
The Influence of Farming Systems, Genotype and Their Interaction on Bioactive Compound, Protein and Starch Content of Bread and Spelt Wheat

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Field Experiment
2.2. Analysis of the Quality Traits and Bioactive Compound, Protein and Starch Content
2.2.1. Physical and Quality Traits of Bread and Spelt Wheat Varieties
2.2.2. Bioactive Compound Content Analysis of Bread and Spelt Wheat Varieties
2.2.3. Protein and Starch Content Analysis of Bread and Spelt Wheat Varieties
2.3. Statistical Analysis
3. Results
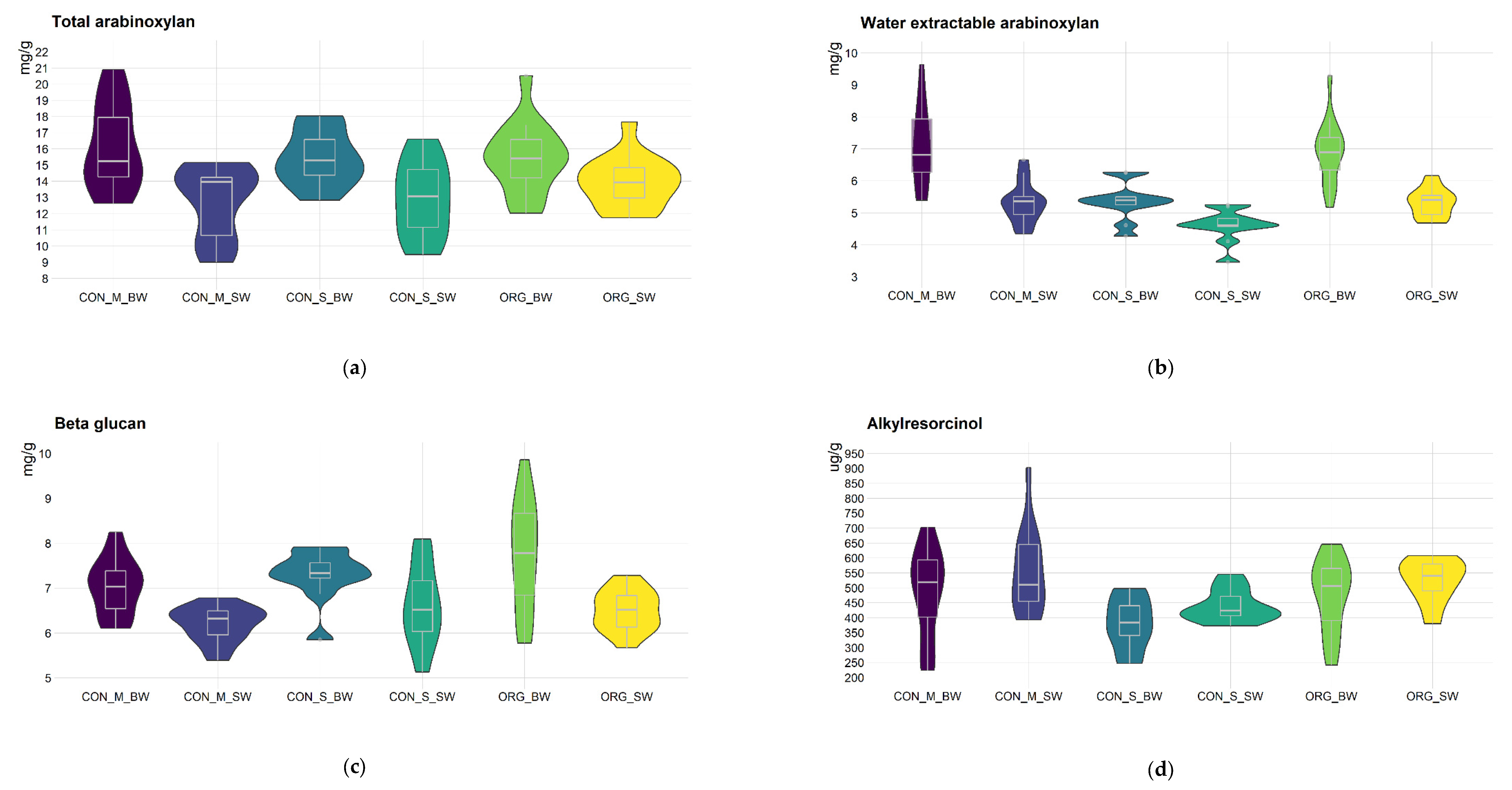
3.1. Differences in Bioactive Compound, Protein and Starch Content between Bread and Spelt Wheat, Conventional and Organic Farming Systems and Serbia and Hungary
3.2. Differences in the Effect of the Variety (Genotype) and the Environment (Farming System and Country) on Bioactive Compound, Protein and Starch Content of Bread and Spelt Wheat
3.3. Correlations of Bioactive Compound, Protein and Starch Content in Bread and Spelt Wheat
3.4. Principal Component Analysis of Bread and Spelt Wheat Grown under Conventional and Organic Farming Systems in Hungary and Serbia
4. Discussion
4.1. Bioactive Compound, Protein and Starch Content and Quality in Bread and Spelt Wheat Species
4.2. The Response of Genotype and the Exceptional Characteristics of the Varieties
4.3. Bioactive Compound, Protein and Starch Content and Quality in Different Farming Systems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kendall, C.W.; Esfahani, A.; Jenkins, D.J. The link between dietary fibre and human health. Food Hydrocoll. 2010, 24, 42–48. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, A.; Costabile, A.; Martin-Pelaez, S.; Vitaglione, P.; Klinder, A.; Gibson, G.R.; Fogliano, V. Potential prebiotic activity of oligosaccharides obtained by enzymatic conversion of durum wheat insoluble dietary fibre into soluble dietary fibre. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M. Prebiotics: The concept revisited. J. Nutr. 2007, 137, 830S–837S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ficco, D.B.M.; Prandi, B.; Amaretti, A.; Anfelli, I.; Leonardi, A.; Raimondi, S.; Pecchioni, N.; De Vita, P.; Faccini, A.; Sforza, S.; et al. Comparison of gluten peptides and potential prebiotic carbohydrates in old and modern Triticum turgidum ssp. genotypes. Int. Food Res. J. 2019, 120, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Shewry, P.R. The HEALTHGRAIN programme opens new opportunities for improving wheat for nutrition and health. Nutr. Bull. 2009, 34, 225–231. [Google Scholar] [CrossRef]
- Sapirstein, H.D. Bioactives in Wheat Bran. In Reference Module in Food Science; Smithers, G.W., Ed.; Elsevier: Amsterdam, The Netherlands, 2016. [Google Scholar] [CrossRef]
- Andersson, A.A.; Andersson, R.; Piironen, V.; Lampi, A.M.; Nyström, L.; Boros, D.; Fraś, A.; Gebruers, K.; Courtin, C.M.; Delcour, J.A.; et al. Contents of dietary fibre components and their relation to associated bioactive components in whole grain wheat samples from the HEALTHGRAIN diversity screen. Food Chem. 2013, 136, 1243–1248. [Google Scholar] [CrossRef]
- Pedrazzani, C.; Vanara, F.; Bhandari, D.R.; Bruni, R.; Spengler, B.; Blandino, M.; Righetti, L. 5-n-Alkylresorcinol Profiles in Different Cultivars of Einkorn, Emmer, Spelt, Common Wheat, and Tritordeum. J. Agric. Food Chem. 2021, 69, 14092–14102. [Google Scholar] [CrossRef]
- Ross, A.B.; Kamal-Eldin, A.; Lundin, E.A.; Zhang, J.X.; Hallmans, G.; Åman, P. Cereal alkylresorcinols are absorbed by humans. J. Nutr. 2003, 133, 2222–2224. [Google Scholar] [CrossRef]
- Sahyoun, N.R.; Jacques, P.F.; Zhang, X.L.; Juan, W.; McKeown, N.M. Whole-grain intake is inversely associated with the metabolic syndrome and mortality in older adults. Am. J. Clin. Nutr. 2006, 83, 124–131. [Google Scholar] [CrossRef] [Green Version]
- Lutsey, P.L.; Jacobs, D.R.; Kori, S.; Mayer-Davis, E.; Shea, S.; Steffen, L.M.; Szklo, M.; Tracy, R. Whole grain intake and its cross-sectional association with obesity, insulin resistance, inflammation, diabetes and subclinical CVD: The MESA Study. Br. J. Nutr. 2007, 98, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, D.R.; Andersen, L.F.; Blomhoff, R. Whole-grain consumption is associated with a reduced risk of noncardiovascular, noncancer death attributed to inflammatory diseases in the Iowa Women’s Health Study. Am. J. Clin. Nutr. 2007, 85, 1606–1614. [Google Scholar] [CrossRef] [Green Version]
- Schatzkin, A.; Park, Y.; Leitzmann, M.F.; Hollenbeck, A.R.; Cross, A.J. Prospective study of dietary fiber, whole grain foods, and small intestinal cancer. Gastroenterology 2008, 135, 1163–1167. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, L.E.O.; Phillips, F.; O’sullivan, K.; Walton, J. Wheat bran: Its composition and benefits to health, a European perspective. Int. J. Food Sci. Nutr. 2012, 63, 1001–1013. [Google Scholar] [CrossRef] [Green Version]
- National Research Council. Understanding agricultural sustainability. In Toward Sustainable Agricultural Systems in the 21st Century, 2nd ed.; Whitacre, P.T., Ed.; National Academies Press: Washington, DC, USA, 2010; pp. 15–39. [Google Scholar]
- Rossi, F.; Bertuzzi, T.; Comizzoli, S.; Turconi, G.; Roggi, C.; Pagani, M.; Craverdi, P.; Pietri, A. Preliminary survey on composition and quality of conventional and organic wheat. Ital. J. Food Sci. 2006, 18, 355–366. [Google Scholar]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/organicag/oa-faq/oa-faq1/en/ (accessed on 8 June 2022).
- Gabriel, D.; Sait, S.M.; Kunin, W.E.; Benton, T.G. Food production vs. biodiversity: Comparing organic and conventional agriculture. J. Appl. Ecol. 2013, 50, 355–364. [Google Scholar] [CrossRef]
- Gomiero, T.; Pimentel, D.; Paoletti, M.G. Environmental impact of different agricultural management practices: Conventional vs. organic agriculture. Crit. Rev. Plant Sci. 2011, 30, 95–124. [Google Scholar] [CrossRef]
- Smith, L.G.; Williams, A.G.; Pearce, B.D. The energy efficiency of organic agriculture: A review. Renew. Agric. Food Syst. 2015, 30, 280–301. [Google Scholar] [CrossRef]
- Cristache, S.E.; Vuță, M.; Marin, E.; Cioacă, S.I.; Vuţă, M. Organic versus conventional farming—A paradigm for the sustainable development of the european countries. Sustainability 2018, 10, 4279. [Google Scholar] [CrossRef] [Green Version]
- Arzani, A.; Ashraf, M. Cultivated ancient wheats (Triticum spp.): A potential source of health-beneficial food products. Compr. Rev. Food Sci. Food Saf. 2017, 16, 477–488. [Google Scholar] [CrossRef]
- Szewczyk, B.F. The influence of morphological features of spelt wheat (Triticum aestivum ssp. spelta) and common wheat (Triticum aestivum ssp. vulgare) varieties on the competitiveness against weeds in organic farming system. J. Food Agric. Environ. 2013, 11, 416–421. [Google Scholar]
- Gebruers, K.; Dornez, E.; Boros, D.; Dynkowska, W.; Bedo, Z.; Rakszegi, M.; Delcour, J.A.; Courtin, C.M. Variation in the content of dietary fiber and components thereof in wheats in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9740–9749. [Google Scholar] [CrossRef] [PubMed]
- Kulathunga, J.; Simsek, S. Dietary fiber variation in ancient and modern wheat species: Einkorn, emmer, spelt and hard red spring wheat. J. Cereal Sci. 2022, 104, 103420. [Google Scholar] [CrossRef]
- Geisslitz, S.; Wieser, H.; Scherf, K.A.; Koehler, P. Gluten protein composition and aggregation properties as predictors for bread volume of common wheat, spelt, durum wheat, emmer and einkorn. J. Cereal Sci. 2018, 83, 204–212. [Google Scholar] [CrossRef]
- MSZ 6367/4-86; Edible, Fodder and Industrial Seeds and Husked Products. Determination of Test Weight, Thousand Kernel Weight and Classification Grade. Hungarian Standards Institution: Budapest, Hungary, 1986.
- ICC 115/1; Method for Using Brabender Farinograph. International Association for Cereal Science and Technology: Vienna, Austria, 1995.
- ICC 137/1; Mechanical Determination of the Wet Gluten Content of Wheat Flour (Glutomatic). International Association for Cereal Science and Technology: Vienna, Austria, 1995.
- ICC 155; Determination of Wet Gluten Quantity and Quality (Gluten Index ac. to Perten) of Whole Wheat Meal and Wheat Flour (Triticum aestivum). International Association for Cereal Science and Technology: Vienna, Austria, 1995.
- Tömösközi, S.; Nádosi, M.; Balázs, G.; Cavanagh, C.; Morgunov, A.; Salgó, A.; Békés, F. Revival of sedimentation value—Method development, quality prediction and molecular background. In Proceedings of the 10th International Gluten Workshop, INRA, Clermont-Ferrand, France, 7–9 September 2009; Branlard, G., Ed.; pp. 104–108. [Google Scholar]
- ICC 116/1; Determination of the Sedimentation Value (according to Zeleny) as an Approximate Measure of Baking Quality. International Association for Cereal Science and Technology: Vienna, Austria, 1997.
- Douglas, S.G. A rapid method for the determination of pentosans in wheat flour. Food Chem. 1981, 7, 139–145. [Google Scholar] [CrossRef]
- Approved Methods 32–23.01; Beta-Glucan Content of Barley and Oats–Rapid Enzymatic Procedure. AACC International: Saint Paul, MN, USA, 1995.
- Tłuścik, F.; Kazubek, A.; Mejbaum-Katzenellenbogen, W. Alkylresorcinols in rye (Secale cereale L.) grains. VI. Colorimetric micromethod for the determination of alkylresorcinols with the use of diazonium salt, Fast Blue B. Acta Soc. Bot. Pol. 1981, 50, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Approved Methods 32–32.01; Measurement of Total Fructan in Foods by an Enzymatic/Spectrophotometric Method. AACC International: Saint Paul, MN, USA, 1995.
- International Association for Cereal Science and Technology. ICC 167; Determination of crude protein in grain and grain products for food and feed by the Dumas combustion principle. International Association for Cereal Science and Technology: Vienna, Austria, 1995.
- ICC 202; Procedure for Near Infrared (NIR) Reflectance Analysis of Ground Wheat and Milled Wheat Products. International Association for Cereal Science and Technology: Vienna, Austria, 1995.
- American Association of Cereal Chemists. General Pasting Method for Wheat or Rye Flour of Starch Using the Rapid Visco Analyser, 8th ed.; Approved Methods of the AACC, Method 76-21, Starch; American Association of Cereal Chemists: Saint Paul, MN, USA, 1991. [Google Scholar]
- Batey, I.L.; Curtin, B.M.; Moore, S.A. Optimization of rapid-visco analyser test conditions for predicting Asian noodle quality. Cereal Chem. 1997, 74, 497–501. [Google Scholar] [CrossRef]
- ICC 172; Flour from Wheat (Triticum aestivum)—Determination of Damaged Starch by an Amperometric Method Using the SDMatic, 2007. International Association for Cereal Science and Technology: Vienna, Austria, 1995.
- Longin, C.F.H.; Würschum, T. Back to the future–tapping into ancient grains for food diversity. Trends Plant Sci. 2016, 21, 731–737. [Google Scholar] [CrossRef]
- Sugár, E.; Fodor, N.; Sándor, R.; Bónis, P.; Vida, G.; Árendás, T. Spelt wheat: An alternative for sustainable plant production at low N-levels. Sustainability 2019, 11, 6726. [Google Scholar] [CrossRef] [Green Version]
- Escarnot, E.; Dornez, E.; Verspreet, J.; Agneessens, R.; Courtin, C.M. Quantification and visualization of dietary fibre components in spelt and wheat kernels. J. Cereal Sci. 2015, 62, 124–133. [Google Scholar] [CrossRef]
- Escarnot, E.; Agneessens, R.; Wathelet, B.; Paquot, M. Quantitative and qualitative study of spelt and wheat fibres in varying milling fractions. Food Chem. 2010, 122, 857–863. [Google Scholar] [CrossRef]
- Suchowilska, E.; Wiwart, M.; Krska, R.; Kandler, W. Do Triticum aestivum L. and Triticum spelta L. Hybrids Constitute a Promising Source Material for Quality Breeding of New Wheat Varieties? Agronomy 2019, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Kulathunga, J.; Reuhs, B.L.; Zwinger, S.; Simsek, S. Comparative study on kernel quality and chemical composition of ancient and modern wheat species: Einkorn, emmer, spelt and hard red spring wheat. Foods 2021, 10, 761. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S. Do “ancient” wheat species differ from modern bread wheat in their contents of bioactive components? J. Cereal Sci. 2015, 65, 236–243. [Google Scholar] [CrossRef] [Green Version]
- Barron, C.; Surget, A.; Rouau, X. Relative amounts of tissues in mature wheat (Triticum aestivum L.) grain and their carbohydrate and phenolic acid composition. J. Cereal Sci. 2007, 45, 88–96. [Google Scholar] [CrossRef]
- Antoine, C.; Peyron, S.; Mabille, F.; Lapierre, C.; Bouchet, B.; Abecassis, J.; Rouau, X. Individual contribution of grain outer layers and their cell wall structure to the mechanical properties of wheat bran. J. Agric. Food Chem. 2003, 51, 2026–2033. [Google Scholar] [CrossRef]
- Tian, X.; Wang, Z.; Yang, S.; Wang, X.; Li, L.; Sun, B.; Ma, S.; Zheng, S. Microstructure observation of multilayers separated from wheat bran. GOST 2021, 4, 165–173. [Google Scholar] [CrossRef]
- Brouns, F.; Hemery, Y.; Price, R.; Anson, N.M. Wheat aleurone: Separation, composition, health aspects, and potential food use. Crit. Rev. Food Sci. Nutr. 2012, 52, 553–568. [Google Scholar] [CrossRef] [Green Version]
- Izydorczyk, M.S.; Symons, S.J.; Dexter, J.E. Fractionation of wheat and barley. In Whole Grain Foods in Health and Disease, 1st ed.; Marquart, L., Slavin, J.L., Fulcher, G.R., Eds.; American Association of Cereal Chemists: Eagan, MN, USA, 2002; pp. 47–82. [Google Scholar]
- Bojňanská, T.; Frančáková, H. The use of spelt wheat (Triticum spelta L.) for baking applications. Rost Vyroba 2002, 48, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Marconi, E.; Carcea, M.; Schiavone, M.; Cubadda, R. Spelt (Triticum spelta L.) pasta quality: Combined effect of flour properties and drying conditions. Cereal Chem. 2002, 79, 634–639. [Google Scholar] [CrossRef]
- Świeca, M.; Dziki, D.; Gawlik-Dziki, U.; Różyło, R.; Andruszczak, S.; Kraska, P.; Kowalczyk, D.; Pałys, E.; Baraniak, B. Grinding and nutritional properties of six spelt (Triticum aestivum ssp. spelta L.) cultivars. Cereal Chem. 2014, 91, 247–254. [Google Scholar] [CrossRef]
- Denčić, S.; Obreht, D.; Kobiljski, B.; Štatkić, S.; Bede, B. Genetic determination of breadmaking quality in wheat. In Proceedings of the 43rd Croatian and 3rd International Symposium on Agriculture, Zagreb, Croatia, 18–21 February 2008; pp. 278–281. [Google Scholar]
- Geneix, N.; Dalgalarrondo, M.; Tassy, C.; Nadaud, I.; Barret, P.; Bakan, B.; Marion, D. Relationships between puroindoline A-prolamin interactions and wheat grain hardness. PLoS ONE 2020, 15, e0225293. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.U.; Steingass, C.B.; Longin, C.F.H.; Würschum, T.; Carle, R.; Schweiggert, R.M. Alkylresorcinol composition allows the differentiation of Triticum spp. having different degrees of ploidy. J. Cereal Sci. 2015, 65, 244–251. [Google Scholar] [CrossRef]
- Andersson, A.A.; Kamal-Eldin, A.; Fras, A.; Boros, D.; Åman, P. Alkylresorcinols in wheat varieties in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2008, 56, 9722–9725. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, J.U.; Schweiggert, R.M.; Würschum, T.; Longin, C.F.H.; Carle, R. Lipophilic antioxidants in wheat (Triticum spp.): A target for breeding new varieties for future functional cereal products. J. Funct. Foods. 2016, 20, 594–605. [Google Scholar] [CrossRef]
- Wang, J.; Chatzidimitriou, E.; Wood, L.; Hasanalieva, G.; Markellou, E.; Iversen, P.O.; Seal, C.; Baranski, M.; Vigar, V.; Ernst, L.; et al. Effect of wheat species (Triticum aestivum vs T. spelta), farming system (organic vs conventional) and flour type (wholegrain vs white) on composition of wheat flour—Results of a retail survey in the UK and Germany—2. Antioxidant activity, and phenolic and mineral content. Food Chem. 2020, 6, 100091. [Google Scholar] [CrossRef]
- Laguerre, M.; López Giraldo, L.J.; Lecomte, J.; Figueroa-Espinoza, M.C.; Barea, B.; Weiss, J.; Decker, E.A.; Villeneuve, P. Chain length affects antioxidant properties of chlorogenate esters in emulsion: The cutoff theory behind the polar paradox. J. Agric. Food Chem. 2009, 57, 11335–11342. [Google Scholar] [CrossRef]
- Suzuki, Y.; Kurano, M.; Esumi, Y.; Yamaguchi, I.; Doi, Y. Biosynthesis of 5-alkylresorcinol in rice: Incorporation of a putative fatty acid unit in the 5-alkylresorcinol carbon chain. Bioorg. Chem. 2003, 31, 437–452. [Google Scholar] [CrossRef]
- Elder, A.S.; Coupland, J.N.; Elias, R.J. Effect of alkyl chain length on the antioxidant activity of alkylresorcinol homologues in bulk oils and oil-in-water emulsions. Food Chem. 2021, 346, 128885. [Google Scholar] [CrossRef]
- Gebruers, K.; Dornez, E.; Bedo, Z.; Rakszegi, M.; Fras, A.; Boros, D.; Courtin, C.M.; Delcour, J.A. Environment and genotype effects on the content of dietary fiber and its components in wheat in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2010, 58, 9353–9361. [Google Scholar] [CrossRef]
- Brandolini, A.; Hidalgo, A.; Moscaritolo, S. Chemical composition and pasting properties of einkorn (Triticum monococcum L. subsp. monococcum) whole meal flour. J. Cereal Sci. 2008, 47, 599–609. [Google Scholar] [CrossRef]
- Zieliński, H.; Ceglińska, A.; Michalska, A. Bioactive compounds in spelt bread. Eur. Food Res. Technol. 2008, 226, 537–544. [Google Scholar] [CrossRef]
- Takač, V.; Tóth, V.; Rakszegi, M.; Mikić, S.; Mirosavljević, M.; Kondić-Špika, A. Differences in processing quality traits, protein content and composition between spelt and bread wheat genotypes grown under conventional and organic production. Foods 2021, 10, 156. [Google Scholar] [CrossRef]
- Call, L.; Kapeller, M.; Grausgruber, H.; Reiter, E.; Schoenlechner, R.; D’Amico, S. Effects of species and breeding on wheat protein composition. J. Cereal Sci. 2020, 93, 102974. [Google Scholar] [CrossRef]
- Dziki, D.; Laskowski, J. Wheat kernel physical properties and milling process. Acta Geophys. 2005, 6, 59–71. [Google Scholar]
- Markowski, M.; Żuk-Gołaszewska, K.; Kwiatkowski, D. Influence of variety on selected physical and mechanical properties of wheat. Ind. Crops Prod. 2013, 47, 113–117. [Google Scholar] [CrossRef]
- Longin, C.F.H.; Ziegler, J.; Schweiggert, R.; Koehler, P.; Carle, R.; Würschum, T. Comparative study of hulled (einkorn, emmer, and spelt) and naked wheats (durum and bread wheat): Agronomic performance and quality traits. Crop Sci. 2016, 56, 302–311. [Google Scholar] [CrossRef]
- Ratajczak, K.; Sulewska, H.; Grażyna, S.; Matysik, P. Agronomic traits and grain quality of selected spelt wheat varieties versus common wheat. J. Crop Improv. 2020, 34, 654–675. [Google Scholar] [CrossRef]
- Biel, W.; Stankowski, S.; Sobolewska, M.; Sadkiewicz, J.; Jaroszewska, A.; Pużyński, S. Effect of selected agronomic factors on the baking quality of winter spelt strains and cultivars (Triticum aestivum ssp. spelta L.) in comparison with common wheat (Triticum aestivum ssp. vulgare). Rom. Agric. Res. 2016, 33, 1–8. [Google Scholar]
- Petrenko, V.; Spychaj, R.; Prysiazhniuk, O.; Sheiko, T.; Khudolii, L. Evaluation of three wheat species (Triticum aestivum L., T. spelta L., T. dicoccum (Schrank) Schuebl) commonly used in organic cropping systems, considering selected parameters of technological quality. Rom. Agric. Res. 2018, 35, 255–264. [Google Scholar]
- Schober, T.J.; Bean, S.R.; Kuhn, M. Gluten proteins from spelt (Triticum aestivum ssp. spelta) cultivars: A rheological and size-exclusion high-performance liquid chromatography study. J. Cereal Sci. 2006, 44, 161–173. [Google Scholar] [CrossRef]
- Pruska-Kedzior, A.; Kedzior, Z.; Klockiewicz-Kaminska, E. Comparison of viscoelastic properties of gluten from spelt and common wheat. Eur. Food Res. Technol. 2008, 227, 199–207. [Google Scholar] [CrossRef]
- Sobczyk, A.; Pycia, K.; Stankowski, S.; Jaworska, G.; Kuźniar, P. Evaluation of the rheological properties of dough and quality of bread made with the flour obtained from old cultivars and modern breeding lines of spelt (Triticum aestivum ssp. spelta). J. Cereal Sci. 2017, 77, 35–41. [Google Scholar] [CrossRef]
- Vida, G.; Szunics, L.; Veisz, O.; Bedő, Z.; Láng, L.; Árendás, T.; Bónis, P.; Rakszegi, M. Effect of genotypic, meteorological and agronomic factors on the gluten index of winter durum wheat. Euphytica 2014, 197, 61–71. [Google Scholar] [CrossRef]
- Podolska, G.; Aleksandrowicz, E.; Szafrańska, A. Bread making potential of Triticum aestivum and Triticum spelta species. Open Life Sci. 2020, 15, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Aydoğan, S.; Şahin, M.; Göçmen Akçacik, A.; Hamzaoğlu, S.; Taner, S. Relationships between farinograph parameters and bread volume, physicochemical traits in bread wheat flours. J. Bahri Dagdas Crop Res. 2015, 3, 14–18. [Google Scholar]
- Denčić, S.; Mladenov, N.; Kobiljski, B. Effects of genotype and environment on breadmaking quality in wheat. Int. J. Plant Prod. 2011, 5, 71–82. [Google Scholar]
- Carvajal-Millan, E.; Rascón-Chu, A.; Márquez-Escalante, J.A.; Micard, V.; de León, N.P.; Gardea, A. Maize bran gum: Extraction, characterization and functional properties. Carbohydr. Polym. 2007, 69, 280–285. [Google Scholar] [CrossRef]
- Nino-Medina, G.; Carvajal-Millán, E.; Rascon-Chu, A.; Marquez-Escalante, J.A.; Guerrero, V.; Salas-Munoz, E. Feruloylated arabinoxylans and arabinoxylan gels: Structure, sources and applications. Phytochem. Rev. 2010, 9, 111–120. [Google Scholar] [CrossRef]
- Biliaderis, C.G.; Izydorczyk, M.S.; Rattan, O. Effect of arabinoxylans on bread-making quality of wheat flours. Food Chem. 1995, 53, 165–171. [Google Scholar] [CrossRef]
- Gallant, D.J.; Bouchet, B.; Baldwin, P.M. Microscopy of starch: Evidence of a new level of granule organization. Carbohydr. Polym. 1997, 32, 177–191. [Google Scholar] [CrossRef]
- Fiedorowicz, G.K.M. Comparison of molecular structure and selected physicochemical properties of spelt wheat and common wheat starches. J. Food Nutr. Res. 2014, 53, 31–38. [Google Scholar]
- Lindeboom, N.; Chang, P.R.; Tyler, R.T. Analytical, biochemical and physicochemical aspects of starch granule size, with emphasis on small granule starches: A review. Starch-Stärke 2004, 56, 89–99. [Google Scholar] [CrossRef]
- Finnie, S.M.; Bettge, A.D.; Morris, C.F. Influence of cultivar and environment on water-soluble and water-insoluble arabinoxylans in soft wheat. Cereal Chem. 2006, 83, 617–623. [Google Scholar] [CrossRef]
- Li, S.; Morris, C.F.; Bettge, A.D. Genotype and environment variation for arabinoxylans in hard winter and spring wheats of the US Pacific Northwest. Cereal Chem. 2009, 86, 88–95. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.L.; Huynh, B.L.; Wallwork, H.; Stangoulis, J. Identification of quantitative trait loci for grain arabinoxylan concentration in bread wheat. Crop Sci. 2011, 51, 1143–1150. [Google Scholar] [CrossRef]
- Martinant, J.P.; Billot, A.; Bouguennec, A.; Charmet, G.; Saulnier, L.; Branlard, G. Genetic and environmental variations in water-extractable arabinoxylans content and flour extract viscosity. J. Cereal Sci. 1999, 30, 45–48. [Google Scholar] [CrossRef]
- Dornez, E.; Gebruers, K.; Joye, I.J.; De Ketelaere, B.; Lenartz, J.; Massaux, C.; Bodson, B.; Delcour, J.A.; Courtin, C.M. Effects of genotype, harvest year and genotype-by-harvest year interactions on arabinoxylan, endoxylanase activity and endoxylanase inhibitor levels in wheat kernels. J. Cereal Sci. 2008, 47, 180–189. [Google Scholar] [CrossRef]
- Ciccoritti, R.; Scalfati, G.; Cammerata, A.; Sgrulletta, D. Variations in content and extractability of durum wheat (Triticum turgidum L. var durum) Arabinoxylans associated with genetic and environmental factors. Int. J. Mol. Sci. 2011, 12, 4536–4549. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Piironen, V.; Lampi, A.M.; Edelmann, M.; Kariluoto, S.; Nurmi, T.; Fernandez-Orozco, R.; Ravel, C.; Charmet, G.; Andersson, A.A.M.; et al. The HEALTHGRAIN wheat diversity screen: Effects of genotype and environment on phytochemicals and dietary fiber components. J. Agric. Food Chem. 2010, 58, 9291–9298. [Google Scholar] [CrossRef]
- Huynh, B.L.; Palmer, L.; Mather, D.E.; Wallwork, H.; Graham, R.D.; Welch, R.M.; Stangoulis, J.C. Genotypic variation in wheat grain fructan content revealed by a simplified HPLC method. J. Cereal Sci. 2008, 48, 369–378. [Google Scholar] [CrossRef]
- Andersson, A.A.; Kamal-Eldin, A.; Åman, P. Effects of environment and variety on alkylresorcinols in wheat in the HEALTHGRAIN diversity screen. J. Agric. Food Chem. 2010, 58, 9299–9305. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, U.M.; Murat, F.; Abrouk, M.; Pont, C.; Confolent, C.; Oury, F.X.; Ward, J.; Boros, D.; Gebruers, K.; Delcour, J.A.; et al. Combined meta-genomics analyses unravel candidate genes for the grain dietary fiber content in bread wheat (Triticum aestivum L.). Funct. Integr. Genom. 2011, 11, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Li, Y.; Zhang, H.; Yan, S.; Li, W. Effects of irrigation schemes on the characteristics of starch and protein in wheat (Triticum aestivum L.). Starch-Stärke 2016, 68, 454–461. [Google Scholar] [CrossRef]
- Nhan, M.T.; Copeland, L. Effects of growing environment on properties of starch from five Australian wheat varieties. Cereal Chem. 2014, 91, 587–594. [Google Scholar] [CrossRef]
- Labuschagne, M.T.; Geleta, N.; Osthoff, G. The influence of environment on starch content and amylose to amylopectin ratio in wheat. Starch-Stärke 2007, 59, 234–238. [Google Scholar] [CrossRef]
- Rhazi, L.; Méléard, B.; Daaloul, O.; Grignon, G.; Branlard, G.; Aussenac, T. Genetic and environmental variation in starch content, starch granule distribution and starch polymer molecular characteristics of French bread wheat. Foods 2021, 10, 205. [Google Scholar] [CrossRef]
- Hu, E.A.; Pan, A.; Malik, V.; Sun, Q. White rice consumption and risk of type 2 diabetes: Meta-analysis and systematic review. Br. Med. J. 2012, 344, e1454. [Google Scholar] [CrossRef] [Green Version]
- Korycińska, M.; Czelna, K.; Jaromin, A.; Kozubek, A. Antioxidant activity of rye bran alkylresorcinols and extracts from whole-grain cereal products. Food Chem. 2009, 116, 1013–1018. [Google Scholar] [CrossRef]
- Biel, W.; Stankowski, S.; Jaroszewska, A.; Pużyński, S.; Bośko, P. The influence of selected agronomic factors on the chemical composition of spelt wheat (Triticum aestivum ssp. spelta L.) grain. J. Integr. Agric. 2016, 15, 1763–1769. [Google Scholar] [CrossRef] [Green Version]
- Bonafaccia, G.; Galli, V.; Francisci, R.; Mair, V.; Skrabanja, V.; Kreft, I. Characteristics of spelt wheat products and nutritional value of spelt wheat-based bread. Food Chem. 2000, 68, 437–441. [Google Scholar] [CrossRef]
- Menkovska, M.; Levkov, V.; Damjanovski, D.; Grojgovska, N.; Knezevic, D.; Nikolova, N.; Andreevska, D. Content of TDF, SDF and IDF in cereals grown by organic and conventional farming—A short report. Polish J. Food Nutr. Sci. 2017, 67, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Park, E.Y.; Baik, B.K.; Miller, P.R.; Burke, I.C.; Wegner, E.A.; Tautges, N.E.; Craig, F.; Morris, E.; Fuerst, E.P. Functional and nutritional characteristics of wheat grown in organic and conventional cropping systems. Cereal Chem. 2015, 92, 504–512. [Google Scholar] [CrossRef]
- Čukelj, N.; Ajredini, S.; Krpan, M.; Novotni, D.; Voučko, B.; Vrana Špoljarić, I.; Hruškar, M.; Ćurić, D. Bioactives in organic and conventional milled cereal products from Croatian market. Croat. J. Food Technol. Biotechnol. Nutr. 2015, 10, 23–30. [Google Scholar]
- Nocente, F.; De Stefanis, E.; Ciccoritti, R.; Pucciarmati, S.; Taddei, F.; Campiglia, E.; Radicetti, E.; Mancinelli, R. How do conventional and organic management affect the healthy potential of durum wheat grain and semolina pasta traits? Food Chem. 2019, 297, 124884. [Google Scholar] [CrossRef]
- Langenkämper, G.; Zorb, C.; Seifert, M.; Mader, P.; Fretzdorff, B.; Betsche, T. Nutritional quality of organic and conventional wheat. J. Appl. Bot. Food Qual. 2006, 80, 150. [Google Scholar]
- Menkovska, M.; Damjanovski, D.; Levkov, V.; Gjorgovska, N.; Knezevic, D.; Nikolova, N.; Stanoev, V. Content of B-glucan in cereals grown by organic and conventional farming. Banat’s. J. Biotechnol. 2017, 8, 39–47. [Google Scholar] [CrossRef]
- Pandino, G.; Mattiolo, E.; Lombardo, S.; Lombardo, G.M.; Mauromicale, G. Organic cropping system affects grain chemical composition, rheological and agronomic performance of durum wheat. Agriculture 2020, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Menkovska, M.; Brankovic, G.; Pajic, V.; Momirovic, G.S.; Knezevic, D. Associations between nutritional composition and farming type of organically and conventionally grown cereals. Nat. Sci. Discov. 2015, 1, 97–101. [Google Scholar] [CrossRef]
- Dickin, E.; Steele, K.; Frost, G.; Edwards-Jones, G.; Wright, D. Effect of genotype, environment and agronomic management on β-glucan concentration of naked barley grain intended for health food use. J. Cereal Sci. 2011, 54, 44–52. [Google Scholar] [CrossRef]
- Güler, M. The effect of different nitrogen and irrigation levels on grain β-glucan content in some durum wheat cultivars. Turk. J. Agric. For. 2010, 34, 101–108. [Google Scholar] [CrossRef]
- Mazzoncini, M.; Antichi, D.; Silvestri, N.; Ciantelli, G.; Sgherri, C. Organically vs conventionally grown winter wheat: Effects on grain yield, technological quality, and on phenolic composition and antioxidant properties of bran and refined flour. Food Chem. 2015, 175, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Baranski, M.; Korkut, R.; Kalee, H.A.; Wood, L.; Bilsborrow, P.; Janovska, D.; Leifert, A.; Winter, S.; Willson, A.; et al. Performance of modern and traditional spelt wheat (Triticum spelta) varieties in rain-fed and irrigated, organic and conventional production systems in a semi-arid environment; results from exploratory field experiments in Crete, Greece. Agronomy 2021, 11, 890. [Google Scholar] [CrossRef]
- Vicente, R.; Martínez-Carrasco, R.; Pérez, P.; Morcuende, R. New insights into the impacts of elevated CO2, nitrogen, and temperature levels on the regulation of C and N metabolism in durum wheat using network analysis. New Biotechnol. 2018, 40, 192–199. [Google Scholar] [CrossRef]
- Morcuende, R.; Kostadinova, S.; Pérez, P.; Del Molino, I.M.; Martínez-Carrasco, R. Nitrate is a negative signal for fructan synthesis, and the fructosyltransferase-inducing trehalose inhibits nitrogen and carbon assimilation in excised barley leaves. New Phytol. 2004, 161, 749–759. [Google Scholar] [CrossRef] [Green Version]
- Rakszegi, M.; Lovegrove, A.; Balla, K.; Láng, L.; Bedő, Z.; Veisz, O.; Shewry, P.R. Effect of heat and drought stress on the structure and composition of arabinoxylan and β-glucan in wheat grain. Carbohydr. Polym. 2014, 102, 557–565. [Google Scholar] [CrossRef]
- Gagliardi, A.; Carucci, F.; Masci, S.; Flagella, Z.; Gatta, G.; Giuliani, M.M. Effects of genotype, growing season and nitrogen level on gluten protein assembly of durum wheat grown under mediterranean conditions. Agronomy 2020, 10, 755. [Google Scholar] [CrossRef]
- Liu, P.; Guo, W.; Jiang, Z.; Pu, H.; Feng, C.; Zhu, X.; Peng, Y.; Kuang, A.; Little, C.R. Effects of high temperature after anthesis on starch granules in grains of wheat (Triticum aestivum L.). J. Agric. Sci. 2011, 149, 159–169. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Variety | Pedigree | Origin | Year |
|---|---|---|---|
| T. aestivum L. subsp. aestivum | |||
| Apache | Axial/NRPB-84-4233 | FRA | 1998 |
| Balkan | Bačka/Bez1//Miron808/3/NS433/4/Skor35 | SRB | 1979 |
| Estevan | Capo/SE-24090 | DEU | 2009 |
| Pobeda | Sremica/Balkan | SRB | 1990 |
| Recital | Mexique-267(R-267)/9369 | FRA | 1986 |
| T. aestivum L. subsp. spelta | |||
| Baulander Spelz | Geiberger Spelz | DEU | 1926 |
| Ostro | Oberkulmer-Rotkorn/Steins-Roter-Tiroler | CHE | 1978 |
| Rouquin | Lignée-24/Ardenne//Altgol | BEL | 1979 |
| Schwabenkorn | (S)LV | DEU | 1988 |
| Oberkulmer-Rotkorn | (S)LV-CHE | CHE | 1948 |
| Developing Conditions | Hungary | Serbia | |
|---|---|---|---|
| Position | geographic coordinates | 47°18′ N, | 45°20′ N, |
| 18°47′ E | 19°51′ E | ||
| Altitude (m) | 115 | 84 | |
| Developing parameters | previous crop: conventional/organic | 2019/20: facelia/facelia 2020/21: oil radish/buckwheat 2021/22: oil radish/buckwheat | 2018/19: soybean 2019/20: soybean |
| sowing density wheat/spelt (seed/m2) | 500/280 | 550 | |
| Soil parameters | soil type | chernozem | chernozem |
| pH (KCl) | 7.25 | 7.41 | |
| humus (m/m%) | 2.8 | 2.6 | |
| P2O5 (mg/kg) | 210 | 208 | |
| K2O (mg/kg) | 210 | 176 | |
| average nitrogen input through nitrogen, phosphorus and potassium (NPK) fertilizer (conventional) per year (active ingredient, kg/ha) | 120 | 100 | |
| 2018/2019 | 2019/2020 | 2020/2021 | ||||
|---|---|---|---|---|---|---|
| Hungary | Serbia | Hungary | Serbia | Hungary | ||
| full season | Growing period (days) | 279 | 264 | 288 | 274 | 288 |
| Cumulative precipitation (mm) | 365.6 | 435.3 | 382.3 | 473.5 | 300.5 | |
| Mean temperature (°C) | 9.3 | 11.5 | 9.48 | 11.6 | 8.81 | |
| Absolute min temperature (°C) | −14.4 | −16.3 | −8.8 | −6.6 | −11.6 | |
| Absolute max temperature (°C) | 36.0 | 35.0 | 33.3 | 34.4 | 37.4 | |
| last 100 days | Cum. precipitation in the last 100 days before harvest (mm) | 225.0 | 265.4 | 132.5 | 226.5 | 116.1 |
| Mean temperature in the last 100 days (°C) | 17.1 | 17.5 | 16.3 | 18.7 | 16.3 | |
| Absolute min temp in the last 100 days (°C) | −0.7 | −0.9 | −3.6 | −5.4 | −6.5 | |
| Absolute max temp in the last 100 days (°C) | 36.0 | 35.0 | 33.3 | 34.4 | 37.4 | |
| abs. min–max | No of days with Tmin ≤ 0 °C | 90 | 75 | 90 | 52 | 85 |
| No of days with Tmin ≤ −10 °C | 6 | 3 | 0 | 0 | 3 | |
| No of days with Tmax ≥ 25 °C | 42 | 38 | 36 | 45 | 40 | |
| No of days with Tmax ≥ 30 °C | 16 | 17 | 6 | 8 | 18 | |
| No of days with Tmax ≥ 35 °C | 1 | 1 | 0 | 0 | 1 | |
| Total | Water Extractable | ||||||
|---|---|---|---|---|---|---|---|
| Trait | Arabinoxylan | Arabinoxylan | β-Glucan | Alkylresorcinol | Fructan | Protein | Starch |
| mg/g | mg/g | mg/g | µg/g | % | % | % | |
| Species | <0.0001 *** | <0.0001 *** | <0.0001 *** | 0.004 ** | <0.0001 *** | <0.0001 *** | <0.0001 *** |
| T. aestivum L. subsp. aestivum | |||||||
| Variety | <0.0001 *** | <0.0001 *** | 0.0300 * | <0.0001 *** | <0.0001 *** | 0.0020 ** | <0.0001 *** |
| Apache | 14.73 a,b | 5.66 a | 7.13 a,b | 509.68 c,d | 1.01 a,b | 10.81 a | 58.81 b |
| Balkan | 16.08 b,c | 6.81 b | 7.64 a,b | 406.17 a,b | 1.08 b | 12.56 b | 57.81 b |
| Estevan | 13.49 a | 5.87 a | 7.27 a,b | 547.18 d | 0.80 a | 13.55 c | 56.36 a |
| Pobeda | 16.21 c | 6.73 b | 7.84 b | 366.29 a | 1.23 b | 12.35 b | 57.99 b |
| Recital | 18.00 d | 7.83 c | 6.87 a | 443.62 b,c | 1.20 b | 10.78 a | 61.93 c |
| Average | 15.70 B | 6.58 B | 7.36 B | 454.87 A | 1.06 B | 12.04 A | 58.49 B |
| Growing season | <0.0001 *** | <0.0001 *** | 0.0020 ** | <0.0001 *** | 0.0770 n.s. | 0.0010 *** | 0.0770 n.s. |
| 2018/2019 | 14.96 a | 6.18 a | 7.75 b | 372.18 a | 1.04 a | 12.87 c | 57.82 a |
| 2019/2020 | 17.12 b | 6.19 a | 7.34 a,b | 457.75 b | 1.14 a | 12.00 b | 58.23 a |
| 2020/2021 | 14.69 a | 7.78 b | 6.82 a | 574.87 c | 0.98 a | 10.86 a | 59.88 b |
| Farming system | 0.7980 n.s. | <0.0001 *** | 0.0140 * | 0.0120 * | <0.0001 *** | <0.0001 *** | <0.0001 *** |
| Hungary | |||||||
| Conventional | 16.12 a | 7.05 b | 7.03 a | 483.56 b | 0.92 a | 13.72 c | 56.26 a |
| Organic | 15.44 a | 6.92 b | 7.73 b | 477.01 b | 1.16 b | 10.54 a | 59.39 b |
| Serbia | |||||||
| Conventional | 15.46 a | 5.36 a | 7.27 a,b | 381.48 a | 1.10 a,b | 11.93 b | 60.28 b |
| T. aestivum L. subsp. spelta | |||||||
| Variety | 0.2560 n.s. | 0.6410 n.s. | <0.0001 *** | 0.0550 n.s. | <0.0001 *** | 0.1220 n.s. | 0.4340 n.s. |
| Baulander Spelz | 12.73 a | 5.10 a | 4.98 a | 454.93 a | 0.91 b | 14.57 b,c | 55.86 a,b |
| Ostro | 13.08 a | 5.19 a | 6.29 b | 515.16 a,b | 1.00 b | 15.11 c | 54.96 a |
| Rouquin | 13.84 a | 4.99 a | 5.57 a,b | 582.54 b | 0.69 a | 13.66 a,b | 55.19 a,b |
| Schwabenkorn | 12.58 a | 5.17 a | 5.89 a,b | 492.63 a,b | 0.87 a,b | 13.25 a | 56.66 b |
| Oberkulmer-Rotkorn | 14.00 a | 5.22 a | 6.67 b | 514.47 a,b | 0.88 b | 15.05 c | 54.64 a |
| Average | 13.25 A | 5.13 A | 5.77 A | 511.62 B | 0.87 A | 14.33 B | 55.45 A |
| Growing season | <0.0001 *** | <0.0001 *** | 0.2200 n.s. | <0.0001 *** | 0.1230 n.s. | <0.0001 *** | <0.0001 *** |
| 2018/2019 | 12.50 a | 4.92 a | 5.13 a | 542.48 b | 0.82 a | 15.25 b | 54.94 b |
| 2019/2020 | 14.52 b | 4.94 a | 6.42 b | 440.13 a | 0.91 a | 15.64 b | 53.26 a |
| 2020/2021 | 12.46 a | 5.74 b | 6.21 b | 596.49 b | 0.89 a | 10.98 a | 59.29 c |
| Farming system | 0.0110 * | <0.0001 *** | 0.0430 * | 0.0010 *** | 0.3950 n.s. | 0.0010 *** | 0.0050 ** |
| Hungary | |||||||
| Conventional | 12.74 a | 5.31 b | 6.17 b | 553.26 b | 0.84 a | 15.06 b | 53.94 a |
| Organic | 13.97 b | 5.31 b | 5.17 a | 525.04 a,b | 0.87 a | 13.08 a | 56.94 c |
| Serbia | |||||||
| Conventional | 12.92 a,b | 4.60 a | 6.62 b | 439.92 a | 0.91 a | 15.10 b | 55.64 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takač, V.; Tóth, V.; Rakszegi, M.; Mikó, P.; Mikić, S.; Mirosavljević, M. The Influence of Farming Systems, Genotype and Their Interaction on Bioactive Compound, Protein and Starch Content of Bread and Spelt Wheat. Foods 2022, 11, 4028. https://doi.org/10.3390/foods11244028
Takač V, Tóth V, Rakszegi M, Mikó P, Mikić S, Mirosavljević M. The Influence of Farming Systems, Genotype and Their Interaction on Bioactive Compound, Protein and Starch Content of Bread and Spelt Wheat. Foods. 2022; 11(24):4028. https://doi.org/10.3390/foods11244028
Chicago/Turabian StyleTakač, Verica, Viola Tóth, Marianna Rakszegi, Péter Mikó, Sanja Mikić, and Milan Mirosavljević. 2022. "The Influence of Farming Systems, Genotype and Their Interaction on Bioactive Compound, Protein and Starch Content of Bread and Spelt Wheat" Foods 11, no. 24: 4028. https://doi.org/10.3390/foods11244028
APA StyleTakač, V., Tóth, V., Rakszegi, M., Mikó, P., Mikić, S., & Mirosavljević, M. (2022). The Influence of Farming Systems, Genotype and Their Interaction on Bioactive Compound, Protein and Starch Content of Bread and Spelt Wheat. Foods, 11(24), 4028. https://doi.org/10.3390/foods11244028