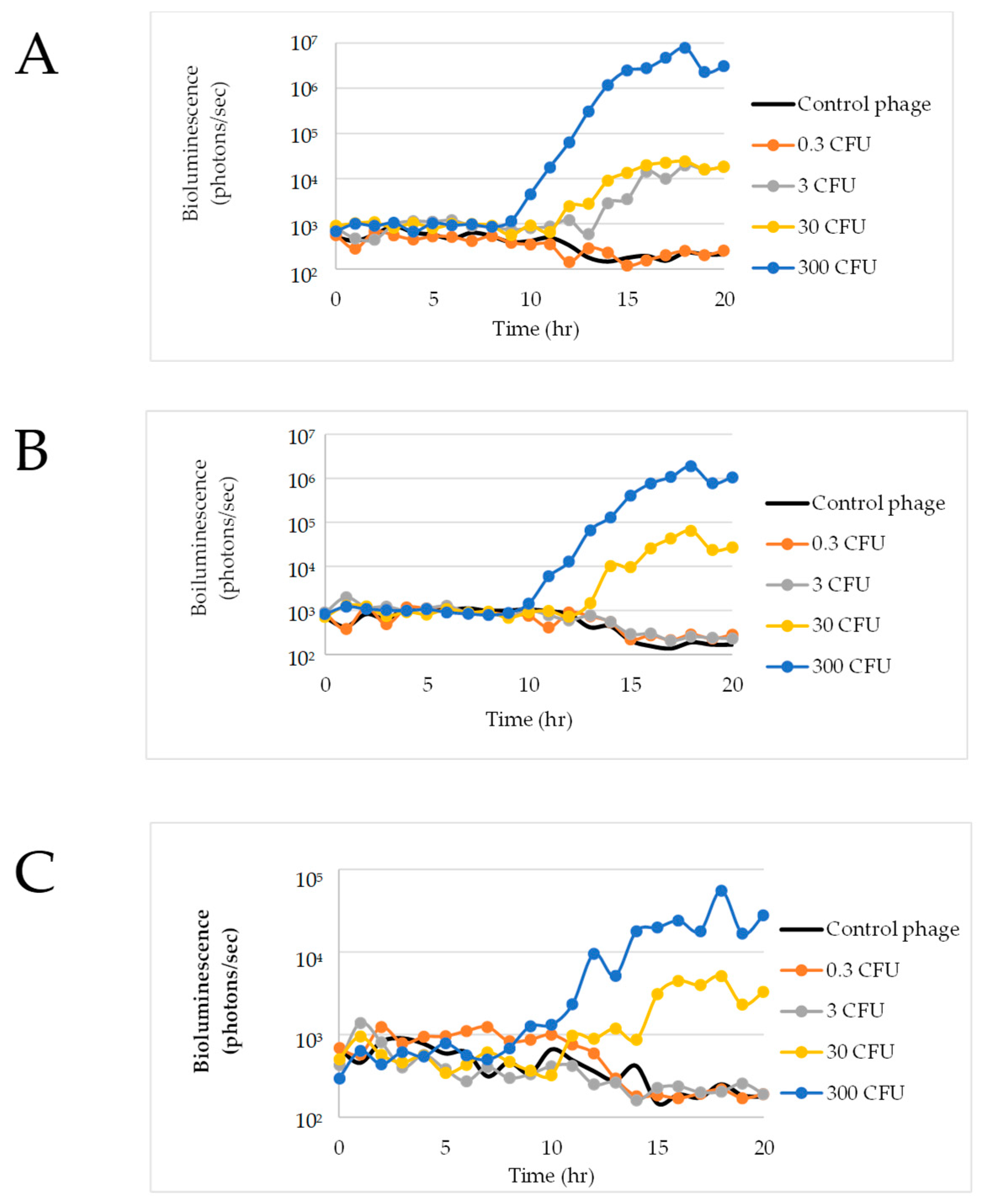
3.4. E. coli O157:H7 Detection in Raw Ground Beef
3.5. Growth of E. coli O157:H7 C7927
3.6. Detection of E. coli O157:H7 in Raw Ground Beef in Nalgene Bottles
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scallan, E.; Hoekstra, R.M.; Angulo, F.J.; Tauxe, R.V.; Widdowson, M.; Roy, S.L.; Jones, J.L.; Griffin, P.M. Foodborne Illness Acquired in the United States—Major Pathogens. Emerg. Infect. Dis. 2011, 17, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Finlay, B.B. Molecular Mechanisms of Escherichia coli Pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Centers of Disease Control and Prevention. E. coli (Escherichia coli). U.S. Department of Health and Human Services. 2018. Available online: https://www.cdc.gov/ecoli/general/index.html (accessed on 15 September 2022).
- Mele, C.; Remuzzi, G.; Noris, M. Hemolytic Uremic syndrome. Sem. Immunopathol. 2014, 36, 399–420. [Google Scholar] [CrossRef] [PubMed]
- EFSA. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2012. EFSA J. 2014, 12, 3547. [Google Scholar] [CrossRef]
- Rangel, J.M.; Sparling, P.H.; Crowe, C.; Griffin, P.M.; Swerdlow, D.L. Epidemiology of Escherichia coli O157:H7 Outbreaks, United States, 1982–2002. Emerg. Infect. Dis. 2005, 11, 603–609. [Google Scholar] [CrossRef]
- Karmali, M.A. Infection by Verocytotoxin-Producing Escherichia coli. Clin. Microbiol. Rev. 1989, 2, 15–38. [Google Scholar] [CrossRef] [PubMed]
- Griffin, P.M.; Tauxe, R.V. The Epidemiology of Infections Caused by Escherichia coli O157:H7, Other Enterohemorrhagic E. coli, and the Associated Hemolytic Uremic Syndrome. Epidemiol. Rev. 1991, 13, 60–98. [Google Scholar] [CrossRef] [PubMed]
- McGowan, K.L.; Wickersham, E.; Strockbine, N.A. Escherichia coli O157:H7 from water (Letter). Lancet 1989, 1, 967–968. [Google Scholar] [CrossRef]
- Swerdlow, D.L.; Woodruff, B.A.; Brady, R.C.; Griffin, P.M.; Tippen, S.; Donnell, H.D., Jr.; Geldreich, E.; Payne, B.J.; Neyer, A., Jr.; Wells, J.G.; et al. A Waterborne Outbreak in Missouri of Escherichia coli O157:H7 Associated with Bloody Diarrhea and Death. Ann. Intern. Med. 1992, 117, 812–819. [Google Scholar] [CrossRef] [PubMed]
- U.S. Department of Agriculture Food Safety and Inspection Service. Report on the Food Safety and Inspection Service’s Microbiological and Residue Sampling Programs. 2011. Available online: https://www.fsis.usda.gov/wps/wcm/connect/0816b926-c7ee-4c24-9222-34ac674ec047/FSIS_Sampling_Programs_Report.pdf?MOD=AJPERES (accessed on 15 September 2022).
- Paoli, G.C.; Wijey, C.; Uhlich, G.A. Genetically Marked Strains of Shiga Toxin-Producing O157:H7 and Non-O157 Escherichia coli: Tools for Detection and Modeling. J. Food Prot. 2015, 78, 888–901. [Google Scholar] [CrossRef] [PubMed]
- Cloke, J.; Crowley, E.; Bird, P.; Bastin, B.; Flanngery, J.; Agin, J.; Goins, D.; Clark, D., Jr.; Radcliff, R.; Wickstrand, N.; et al. Validation of the Thermo Scientific SureTect Escherichia coli O157:H7 Real-Time PCR Assay for Raw Beef and Produce Matrixes. J. AOAC Int. 2015, 98, 1301–1314. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Weagant, S.D.; Jinneman, K. Diarrheagenic Escherichia coli. Bacteriological Analytical Manual Chapter 4A. U.S. Food & Drug Administration. 2020. Available online: https://www.fda.gov/food/laboratory-methods-food/bam-chapter-4a-diarrheagenic-escherichia-coli (accessed on 15 September 2022).
- Panwar, S.; Duggirala, K.S.; Yadav, P.; Debnath, N.; Yadav, A.K.; Kumar, A. Advanced Diagnostic Methods for Identification of Bacterial Foodborne Pathogens: Contemporary and Upcoming Challenges. Crit. Rev. Biotechnol. 2022. advanced online publication. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Hammerl, J.A.; Boone, I.; Jansen, W.; Fohler, S.; Klein, G.; Diekmann, R.; Al Dahouk, S. Overview of Validated Alternative Methods for the Detection of Foodborne Bacterial Pathogens. Trends Food Sci. Technol. 2017, 62, 113–118. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture-Food Safety and Inspection Service. MLG 5C.02. Detection, Isolation, and Identification of Top Seven Shiga Toxin-Producing Escherichia coli (STEC) from Meat Products and Carcass and Environmental Sponges. 2021. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/2021-08/MLG-5C.02.pdf (accessed on 15 September 2022).
- U.S. Department of Agriculture-Food Safety and Inspection Service. MLG Appendix 1.10. Media and Reagents. 2022. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/2022-03/MLG_Appendix_1.10.pdf (accessed on 15 September 2022).
- U.S. Department of Agriculture-Food Safety and Inspection Service. Use of FSIS Regulatory Verification Sampling to Generate Prevalence Estimates. DCC Prevalence Estimate Workgroup. 2012. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/2020-09/Prevalence_Estimates_Report.pdf (accessed on 15 September 2022).
- U.S. Department of Agriculture-Food Safety Inspection Service. Sampling Results for FSIS Regulated Products. 2022. Available online: http://www.fsis.usda.gov/sites/default/files/media_file/documents/Sampling_Project_Results_Data_20210701_20220630.pdf (accessed on 15 September 2022).
- U.S. Department of Agriculture-Food Safety and Inspection Service. Raw Beef Product Sampling. Inspection Methods. 2021. Available online: https://www.fsis.usda.gov/sites/default/files/media_file/2021-11/18_IM_Raw-Beef-Product-Sampling-11032021.pdf (accessed on 15 September 2022).
- Zhang, D.; Coronel-Aguilera, C.P.; Romero, P.L.; Perry, L.; Minocha, U.; Rosenfield, C.; Gehring, A.; Paoli, G.C.; Bhunia, A.K.; Applegate, B. The Use of a Novel NanoLuc-Based Reporter Phage for the Detection of Escherichia coli O157:H7. Sci. Rep. 2016, 6, 33235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, M.P.; Unch, J.; Binkowski, B.F.; Valley, M.P.; Butler, B.L.; Wood, M.G.; Otto, P.; Zimmerman, K.; Vidugiris, G.; Machleidt, T.; et al. Engineered Luciferease Reporter from a Deep Sea Shrimp Utilizing a Novel Imidazopyrazinone Substrate. ACS Chem. Biol. 2012, 7, 1848–1857. [Google Scholar] [CrossRef] [PubMed]
- Khakhria, R.; Duck, D.; Lior, H. Extended Phage-Typing Scheme for Escherichia coli O157:H7. Epidemiol. Infect. 1990, 105, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Perry, L.L.; SanMiguel, P.; Minocha, U.; Terekhov, A.I.; Shroyer, M.L.; Farris, L.A.; Bright, N.; Reuhs, B.L.; Applegate, B.M. Sequence Analysis of Escherichia coli O157:H7 Bacteriophage ΦV10 and Identification of a Phage-encoded Immunity Protein that Modifies the O157 Antigen. FEMS Microbiol. Let. 2009, 292, 182–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, G.S.A.; Williams, P. Lux Genes and the Applications of Bacterial Bioluminescence. J. General Microbiol. 1992, 138, 1289–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, T.; Doyle, M.; Besser, R. Fate of Enterohemorrhagic Escherichia coli O157:H7 in Apple Cider with and without Preservatives. Appl. Environ. Microbiol. 1993, 59, 2526–2530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonilla, N.; Rojas, M.I.; Cruz, G.N.F.; Hung, S.; Rohwer, F.; Barr, J.J. Phage on Tap—A Quick and Efficient Protocol for the Preparation of Bacteriophage Laboratory Stocks. PeerJ 2016, 4, e2261. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Cabezas, E. Bacteriophage Infection in Ground Beef. In Undergraduate Research Symposium; Purdue University: West Lafayette, IN, USA, 2016. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Centrifugation Cycle | Replicates | ||
|---|---|---|---|
| Filtrate Luminescence (RLU/s) | |||
| 1 | 2 | 3 | |
| 1 | 419,040,100 | 371,692,000 | 349,226,900 |
| 2 | 76,547,500 | 64,513,800 | 58,367,400 |
| 3 | 9,490,580 | 5,744,485 | 8,108,047 |
| 4 | 382,266 | 228,219 | 532,749 |
| 5 | 21,660 | 16,011 | 30,201 |
| 6 | 2954 | 3181 | 3045 |
| 7 | 3486 | 4083 | 2552 |
| Phage Concentration (pfu/mL) | Luminescence Prior to Purification (RLU/s) | Luminescence After Purification (RLU/s) |
|---|---|---|
| 7.20 × 105 | 5.39 × 108 | 6.08 × 103 |
| 1.62 × 105 | 7.08 × 108 | 8.08 × 103 |
| 1.62 × 105 | 8.62 × 108 | 1.38 × 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Coronel-Aguilera, C.P.; Applegate, B.M.; Gehring, A.G.; Bhunia, A.K.; Paoli, G.C. Studies on Simultaneous Enrichment and Detection of Escherichia coli O157:H7 during Sample Shipment. Foods 2022, 11, 3653. https://doi.org/10.3390/foods11223653
Chen C, Coronel-Aguilera CP, Applegate BM, Gehring AG, Bhunia AK, Paoli GC. Studies on Simultaneous Enrichment and Detection of Escherichia coli O157:H7 during Sample Shipment. Foods. 2022; 11(22):3653. https://doi.org/10.3390/foods11223653
Chicago/Turabian StyleChen, Chuyan, Claudia P. Coronel-Aguilera, Bruce M. Applegate, Andrew G. Gehring, Arun K. Bhunia, and George C. Paoli. 2022. "Studies on Simultaneous Enrichment and Detection of Escherichia coli O157:H7 during Sample Shipment" Foods 11, no. 22: 3653. https://doi.org/10.3390/foods11223653
APA StyleChen, C., Coronel-Aguilera, C. P., Applegate, B. M., Gehring, A. G., Bhunia, A. K., & Paoli, G. C. (2022). Studies on Simultaneous Enrichment and Detection of Escherichia coli O157:H7 during Sample Shipment. Foods, 11(22), 3653. https://doi.org/10.3390/foods11223653