
Increase the Content of Ester Compounds in Blueberry Wine Fermentation with the Ester-Producing Yeast: Candida glabrata, Pichia anomala, and Wickerhamomyces anomalus

Abstract
:1. Introduction
2. Materials and Methods
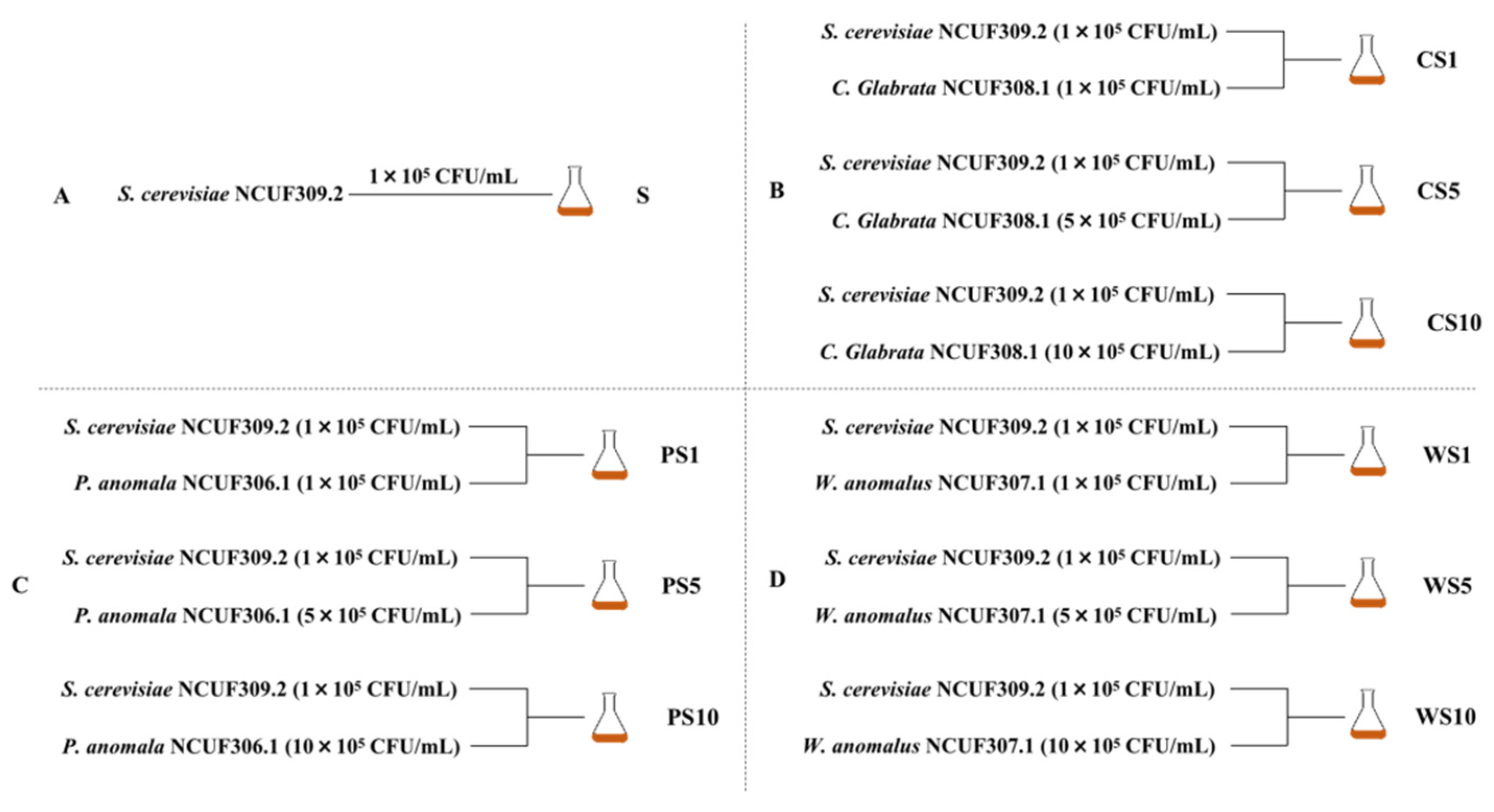
2.1. Strains and Media
2.2. Fermentation Conditions
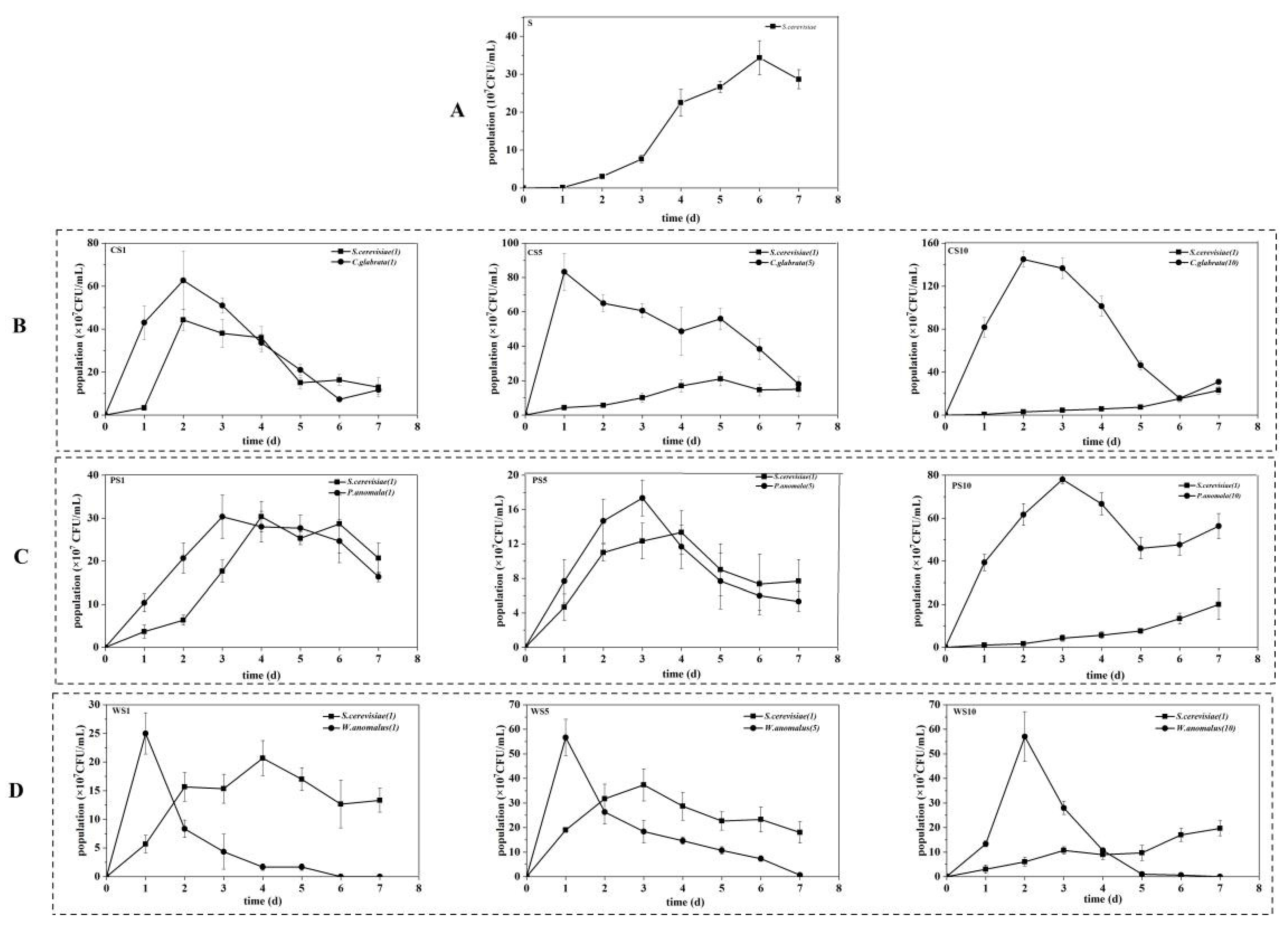
2.3. Determination of Growth of Yeasts
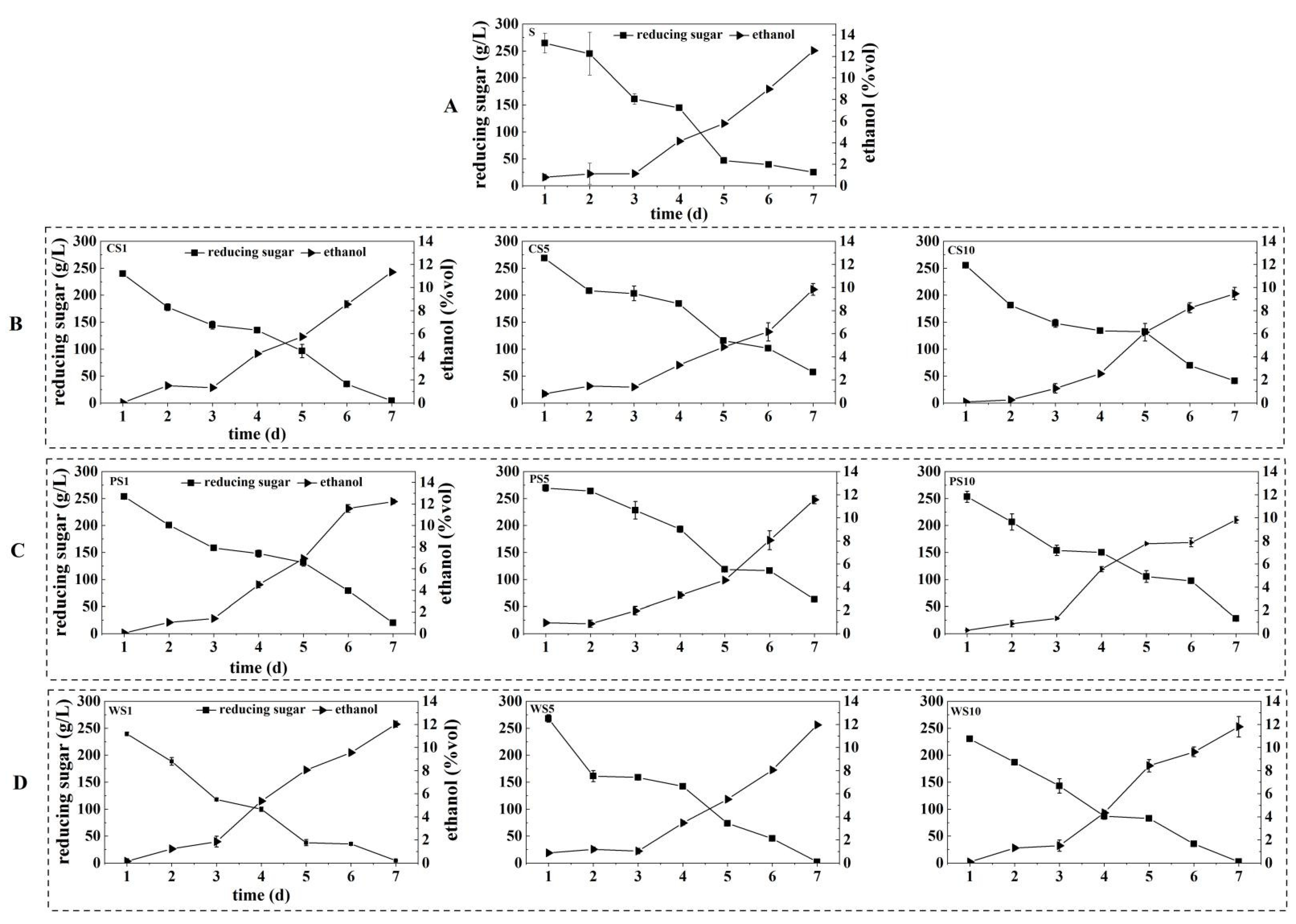
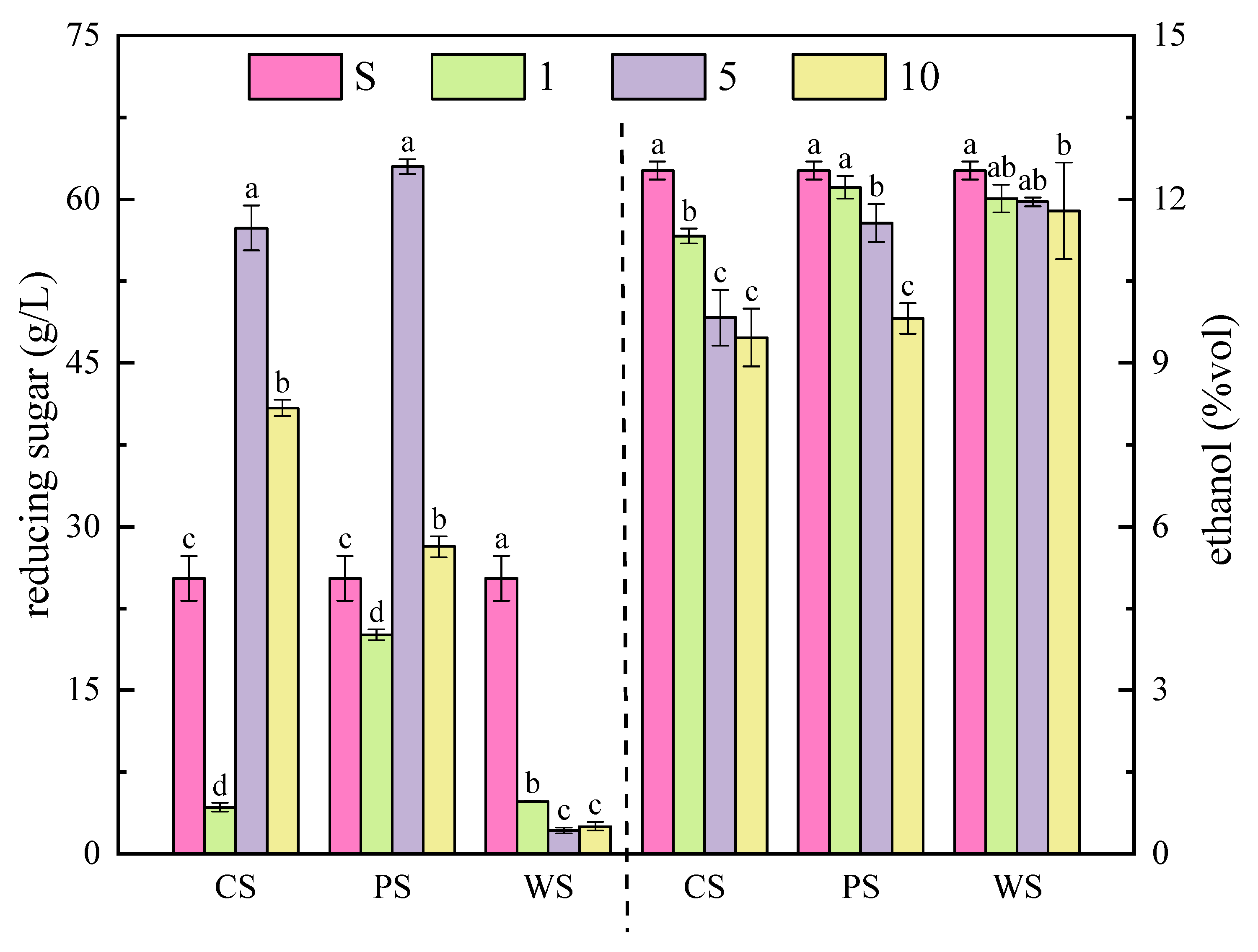
2.4. Determination of the Content of Reducing Sugar and Ethanol
2.5. Analysis of Ester Compounds
2.6. Data Analysis
3. Results and Discussion
3.1. Growth of Yeasts during the Simulated Fermentation of Blueberry Wine
3.2. The Content of Reducing Sugar and Ethanol during the Simulated Fermentation of Blueberry Wine
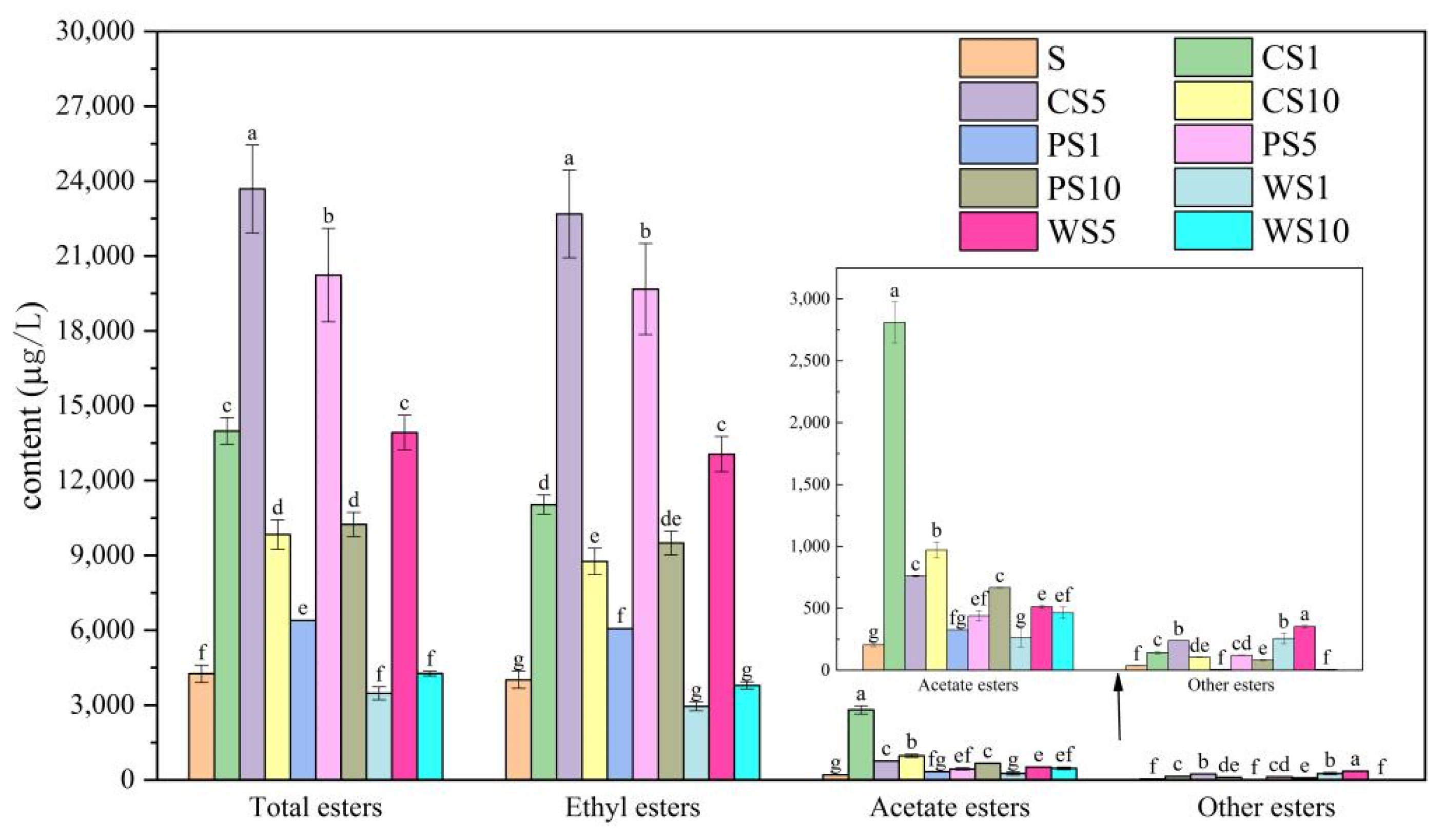
3.3. Accumulation of Ester Compounds during the Simulated Fermentation of Blueberry Wine
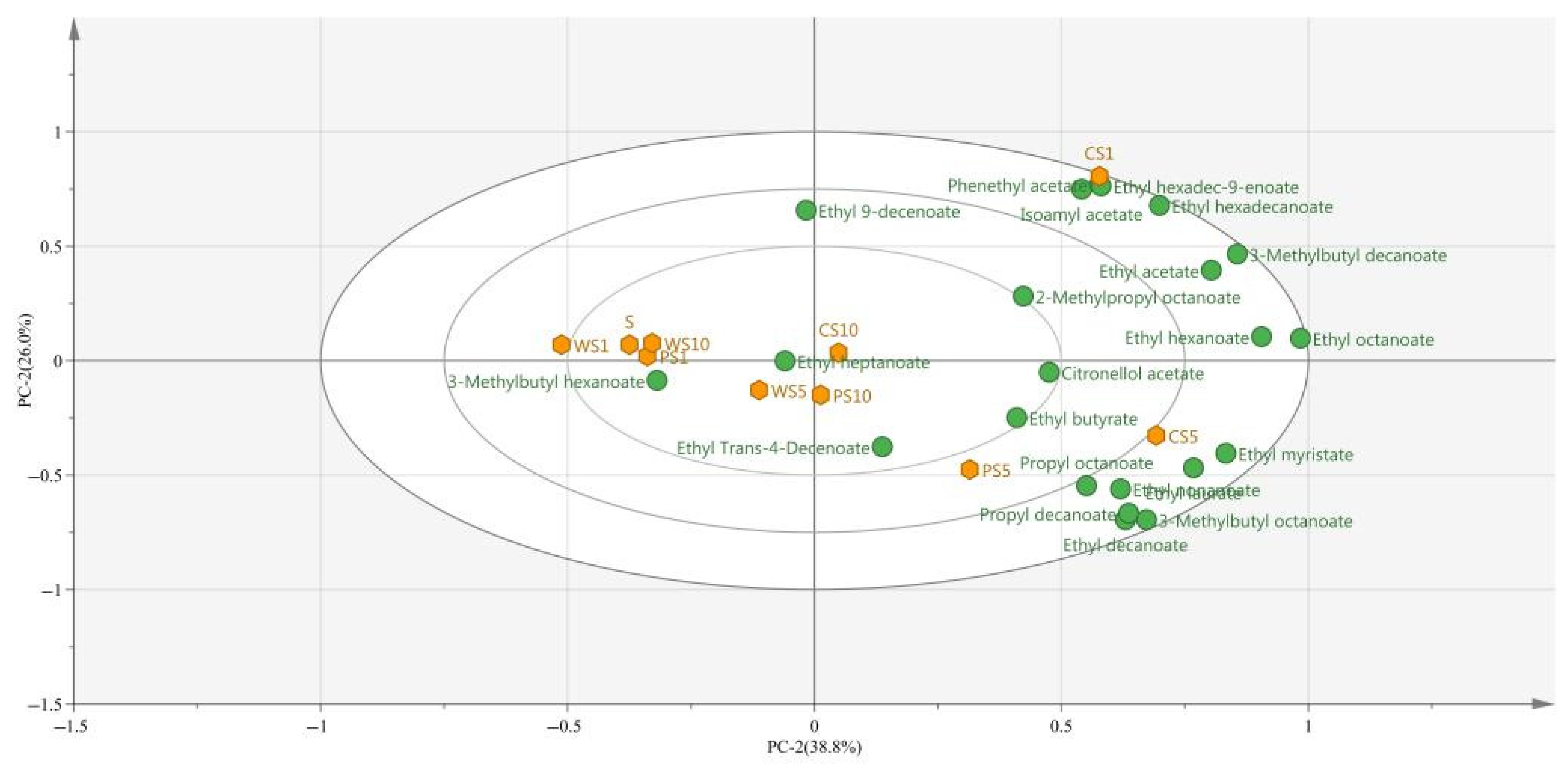
3.4. Principal Component Analysis (PCA) of Ester Compounds during the Simulated Fermentation of Blueberry Wine
3.5. Volatile Odor Active Ester Compounds during the Simulated Fermentation of Blueberry Wine
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, X.; Yan, Z.; Zhu, T.; Zhu, J.; Wang, Y.; Li, B.; Meng, X. Effects on the color, taste, and anthocyanins stability of blueberry wine by different contents of mannoprotein. Food Chem. 2019, 279, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Shokri, S.; Gao, B.; Xu, Z.; Li, B.; Zhu, T.; Wang, Y.; Zhu, J. Improving effects of three selected co-pigments on fermentation, color stability, and anthocyanins content of blueberry wine. Lwt-Food Sci. Technol. 2022, 156, 113070. [Google Scholar] [CrossRef]
- Zhong, W.; Liu, S.; Yang, H.; Li, E. Effect of selected yeast on physicochemical and oenological properties of blueberry wine fermented with citrate-degrading Pichia fermentans. Lwt-Food Sci. Technol. 2021, 145, 111261. [Google Scholar] [CrossRef]
- Alonso-del-Real, J.; Contreras-Ruiz, A.; Castiglioni, G.L.; Barrio, E.; Querol, A. The use of mixed populations of Saccharomyces cerevisiae and S. kudriavzevii to reduce ethanol content in wine: Limited Aeration, inoculum proportions, and sequential inoculation. Front. Microbiol. 2017, 8, 2087. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Yuan, Y.; Dai, L.; Yue, T. Characteristic fruit wine production via reciprocal selection of juice and non-Saccharomyces species. Food Microbiol. 2019, 79, 66–74. [Google Scholar] [CrossRef]
- Escribano, R.; Gonzalez-Arenzana, L.; Portu, J.; Garijo, P.; Lopez-Alfaro, I.; Lopez, R.; Santamaria, P.; Gutierrez, A.R. Wine aromatic compound production and fermentative behaviour within different non-Saccharomyces species and clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef]
- Li, S.; Bi, P.; Sun, N.; Gao, Z.; Chen, X.; Guo, J. Effect of sequential fermentation with four non-Saccharomyces and Saccharomyces cerevisiae on nutritional characteristics and flavor profiles of kiwi wines. J. Food Compos. Anal. 2022, 109, 104480. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Challenges of the Non-Conventional Yeast Wickerhamomyces anomalus in Winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Ivanova-Petropulos, V.; Duan, C.; Yan, G. Distinctive chemical and aromatic composition of red wines produced by Saccharomyces cerevisiae co-fermentation with indigenous and commercial non-Saccharomyces strains. Food Biosci. 2021, 41, 100925. [Google Scholar] [CrossRef]
- Zhang, B.Q.; Shen, J.Y.; Duan, C.Q.; Yan, G.L. Use of indigenous Hanseniaspora vineae and Metschnikowia pulcherrima co-fermentation with Saccharomyces cerevisiae to improve the aroma diversity of vidal blanc icewine. Front. Microbiol. 2018, 9, 2303. [Google Scholar] [CrossRef]
- Hernandez-Orte, P.; Cersosimo, M.; Loscos, N.; Cacho, J.; Garcia-Moruno, E.; Ferreira, V. The development of varietal aroma from non-floral grapes by yeasts of different genera. Food Chem. 2008, 107, 1064–1077. [Google Scholar] [CrossRef]
- Pretorius, I.S. Tailoring wine yeast for the new millennium: Novel approaches to the ancient art of winemaking. Yeast 2000, 16, 675–729. [Google Scholar] [CrossRef]
- Zhu, F.; Du, B.; Li, J. Aroma enhancement and enzymolysis regulation of grape wine using beta-glycosidase. Food Sci. Nutr. 2014, 2, 139–145. [Google Scholar] [CrossRef] [PubMed]
- van Wyk, N.; Grossmann, M.; Wendland, J.; von Wallbrunn, C.; Pretorius, I.S. The whiff of wine yeast innovation: Strategies for enhancing aroma production by yeast during wine fermentation. J. Agric. Food Chem. 2019, 67, 13496–13505. [Google Scholar] [CrossRef] [PubMed]
- Swangkeaw, J.; Vichitphan, S.; Butzke, C.E.; Vichitphan, K. Characterization of beta-glucosidases from Hanseniaspora sp and Pichia anomala with potentially aroma-enhancing capabilities in juice and wine. World J. Microbiol. Biotechnol. 2011, 27, 423–430. [Google Scholar] [CrossRef]
- Rossouw, D.; Bauer, F.F. Exploring the phenotypic space of non-Saccharomyces wine yeast biodiversity. Food Microbiol. 2016, 55, 32–46. [Google Scholar] [CrossRef]
- Aung, M.T.; Lee, P.; Yu, B.; Liu, S. Cider fermentation with three Williopsis saturnus yeast strains and volatile changes. Ann. Microbiol. 2015, 65, 921–928. [Google Scholar] [CrossRef]
- Ye, M.; Yue, T.; Yuan, Y. Effects of sequential mixed cultures of Wickerhamomyces anomalus and Saccharomyces cerevisiae on apple cider fermentation. FEMS Yeast Res. 2014, 14, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Duarte, W.F.; Amorim, J.C.; Schwan, R.F. The effects of co-culturing non-Saccharomyces yeasts with S-cerevisiae on the sugar cane spirit (cacha double dagger a) fermentation process. Anton. Leeuw. Int. J. G. 2013, 103, 175–194. [Google Scholar] [CrossRef]
- Hu, K.; Jin, G.J.; Mei, W.; Li, T.; Tao, Y.S. Increase of medium-chain fatty acid ethyl ester content in mixed H. uvarum/S. cerevisiae fermentation leads to wine fruity aroma enhancement. Food Chem. 2018, 239, 495–501. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non- Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Deng, M.F.; Chen, Y.R.; Wan, Y.; Liu, C.M.; Fu, G.M. Identification and fermentation characteristics of ester-producing yeast from high acidity fruit wine. China Brew. 2020, 39, 103–108. [Google Scholar]
- Rossouw, D.; Du Toit, M.; Bauer, F.F. The impact of co-inoculation with Oenococcus oeni on the trancriptome of Saccharomyces cerevisiae and on the flavour-active metabolite profiles during fermentation in synthetic must. Food Microbiol. 2012, 29, 121–131. [Google Scholar] [CrossRef]
- Yin, L.; Wang, C.; Zhu, X.; Ning, C.; Gao, L.; Zhang, J.; Wang, Y.; Huang, R. A multi-step screening approach of suitable non-Saccharomyces yeast for the fermentation of hawthorn wine. Lwt-Food Sci. Technol. 2020, 127, 109432. [Google Scholar] [CrossRef]
- Shi, W.; Wang, J.; Chen, F.; Zhang, X. Effect of Issatchenkia terricola and Pichia kudriavzevii on wine flavor and quality through simultaneous and sequential co-fermentation with Saccharomyces cerevisiae. Lwt-Food Sci. Technol. 2019, 116, 108477. [Google Scholar] [CrossRef]
- Ulrich, D.; Hoberg, E.; Neugebauer, W.; Tiemann, H.; Darsow, U. Investigation of the boiled potato flavor by human sensory and instrumental methods. Am. J. Potato Res. 2000, 77, 111–117. [Google Scholar] [CrossRef]
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional yeast species for lowering ethanol content of wines. Front. Microbiol. 2016, 7, 642. [Google Scholar] [CrossRef] [Green Version]
- Bagheri, B.; Bauer, F.F.; Setati, M.E. The Impact of Saccharomyces cerevisiae on a wine yeast consortium in natural and inoculated fermentations. Front. Microbiol. 2017, 8, 1988–2000. [Google Scholar] [CrossRef]
- Ge, Q.; Guo, C.; Zhang, J.; Yan, Y.; Zhao, D.; Li, C.; Sun, X.; Ma, T.; Yue, T.; Yuan, Y. Effects of simultaneous co-fermentation of five indigenous non-Saccharomyces Strains with S. cerevisiae on vidal icewine aroma quality. Foods 2021, 10, 1452. [Google Scholar] [CrossRef]
- Taillandier, P.; Quoc Phong, L.; Julien-Ortiz, A.; Brandam, C. Interactions between Torulaspora delbrueckii and Saccharomyces cerevisiae in wine fermentation: Influence of inoculation and nitrogen content. World J. Microbiol. Biotechnol. 2014, 30, 1959–1967. [Google Scholar] [CrossRef] [PubMed]
- Luyt, N.A.; Beaufort, S.; Divol, B.; Setati, M.E.; Taillandier, P.; Bauer, F.F. Phenotypic characterization of cell-to-cell interactions between two yeast species during alcoholic fermentation. World J. Microbiol. Biotechnol. 2021, 37, 186. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Yan, X.; Wang, Q.; Zhang, Y.; Tao, Y. Performance of selected P. fermentans and its excellular enzyme in co-inoculation with S. cerevisiae for wine aroma enhancement. Lwt-Food Sci. Technol. 2017, 86, 361–370. [Google Scholar] [CrossRef]
- Belda, I.; Navascues, E.; Marquina, D.; Santos, A.; Calderon, F.; Benito, S. Dynamic analysis of physiological properties of Torulaspora delbrueckii in wine fermentations and its incidence on wine quality. Appl. Microbiol. Biotechnol. 2015, 99, 1911–1922. [Google Scholar] [CrossRef] [Green Version]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Zhang, Y.; Qiu, Y.; Guo, H.; Ju, H.; Wang, Y.; Yuan, Y.; Yue, T. Chemical composition, sensorial properties, and aroma-active compounds of ciders fermented with Hanseniaspora osmophila and Torulaspora quercuum in co- and sequential fermentations. Food Chem. 2020, 306, 125623. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The oenological potential of Hanseniaspora uvarum in simultaneous and sequential co-fermentation with Saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [Green Version]
- Lee, P.; Kho, S.H.C.; Yu, B.; Curran, P.; Liu, S. Yeast ratio is a critical factor for sequential fermentation of papaya wine by Williopsis saturnus and Saccharomyces cerevisiae. Microb. Biotechnol. 2013, 6, 385–393. [Google Scholar] [CrossRef]
- Hu, L.; Liu, R.; Wang, X.; Zhang, X. The sensory quality improvement of citrus wine through co-fermentations with selected non-Saccharomyces yeast strains and Saccharomyces cerevisiae. Microorganisms 2020, 8, 323. [Google Scholar] [CrossRef] [Green Version]
- Stribny, J.; Gamero, A.; Perez-Torrado, R.; Querol, A. Saccharomyces kudriavzevii and Saccharomyces uvarum differ from Saccharomyces cerevisiae during the production of aroma-active higher alcohols and acetate esters using their amino acidic precursors. Int. J. Food Microbiol. 2015, 205, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Januszek, M.; Satora, P.; Tarko, T. Oenological characteristics of fermented apple musts and volatile profile of brandies obtained from different apple cultivars. Biomolecules 2020, 10, 853. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, H.; Xue, J.; Tang, C.; Duan, C.; Yan, G. Use of Torulaspora delbrueckii and Hanseniaspora vineae co-fermentation with Saccharomyces cerevisiae to improve aroma profiles and safety quality of Petit Manseng wines. Lwt-Food Sci. Technol. 2022, 161, 113360. [Google Scholar] [CrossRef]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef]
- Pu, D.; Zhang, Y.; Zhang, H.; Sun, B.; Ren, F.; Chen, H.; Tang, Y. Characterization of the key aroma compounds in traditional hunan smoke-cured pork leg (Larou, THSL) by aroma extract dilution analysis (AEDA), odor activity value (OAV), and sensory evaluation experiments. Foods 2020, 9, 413. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Li, D.; Ren, L.; Song, S.; Ma, X.; Rong, Y. Effects of simultaneous and sequential cofermentation of Wickerhamomyces anomalus and Saccharomyces cerevisiae on physicochemical and flavor properties of rice wine. Food Sci. Nutr. 2021, 9, 71–86. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Farina, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Wang, J.; Capone, D.L.; Wilkinson, K.L.; Jeffery, D.W. Chemical and sensory profiles of rose wines from Australia. Food Chem. 2016, 196, 682–693. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Content (μg/L) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S | CS1 | CS5 | CS10 | PS1 | PS5 | PS10 | WS1 | WS5 | WS10 | |
| Acetate esters | ||||||||||
| Ethyl acetate | 8.28 ± 1.24 f | 326.48 ± 11.65 a | 302.71 ± 1.75 b | 107.51 ± 2.05 c | 15.09 ± 0.1 f | 23.68 ± 1.12 e | 11.43 ± 1.50 f | 14.17 ± 3.33 f | 33.76 ± 5.63 d | 36.49 ± 6.64 d |
| Isoamyl acetate | 24.93 ± 1.78 f | 805.01 ± 39.97 a | 126.93 ± 25.73 d | 456.2 ± 30.69 b | 73.34 ± 2.18 e | 79.11 ± 5.12 e | 186.68 ± 2.61 c | 14.51 ± 3.82 f | 105.49 ± 10.04 de | 173.91 ± 15.36 c |
| Phenethyl acetate | 170.96 ± 20.35 c | 1671.16 ± 256.88 a | 322.26 ± 35.56 bc | 501.54 ± 81.34 b | 238.74 ± 1.45 c | 335.12 ± 68.01 bc | 455.92 ± 0.56 b | 214.89 ± 97.86 c | 348.59 ± 14.23 bc | 254.87 ± 60.73 c |
| Citronellyl acetate | n.d | 7.61 ± 1.45 b | 8.53 ± 3.37 b | 6.73 ± 1.38 b | n.d | 2.44 ± 0.17 c | 13.85 ± 0.97 a | n.d | 11.68 ± 1.49 a | n.d |
| Decyl acetate | n.d | n.d | n.d | n.d | n.d | n.d | n.d | n.d | 13.11 ± 2.65 | n.d |
| Ethyl esters | ||||||||||
| Ethyl butyrate | n.d | 3.44 ± 0.4 d | 4.85 ± 0.4 bc | 5.84 ± 0.44 ab | 4.12 ± 0.06 cd | 6.74 ± 0.82 a | 5.91 ± 0.36 ab | n.d | n.d | 6.73 ± 1.25 a |
| Ethyl hexanoate | 62.41 ± 7.664 e | 582.13 ± 76.57 a | 520.42 ± 73.34 ab | 407.55 ± 48.56 bc | 192.75 ± 1.38 d | 368.06 ± 62.20 c | 315.04 ± 160.49 cd | 33.05 ± 1.89 e | 417.24 ± 61.95 bc | 195.36 ± 0.99 d |
| Ethyl heptanoate | 3.97 ± 1.3 d | 5.7 ± 1.45 cd | 3.23 ± 0.38 d | 23.80 ± 8.34 b | 4.56 ± 0.01 d | n.d | 38.36 ± 5.99 a | 12.00 ± 0.56 c | n.d | 2.83 ± 0.45 d |
| Ethyl octanoate | 1763.28 ± 76.86 e | 6750.23 ± 481.42 a | 6347.14 ± 42.91 a | 3996.98 ± 506.14 c | 1794.26 ± 249.20 e | 5085.02 ± 689.79 b | 3028.91 ± 525.23 d | 1427.26 ± 254.62 e | 3376.21 ± 498.56 cd | 1859.77 ± 204.61 e |
| Ethyl nonanoate | 5.62 ± 0.68 b | 1.96 ± 0.17 c | 18.18 ± 0.45 a | 17.35 ± 1.19 a | n.d | 16.93 ± 1.55 a | 4.92 ± 0.29 b | n.d | 5.21 ± 0.45 b | n.d |
| Ethyl 9-decenoate | 1567.7 ± 153.76 a | 1462.57 ± 46.68 a | 1023.39 ± 236.30 b | 732.31 ± 125.88 c | 567.44 ± 110.42 d | 27.91 ± 2.77 e | 45.82 ± 7.6 e | 1054.97 ± 64.15 b | 560.53 ± 71.10 d | 806.55 ± 146.26 c |
| Ethyl decanoate | 484.76 ± 357.86 e | 1006.54 ± 73.59 de | 12156.92 ± 2556.84 a | 3200.49 ± 202.31 c | 2493.39 ± 456.48 cd | 10439.39 ± 2087.73 a | 2612.85 ± 238.33 cd | 320.28 ± 23.88 e | 7693.33 ± 519.71 b | 811.22 ± 99.28 de |
| Ethyl trans-4-decenoate | 12.80 ± 1.94 c | n.d | n.d | n.d | n.d | 1587.57 ± 166.99 b | 3158.02 ± 48.04 a | n.d | n.d | n.d |
| Ethyl E-11-hexadecenoate | n.d | 18.39 ± 0.55 | n.d | n.d | n.d | n.d | n.d | n.d | n.d | 7.51 ± 0.87 |
| Ethyl laurate | 88.08 ± 12.05 de | 843.53 ± 37.61 c | 2490.92 ± 244.30 a | 312.81 ± 21.5 d | 822.51 ± 143.81 c | 2064.04 ± 320.08 b | 228.19 ± 0.78 de | 35.99 ± 4.19 e | n.d | 67.9 ± 3.68 de |
| Ethyl undecanoate | n.d | n.d | n.d | n.d | 128.59 ± 23.89 | n.d | n.d | n.d | n.d | n.d |
| Ethyl myristate | 2.37 ± 0.23 c | 6.29 ± 0.46 bc | 24.57 ± 0.45 a | 3.90 ± 0.67 c | n.d | 11.15 ± 8.79 b | 5.01 ± 0.18 c | n.d | 3.74 ± 0.3 c | n.d |
| Ethyl hexadec-9-enoate | 16.68 ± 1.68 d | 199.43 ± 12.59 a | 46.44 ± 11.86 b | 40.30 ± 3.56 b | 44.74 ± 3.71 b | 38.57 ± 19.42 b | 44.79 ± 5.38 b | 22.69 ± 2.09 cd | 10.93 ± 3.28 d | 33.44 ± 0.27 bc |
| Ethyl hexadecanoate | 6.93 ± 0.95 d | 167.22 ± 9.24 a | 51.05 ± 11.19 b | 19.16 ± 3.03 c | 7.43 ± 0.63 d | 28.47 ± 7.11 c | 6.71 ± 3.52 d | 5.27 ± 0.29 d | 18.74 ± 3.79 c | 7.67 ± 0.88 d |
| Ethyl oleate | n.d | 6.75 ± 1.21 | n.d | n.d | n.d | n.d | n.d | n.d | 2.57 ± 0.43 | n.d |
| Other esters | ||||||||||
| Isobutyl caproate | n.d | n.d | n.d | 1.23 ± 0.35 | n.d | n.d | n.d | n.d | n.d | n.d |
| 3-methylbutyl hexanoate | n.d | n.d | 7.98 ± 0.77 c | 10.26 ± 2.53 c | 1.88 ± 0.19 c | n.d | 12.09 ± 2.22 c | 250.94 ± 43.32 b | 298.38 ± 26.12 a | 0.95 ± 0.43 c |
| Propyl octanoate | 1.89 ± 0.21 c | 2.25 ± 0.39 c | 10.69 ± 1.75 b | n.d | n.d | 13.46 ± 2.64 ab | 14.53 ± 2.56 a | n.d | n.d | n.d |
| 2-methylpropyl octanoate | 10.50 ± 1.34 c | 14.85 ± 1.62 b | 5.92 ± 1.18 de | 8.50 ± 1.81 cd | n.d | 4.48 ± 0.86 e | 19.38 ± 2.99 a | n.d | n.d | n.d |
| 3-methylbutyl octanoate | 22.12 ± 2.48 d | 7.7 ± 0.79 e | 89.52 ± 13.69 a | 39.19 ± 2.01 c | 3.87 ± 0.36 e | 66.99 ± 10.39 b | 36.01 ± 3.46 c | 5.67 ± 0.62 e | 34.11 ± 7.83 c | 4.52 ± 2.04 e |
| Octyl 2-methylbutyrate | n.d | n.d | 19.22 ± 1.37 | 5.24 ± 0.45 | n.d | n.d | n.d | n.d | n.d | n.d |
| Propyl decanoate | n.d | n.d | 18.20 ± 0.36 a | 2.26 ± 0.67 d | n.d | 15.92 ± 1.42 b | n.d | n.d | 8.02 ± 0.68 c | n.d |
| Isobutyl decanoate | n.d | n.d | 7.42 ± 2.16 | 4.24 ± 0.54 | n.d | n.d | n.d | n.d | n.d | n.d |
| 3-methylbutyl decanoate | 1.97 ± 0.16 d | 117.24 ± 23.68 a | 81.58 ± 13.34 b | 33.6 ± 2.42 c | n.d | 20.12 ± 4.57 c | n.d | n.d | 11.95 ± 1.72 d | 1.23 ± 0.09 d |
| Compounds | Threshold (μg/L) | OAV | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| S | CS1 | CS5 | CS10 | PS1 | PS5 | PS10 | WS1 | WS5 | WS10 | ||
| Phenethyl acetate | 250 | 0.68 ± 0.06 c | 6.68 ± 0.59 a | 1.29 ± 0.08 bc | 2.01 ± 0.19 b | 0.95 ± 0.00 c | 1.34 ± 0.19 bc | 1.82 ± 0.00 b | 0.86 ± 0.39 c | 1.39 ± 0.04 bc | 1.02 ± 0.17 c |
| Isoamyl acetate | 30 | 0.83 ± 0.04 f | 26.83 ± 0.77 a | 4.23 ± 0.50 d | 15.21 ± 0.59 b | 2.44 ± 0.05 e | 2.64 ± 0.12 e | 6.22 ± 0.06 c | 0.48 ± 0.13 f | 3.52 ± 0.24 de | 5.80 ± 0.36 c |
| Ethyl octanoate | 5 | 352.66 ± 10.87 e | 1350.05 ± 55.59 a | 1269.43 ± 4.95 a | 799.4 ± 58.44 c | 358.85 ± 35.24 e | 1017.00 ± 97.55 b | 605.78 ± 74.28 d | 285.45 ± 50.92 e | 675.24 ± 70.51 cd | 371.95 ± 28.94 e |
| Ethyl hexanoate | 5 | 12.48 ± 1.08 e | 116.43 ± 8.84 a | 104.08 ± 8.47 ab | 81.51 ± 5.61 bc | 38.55 ± 0.20 d | 73.61 ± 8.80 bc | 63.00 ± 22.70 cd | 6.61 ± 0.38 e | 83.45 ± 8.76 bc | 39.07 ± 0.14 d |
| Ethyl 9-decenoate | 100 | 15.68 ± 1.09 a | 14.63 ± 0.27 a | 10.23 ± 1.36 b | 7.32 ± 0.73 c | 5.67 ± 0.78 d | 0.28 ± 0.02 e | 0.46 ± 0.05 e | 10.55 ± 0.58 b | 5.61 ± 0.50 d | 8.07 ± 1.03 c |
| Ethyl decanoate | 200 | 2.42 ± 1.27 e | 5.03 ± 0.21 de | 60.78 ± 7.38 a | 16.00 ± 0.58 c | 12.47 ± 1.61 cd | 52.20 ± 7.38 a | 13.06 ± 0.84 cd | 1.60 ± 0.12 e | 38.47 ± 1.84 b | 4.06 ± 0.35 de |
| Ethyl laurate | 800 | 0.11 ± 0.01 de | 1.05 ± 0.03 c | 3.11 ± 0.18 a | 0.39 ± 0.02 d | 1.03 ± 0.13 c | 2.58 ± 0.28 b | 0.29 ± 0.00 de | 0.04 ± 0.01 e | n.d | 0.08 ± 0.00 de |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, W.; Li, B.; Chen, Y.; Fu, G.; Fan, H.; Deng, M.; Wan, Y.; Liu, N.; Li, M. Increase the Content of Ester Compounds in Blueberry Wine Fermentation with the Ester-Producing Yeast: Candida glabrata, Pichia anomala, and Wickerhamomyces anomalus. Foods 2022, 11, 3655. https://doi.org/10.3390/foods11223655
Cai W, Li B, Chen Y, Fu G, Fan H, Deng M, Wan Y, Liu N, Li M. Increase the Content of Ester Compounds in Blueberry Wine Fermentation with the Ester-Producing Yeast: Candida glabrata, Pichia anomala, and Wickerhamomyces anomalus. Foods. 2022; 11(22):3655. https://doi.org/10.3390/foods11223655
Chicago/Turabian StyleCai, Wenqin, Bang Li, Yanru Chen, Guiming Fu, Haowei Fan, Mengfei Deng, Yin Wan, Na Liu, and Mengxiang Li. 2022. "Increase the Content of Ester Compounds in Blueberry Wine Fermentation with the Ester-Producing Yeast: Candida glabrata, Pichia anomala, and Wickerhamomyces anomalus" Foods 11, no. 22: 3655. https://doi.org/10.3390/foods11223655
APA StyleCai, W., Li, B., Chen, Y., Fu, G., Fan, H., Deng, M., Wan, Y., Liu, N., & Li, M. (2022). Increase the Content of Ester Compounds in Blueberry Wine Fermentation with the Ester-Producing Yeast: Candida glabrata, Pichia anomala, and Wickerhamomyces anomalus. Foods, 11(22), 3655. https://doi.org/10.3390/foods11223655